Immunomodulatory Effects of Bio Clean II on Some Inflammatory Cytokines in Rats Exposed to Purified Bacterial Lipopolysaccharide
Bacterial inflammatory diseases have continued to be a major threat to public health. The aim of this study is to determine the effects of Bio-Clean II on serum levels of some selected cytokines in rats exposed to purified bacterial lipopolysaccharide (LPS). A total of 36 male Wistar rats weighing 150g±50g (mean±SD) were purchased and randomly assigned to six (6) groups of 6 rats each. Group 1, 2, 3, 5 and 6 were induced with a single dose of 5mg/Kg of purified LPS® (E.coli 0127:B8, Sigma-Aldrich, St. Louis, USA), administered through intraperitoneal route using 1ml sterile needle and syringe, except for group 4 which served as the zero control (given water and feed only throughout the experiment). Group 1 served as the inflammation control. Group 2 which served as the positive control received 50 mg diclofenac/kg [bid] and 500 mg ciprofloxacin/kg [bid] (positive control) in place of the Bio-Clean. Group 3 which served as the negative control received sterile phosphate buffer saline (PBS). While rats in group 5 and group 6 were treated orally with the herbal remedy “Bio-Clean II” for 7 days and 14 days, respectively. After which, the rats were sacrificed and a cardiac blood specimen was taken from each rat by terminal bleeding from the heart. The serum levels of Interleukin-6 (IL-6), Interleukin-10 (IL-10), Interferon-Gamma (IFN-γ), and Tissue Necrosis Factor-Alpha (TNF-α) were determined using ELISA technique. The outcome of this investigation shows that treatment with the Bio-Clean II resulted in a significant decrease (p=0.000*) in the serum level of IL-6, IFN-γ and TNF-α, but a significant (p=0.000*) increase in the serum level of IL-10 (in a duration-dependent manner) in rats exposed to LPS when compared to the inflammation control group. The outcome of this study suggests that Bio-Clean could find some use in the future for the alleviation of bacterial inflammatory diseases through modulations of host inflammatory responses to pathogens.
Introduction
Bacterial infections have continued to have an enormous impact on the human society despite the discovery of antibiotics [1]. One outcome of bacterial infection is Inflammation [2]. Bacterial lipopolysaccharides (LPS) are powerful provokers of inflammation [3]. Exposure to bacterial lipopolysaccharide (LPS) has been associated with inflammation of body tissues characterized by swelling of the affected area, pain, redness, and even loss of function. Recognition of LPS by the immune system is a complex process involving molecules that bind LPS and pass it on to cells membrane associated receptors on leukocytes, endothelial and other cells, which initiate pro-inflammatory cascade [3, 4]. The natural resolution of inflammation is a highly complex, multifactorial, and tightly controlled process driven by removal of the initial stimulus, decrease in pro- inflammatory mediators (mainly cytokines), elimination of damaged and inflammatory cells, and promotion of repair [2, 4, 5].
Helper and suppressor cells, cytokines, catecholamines, corticosterone, and other soluble compounds all play a role in immune system regulation. Medicinal plants, nutrients, and other variety of natural products/drugs have abilities to impact upon practically every part of the immune system [6, 7, 8, 9]. Over the years, the association between the immune system and use of medicinal plants (or their extracts/by- products) has been well-examined, and in many cases the results suggest that use of this plants/extracts/by product can help improve immunological function [10].
Herbal medicines are an important part of African Traditional Medicine (ATM) because of their “green” image, cultural importance, and availability to people from all walks of life [11, 12, 13]. Among the myriad of constituents in these plants, anti-oxidant-rich Phytoextracts are increasingly receiving attention for potential use as anti-inflammatory agents and immunological boosters in general [14]. In Nigeria, with a large market for herbal medications, the majority of commercially available medicines are hailed as “wonder drugs” capable of healing a wide range of pathologies, including inflammation and immunosuppression [10].
The herbal remedy “Bio-Clean II” has been found to possess some important phytochemicals and trace- elements [15]. It has been previously reported to fight viral infection and boost immunity. Specifically, it has been shown to induce an increase in CD4 Cell Count and body weight, while decreasing viral load in a cohort of HIV positive women [16, 17, 18]. Still it has been previously shown to modulate circulating levels of white blood cells including T-Helper 4 and Cytotoxic T-Lymphocytes, as well as serum levels of C-reactive protein, Corticosterone and anti-phospholipid antibody in rats exposed to purified bacterial LPS [19, 20, 21].
Many of the immunomodulatory properties associated with Bio-Clean II are believed to be directly or indirectly attributable to the presence of some important phytochemicals including alkaloids, saponins, anthraquinones, tannins, and cardiac glycosides [15]. To gain a better understanding of how an herbal remedy like Bio-Clean II might be acting to cause these effects on immune cells in vivo, this study was undertaken to assess the impact of treatment with Bio-Clean II on the LPS-inducible formation of key pro- and anti-inflammatory cytokines. To achieve this, rats were exposed to purified bacterial LPS and then given the Bio-Clean twice daily (by gavage) for 7 or 14 day. Thereafter, serum levels of interleukin (IL)-6, IL- 10, Interferon (IFN)-γ, and Tumor Necrosis Factor (TNF)-α were quantified.
Methodology
Study Design
This is an analytic experimental study using animal models.
Duration of Study
The study lasted for a period of 3 months (April-July, 2021).
Study Area
The study was carried out at the Experimental Animal House, Babcock University, Ilishan-Remo, Ogun State.
Reagents
Bio-Clean II was procured from the manufacturer/maker on demand. Purified lipopolysaccharide (Type 0127:B8 from Escherichia coli, Sigma, St. Louis, MO) was purchased as lyophilized powder. For use in the experiments, the LPS were reconstituted in sterile phosphate-buffered saline (PBS, pH 7.4). Diclofenac and Ciprofloxacin were obtained from the Pharmacy Department of Babcock University Teaching
Hospital, Ilishan-Remo, Ogun State, Nigeria and prepared in sterile distilled water for use in the exposures. All other reagents and kits used in this study were purchased from Elabscience (USA), unless specified otherwise.
Experimental Animals
A total of 36 male Wistar rats weighing 150±50g (mean±SD) were procured from the University of Ibadan’s small animal house and clinically checked upon arrival for any signs of infection or defect. The animals were housed separately in well-ventilated wire-bottom steel cages under hygienic conditions at 25±2oC and a relative humidity of 45–50% at the Experimental Animal House, Department of Animal Science, School of Agriculture and Industrial Technology, Babcock University, Ilishan-Remo (Ogun State, Nigeria). The rats were indiscriminately divided into six groups of six rats each and provided a regular rat diet (10g/100g body weight) twice daily as well as free access to tap water. All rats were allowed to acclimate for 14 days prior to use in any experiment in the Animal House with a regular 12-hour light:dark cycle. All animal experimentations were carried out in compliance with the Institute for Laboratory Animal Research’s Current Animal Care Regulations and Standards [22].
Experimentation
For the study, rats were randomly allocated into six groups (n = 6/group) and each provided a regular rat diet of standard rodent chow as well as ad libitum access to filtered tap water throughout the experimental period. The groups included rats to receive: only LPS (single intraperitoneal injection at 5 mg/ kg; inflammation control); LPS and both 50 mg diclofenac/kg [bid] and 500 mg ciprofloxacin/kg [bid] (positive control); LPS and sterile phosphate-buffered saline (negative control), no treatments [water and feed only; zero control); LPS and Bio- clean II for 7 d; and, LPS and Bio-clean II for 14 d. All animals, including the controls were gavaged for 7 or 14 days, as the case may be. Therapy with Bio-clean II was initiated by day 2 post-LPS challenge. Before each treatment, the volume of Bio- clean II suspension to be delivered to each rat via intragastric tube was estimated based on host weight. The rats in those groups were given an appropriate volume of suspension (never to exceed 5 ml/kg) twice daily, every 12 hrs. (between 6.00-6.30 AM and PM) for the total number of days specified in the experimental protocol. The suspension was shaken gently before administration and administered. Each time, the rats were monitored for any changes in appearance, appetence, etc. At Days 8 and 15 (24 hr after the respective final exposures for the 7 and 14-d regimens), the rats were euthanized by cervical dislocation as described by Ochei & Kolhatkar [23]. At necropsy, cardiac blood specimens were collected and serum prepared for later evaluation of select inflammatory cytokines.
Laboratory Analyses
Serum interleukin (IL)-6, IL-10, interferon (IFN)-γ, and tissue necrosis factor (TNF)-α levels were determined using commercial rat ELISA kits (Elabscience, USA) as described by Chiswick, et al. [24] according to the manufacturer’s instruction. In brief, the serum samples were diluted (10- fold steps) using serum matrix diluent as provided in the kit by manufacturer. One hundred microlitres (100µL) of standard or sample was added to each well (in triplicates) and then incubated for 90 minutes at 37ᴼC. The sample was then decanted and 100 microlitres of Biotinylated Detection Ab/Ag was added and incubated for 1 hour at 37ᴼC. After incubation, it was then aspirated and washed 3 times. 100 microlitres of Avid-HRP Conjugate was added and then incubated for 30 minutes at 37ᴼC. After incubation it was then aspirated and washed 5 times with wash buffer. Ninety microlitres (90µL) of Sub state Reagent was added and the incubated for 15 minutes at 37ᴼC. Then fifty microliter (50µL) of Stop Solution was then added to stop the reaction. The optical densities of the plates were immediately read at a wave length of 450nm using a KC-100 micro plate ELISA reader (BMG Labtech, Germany). All data were recorded as absorbance and conversion made based on extrapolated standards.
Data Analyses
All data are reported as means ± SD. All data were analyzed using a one-way analysis of variance (ANOVA) and a Turkey-Kramer Multiple Comparison Test. A p-value ≤ 0.05 was taken as significant by Shott [25]. Analyses were performed using SPSS-20.0 software (Statistical Package for Social Scientists, Chicago, IL).
Results
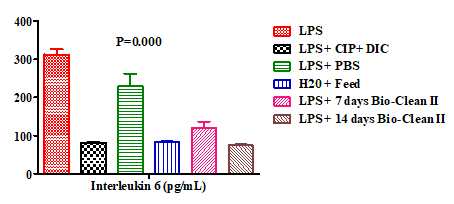
This present study assessed the effect of “Bio-clean II” on some cytokines in experimental inflammation using an animal model. The effect of Bio-Clean II on the serum levels of interleukin 6 (IL-6) in rats exposed to purified bacterial lipopolysaccharide were presented using a bar chart (Figure 1). The serum level of IL-6 of the 7 days (Group 5: 119.35±17.36pg/mL) and 14 days (Group 6: 74.47±2.27pg/ mL) Bio-Clean II treated rats was found to be significantly lower (p=0.000* and p=0.001*, respectively) when compared to the inflammation control (Group 1: 312.62±13.93pg/ mL). It also decreased significantly (p=0.030* and p=0.012*, respectively) when compared to the Negative control (Group 3: 229.89±32.74pg/mL). Although when compared to the positive control (Group 2: 81.42±1.11pg/mL), it was found to be non-significantly lower (p= 0.773 and p=0.412, respectively), and when compared to the Zero control (Group 4: 83.25±1.55pg/mL) it was non-significantly higher (p= 0.814 and p= 0.204, respectively). Meanwhile, the serum level of IL-6 of the 14 days Bio-Clean II treated group (Group 6: 74.47±2.27pg/mL), when compared with that of the 7 days Bio-Clean II treated group (Group 5: 119.35±17.36pg/ mL) was non-significantly lower (p= 0.607). Meanwhile, the serum level of IL-6 of the positive control (Group 2: 81.42±1.11pg/mL) when compared to that of the negative control group (Group 3: 229.89±32.74pg/mL) was found to be significantly lower (p=0.014).
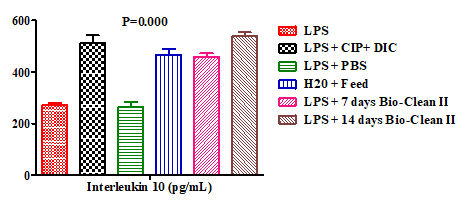
Furthermore, the effect of Bio-Clean II on the serum levels of interleukin 10 (IL-10) in rats exposed to purified bacterial lipopolysaccharide were presented using a bar chart (Figure 2). The serum level of IL-10 of the 7 days (458.81±12.92pg/mL) and 14 days Bio-clean II treated groups (538.96±15.79pg/mL) was found to be significantly higher (p=0.000* for each) when compared to the inflammation control group (272.76±8.16pg/mL). It also increased significantly ((p=0.002* and p=0.000*, respectively) when compared to the negative control group (262.68±20.42pg/mL). Although when compared to the positive control (512.36±29.25pg/mL), it was found to be non-significantly higher (p=0.912 and p=1.000, respectively) and when compared to the zero control (466.82±22.72pg/ mL), it was non-significantly higher (p=1.000 and p= 0.409, respectively). Moreover, the serum level of IL-10 of the 14 days Bio-clean II treated group (group 6: 538.96±15.79pg/ mL) when compared with that of the 7 days Bio-clean II treated group (group 5: 458.81±12.92pg/mL) was non- significantly higher (p= 0.069). Meanwhile, the serum level of IL-10 of the positive control group (512.36±29.25pg/mL) was found to be significantly higher (p=0.003*) than that of the negative control group (262.68±20.42pg/mL).
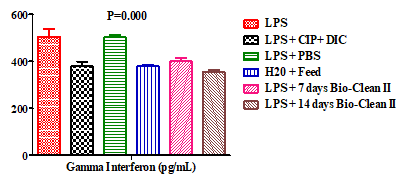
In addition, the effect of Bio-Clean II on the serum level of Interferon-gamma (INF-ƴ) in rats exposed to purified bacterial lipopolysaccharide were presented using a bar chart (Figure 3). The serum level of INF-ƴ of 7 days (400.65±14.62pg/mL) and 14 days (355.15±9.76pg/mL) Bio-Clean II treated group was found to be significantly p=0.030* and p=0.003* respectively) lower when compared with the inflammation control (506.12±32.04pg/mL). When compared with the negative control (502.47±11.65pg/mL), the serum level of 7 days (400.65±14.62pg/mL) and 14 days (355.15±9.76pg/mL) in Bio-Clean II treated rats was found to be non-significantly lower (p=0.11) and significantly lower (p=0.000*), respectively. However, when compared with the Positive control (380.30±19.26pg/mL), the serum level of 7 days Bio-Clean II treated rats (400.65±14.62pg/ mL) was found to be non-significantly higher (p=1.000), while that of 14 days (355.15±9.76pg/mL) was found to be non-significantly lower (p=0.994). The INF-ƴ serum level of the 7 days Bio-Clean II treated group (400.65±14.62pg/mL) was found to be non-significantly higher (p=0.983) than that of the zero control (379.41±6.60pg/mL), while that of the 14 days Bio-Clean II treated group (355.15±9.76pg/ mL) was found to be non-significantly lower (p=0.706). The INF-ƴ serum level of the 14 days Bio-Clean II treated group (355.15±9.76pg/m) was found to be non-significantly lower (P=0.424) when compared to that of 7 days (400.65±14.62pg/mL). Meanwhile, the INF-ƴ serum level of the positive control (380.30±19.26pg/mL) was found to be significantly lower (p=0.017*) when compared with that of the negative control group (Group 3: 502.47±11.65pg/ mL).
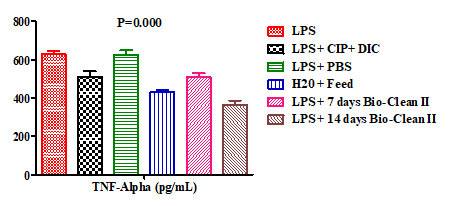
Still, the effect of Bio-Clean II on the serum levels of Tissue Necrosis Factor-Alpha (TNF-α) in rats exposed to purified bacterial lipopolysaccharide were presented using a bar chart (Figure 4). The TNF-α serum level of the 7 days (509.20±19.30pg/mL) and 14 days (365.80±20.73pg/mL) Bio-Clean II treated rats was found to be significantly lower (p=0.026* and p=0.000*, respectively), when compared to the inflammation control (627.60±16.55pg/mL). Meanwhile, the serum level of the 7 days Bio-Clean II treated rats was noticed to be non-significantly lower (p=0.082) when compared to the Negative control (626.00±23.94pg/ mL), while that of the 14days was found to be significantly lower (p=0.001*). Moreover, when compared to the positive control (511.00±29.08pg/mL), it was noticed that TNF-α serum levels of the 7 days (509.20±19.30pg/mL) and 14 days (365.80±20.73pg/mL) Bio-Clean II treated rats were both found to be non-significantly lower (p=1.000 and p=0.065). We also found out that the TNF-α serum level of the 14 days Bio-Clean II treated group (365.80±20.73pg/ mL) was found to be significantly lower (p=0.015*) when compared to that of the 7 days (509.20±19.30pg/mL). The serum level of Tissue Necrosis Factor-Alpha (TNF-α) of the positive control (511.00±29.08pg/mL) when compared with that of the negative control (626.00±23.94pg/mL) was found to be non-significantly lower (p=0.220).
Discussion
The human immune system plays a critical function in protecting the body against foreign chemicals and other disease-causing agents [10]. Due to the development of antibiotic resistance by bacteria [26], researchers are now thinking of using immunomodulation as a means to help control infections [27]. This approach can include the pharmacological manipulation of immune system activity to cause an increase or decrease in magnitude of the immune response [28].
This present study assessed the effects of Bio-Clean II on the serum level of selected cytokines in experimental LPS- induced inflammation animal model. Lipopolysaccharides (LPS) are stronger provokers of inflammation. They are characteristic components of the cell wall of Gram negative bacteria (absent in Gram positive bacteria). They are localized in the outer layer of the membrane and are in non-capsulated strains, exposed on the cell surface. They contribute to the integrity of the outer membrane, and protect the cell against the action of bile salts and lipophilic antibiotics. They are recognized by immune cells as a pathogen-associated molecule through Toll-like receptor [29]. According to Gardner, et al. [30], the activation of mononuclear cells by endotoxin forms of LPS leads to the secretion of a wide spectrum of endogenous mediators, among which are pro- inflammatory cytokines such as IL-6.
In this study, the serum level of interleukin-6 was significantly reduced in a duration dependent manner in the 7- and 14-day (p<0.001) Bio-Clean II treated rats when compared to the inflammation (LPS only) control. This suggests that Bio-clean II could impart antioxidant and anti-inflammatory effect in vivo. This particular outcome is consistent with the work of Ogunana, et al. [31] who assessed the ameliorative effect of ruzu herbal bitters on the biochemical and antioxidant abnormalities induced by high fat diet in wistar rats. It is also consistent with the work of Lee, et al. [32] who investigated the anti-inflammatory effects of Pulsatila Korean (an herbal drug). In that study, plasma levels of IL-6 were measured before and after injection of the herbal drug; it was seen that while there was a 3-fold increase in IL-6 within 5 hr of LPS injection, drug treatment attenuated the magnitude of this increase in the IL-6 levels in a dose-dependent manner. The present findings also agree with the work of Mirzaee, et al. [33] who demonstrated the potential of “IMOD” herbal medicine to modulate the immune functions of human dendritic cells (DC). Specifically, IMOD caused a decrease in production of IL-6 by LPS-activated DC at both the mRNA and protein levels. The outcome of the current study was also in agreement with the work of Park, et al. [34] who investigated the effect of “SC-E1” (novel herbal formulation in Korea) on LPS-stimulated macrophages. Those investigators found that the herbal formulation could inhibit inducible production of pro-inflammatory cytokines (including IL-6) without affecting cell viability. Mechanistic studies showed that LPS-induced nuclear factor (NF)- kB activation and increased activity within the mitogen- activated protein kinase (MAPK) pathway were both inhibited by SC-E1. Lastly, Ogunlana, et al. [35] were able to show that a Ruzu bitter herbal treatment could modulate the levels of cytokines, resulting in a lowering of inflammation, in part due to a likely inactivation of the NF-κB pathway and positive regulation of proliferator-activated receptor (PPAR)-α, a ligand-activated transcriptional factor that belongs to the family of nuclear receptors and which itself has a role in decreasing inflammation, mainly through direct interaction with NF-κB [36].
The authors here note that when the effects in the positive control hosts was compared with those in the inflammatory (LPS only) control; the serum level of IL-6 was considerably lower in the positive control. This is likely because ciprofloxacin and diclofenac differentially modulate IL-6 expression through impact on a response to infection (controlled by a cascade of cytokines and other mediators), resulting in changes in local cytokine concentrations [37]. Whether the effects here are being mediated through endothelium cells as noted in that study remains to be verified. It is possible other cell types may also be modulated by the two drugs.
Interleukin (IL)-10 is an anti-inflammatory cytokine that plays a key role in suppressing a host immune response to infection, thereby minimizing harm from inflammatory and auto-immune-related pathologies, and helping to maintain normal tissue homeostasis. This is borne out by the fact that IL-10 dysregulation can give rise to increased immunopathologies in response to infections [38, 39]. While an IL-10 deficiency/abnormal expression can boost a host inflammatory response to bacterial challenge, this scenario can also lead to inflammatory bowel disease and a variety of autoimmune diseases [40]. As a result, impaired IL-10 expression or signaling can improve pathogen clearance during an acute infection, but concurrently lead to exacerbated inflammatory responses, resulting in immunopathologies and tissue damage [41]. Some pathogens can use the immunosuppressive properties of IL-10 to suppress a host immune response, resulting in long-term infection. Overall, IL-10 plays a largely non-redundant role in mediating host anti-inflammatory responses. Thus, identifying cellular sources of IL-10, as well as molecular mechanisms that regulate IL-10 expression are critical for the development of therapeutic strategies to address pathology-related IL-10 production impairment [42].
In this study, animals treated with the Bio-Clean II had a significant increase in serum levels of IL-10 post-LPS induction. This outcome is in agreement with results of a study done by Lee, et al. [43] that investigated anti-inflammatory effects of a Korean herbal medicine Pulsatilla koreana in rats treated with LPS. Pulsatilla koreana (at a dose of 200 mg/kg) boosted plasma and hepatic levels of anti-inflammatory IL- 10 in the LPS-exposed rats (200% above control rat values) - but caused lower levels of pro-inflammatory IL-1, IL-6, and TNFα at both sites. This change in IL-10 was also evident here, when the serum IL-10 value of the 14-d Bio-Clean- treated rats was compared to the inflammation control (LPS only) rats.
Interferon (IFN)-γ, a major component in cellular immunity, can orchestrate a variety of protective actions to boost immune responses in infections and malignancies. Enhancing antigen processing and presentation, increasing leukocyte trafficking, generating an antiviral state, strengthening antimicrobial capabilities, and altering cellular proliferation and death are all examples of immunomodulatory actions of IFNγ [41]. T-helper (TH) cells are a type of T-lymphocyte that aids in the enhancement or potentiation of the activity of other immune cells (mostly TH1 and TH2 cells) by releasing T-cell cytokines, TH1 lymphocytes, which produce IFNγ and TNFα, contribute to cellular immunity.
In the current study, although non-significant, serum IFNγ levels increased in the rats given only LPS and those that received LPS + phosphate buffer. This is in agreement with a study by Lee and Sullivan [43] who suggested that IFNγ has the ability to up-regulate the LPS-inducible TNFα mRNA and protein production. LPS is a potent inducer of monocyte and macrophage TNFα expression. However, IFNγ potentiated the effects of LPS on TNFα [44]. IFNγ potentiates the generation of TNFα by macrophages in a concentration and time-dependent way, and this action is blocked by anti- IFNγ treatment, as demonstrated in the group given anti- inflammatory drugs and anti-LPS [45].
The inflammation control group showed no significant difference in the mean values of TNFα when compared to group two (positive control) and group three (negative control), but showed a statistically significant difference in mean values when compared with the remaining negative control administered with water and experimental groups with different duration of Bio-Clean II administration. This is an evidence of elevated TNFα in response to solitary LPS exposure, with a time-dependent decrease in concentration. Antibiotics, anti-oxidants, and anti-inflammatory drugs reduce Interferon-gamma, which is considered the action of the flavonoid and tannin component of Bio-Clean II in this case study. The drop in TNFα levels, either as a result of time-dependent Interferon-gamma or as a result of anti-inflammation and anti-oxidative stress properties phytoconstituents of Bio-Clean II, can be argued to support the aforesaid theory.
The outcome of this study was consistent with the work of Ahmad, et al. [46] that examined immunomodulating effects of the extract of Tinospora crispa (TCE) on Wistar Kyoto rat innate immune responses. In those studies, it was observed that rats treated with TCE (for 21 d) had significantly increased blood IFNγ (400.2 [± 16.2] pg/ml) and TNFα (550.30 [±11.4] pg/ml) levels in comparison with control (120.0 [± 8.3] and (180.2 [± 12.3] pg/ml, respectively) values.
Furthermore, when compared with a similar study done by Peng, et al. [47] on the effects of Jianpi huoxue decoction on inflammatory cytokine secretion pathway in rat liver with lipopolysaccharide challenge, (for 21 days), serum levels of TNFα when compared to controls were significantly reduced (12 225.40±9007.3pg/mL vs 6031.70±2296.56pg/mL). Additionally, in comparison with a previous study carried out by Zhang, et al. [48] on bioactive components from Qingwen Baidu Decoction against LPS-induced acute lung injury in rats for 7 days, the levels of tumour necrosis factor-a were evidently reduced when compared to the LPS group.
Conclusion
The results of this study show that the herbal remedy Bio-Clean II imparts anti-inflammatory properties in rats by regulating the serum level of IL-6, IL-10, TNFα, and IFNγ From this, with further study, it is possible that one day the use of Bio-Clean II might be considered a suitable alternative therapy for the treatment of bacterial inflammatory diseases. This study showed that these effects of the herbal remedy were exposure duration-dependent, albeit that the caveat remains that caution should be taken to try to avoid potential cellular toxicities that may be associated with prolonged exposures. Data generated in this study could potentially be extrapolated to humans; however, further studies will be needed to understand the molecular mechanisms behind the effects observed herein.
Ethical Approval
Ethical approval for the study was obtained from the Babcock University Health Research Ethics Committee (BUHREC) with ethical approval registration number: BUHREC 388/21.
Declaration of Interest
The authors report no conflict of interest. The authors alone are responsible for the content of this manuscript.
Data Availability
The data that support the findings of this study are available from the corresponding author, [Enitan S. S.], upon reasonable request.
References
-
Janeway C, Travers P, Walport M, Shlomchik M (1996) Immunobiology: the immune system in health and disease. London: Current Biology 7: 26.
-
Levi M, Keller TT, van Gorp E, ten Cate H (2003) Infection and inflammation and the coagulation system. Cardiovasc Res 60(1): 26-39.
-
Mayer H, Tharanathan RN, Weckesser J (1985) 6 Analysis of Lipopolysaccharides of Gram-negative bacteria. Meth Microbiol 18: 157-207.
-
Dube MP, Sattler FR (2010) Inflammation and complication of HIV disease. J Infect Dis 201(12): 1783- 1785.
-
Kussmann M (2010) Nutrition and Immunity. In: Fay LB (Eds.), Mass Spectrometry and Nutrition Research. Royal Society of Chemistry, pp: 268-309.
-
Kandil OM, Abdellah TH, Elkadi A (1987) Garlic and the immune system in humans: its effects on natural killer cells. Federal Procedures 46: 441.
-
Arunachalam K, Gill HS, Chandra RK (2000) Enhancement of natural immune function by dietary consumption of Bifidobacterium lactis (HN019). Eur J Clin Nutr 54(3): 263-267.
-
Ali HI, Al-Shawi SG, Habib HN (2019) The Effect of Nutrition on Immune System. Food Science and Quality Management 90: 31-35.
-
Wu D, Lewis ED, Pae M, Meydani SN (2019) Nutritional Modulation of Immune Function: Analysis of Evidence, Mechanisms, and Clinical Relevance. Front Immunol 9: 3160.
-
Mirabeau TY, Samson ES (2012) Effects of Allium sativum and Allium cepa on some Immunological Cells in Rats. Afr J Tradit Complement Altern Med 9(3): 374-379.
-
Iwu MM (2002) Ethnobotanical approach to pharmaceutical drug discovery: strengths and limitations. In: Ewe MM (Eds), Ethnomedicine and Drug Discovery. Elsevier, Amsterdam, pp: 309-320.
-
World Health Organization (2001) Programme on Traditional Medicine: Legal Status of Traditional Medicine and Complementary/ Alternative Medicine, Worldwide Review.
-
World Health Organization (2008) Herbal Medicine. WHO, Geneva.
-
Ngcobo M, Gqaleni N (2016) Evaluation of the immunomodulatory effects of a South African commercial traditional immune booster in human peripheral blood mononuclear cells. BMC Complement Altern Med 16(1): 300.
-
Enitan SS, Ibeh IN, Isitua CC, Dada MO, Mensah-Agyei GO et al. (2022). Bacteriological Quality, Phytochemical Screening, Trace Element and Proximate Analysis of Bio-Clean II Herbal Remedy. Journal of Medicinal Herbs, Article ID: MEDHERB-2208-1473 (In Press).
-
Ibeh IN, Akanu N, Mkpa AM, Isitua CC, Ogefere HO, et al. (2013) Evaluation of the Anti-Human Immunodeficiency Virus (HIV) Properties of DXL (Decoction X-Liquid- Bioclean II). J Clin Toxicol 12(4): 1-7.
-
Ibeh IN, Okungbowa MA, Ibeh NI, Adejumo BI (2016) Comparative studies on the effects of Zidovudine + Nevirapine + Lamivudine and Bio-clean II on female HIV/ AIDS cases in Nigeria. Journal of Advances in Medical and Pharmaceutical Sciences 9(3): 1-7.
-
Ibeh IN, Okungbowa MA, Ibeh NI, Isitua CC (2016) Studies on HIV Resistance to Multiple Drug Therapy (Lamivudine, Zidovudine and Nevirapine) in Benin Metropolis, Nigeria. Int J Sci Res Environ Sci Toxicol 1(1): 1-3.
-
Enitan SS, Ibeh IN, Nwaenyi KE, Adelakun A, Akele RY, et al. (2022) Immunomodulatory Effects of Bio-Clean II on T-Helper 4 and Cytotoxic T-Lymphocytes in Rats Exposed to Purified Bacterial Lipopolysaccharide. Ann Immunol Immunother 4(1): 000165.
-
Enitan SS, Ibeh IN, Olayanju AO, Ileoma EO, Durosinmi A, et al. (2022) Immunomodulatory Effects of Bio-Clean II on Total and Differential Leucocyte Counts in Rats Exposed to Purified Bacterial Lipopolysaccharide. Open J Pathol Toxicol Res 1(2): 1-8.
-
Enitan SS, Ibeh IN, Adelakun AA, Akele RY, Olayanju AO, et al. (2022). Immunomodulatory Effects of Bio-Clean II Herbal Remedy on C-reactive protein, Corticosterone and Antiphospholipid antibodies in Rats exposed to Purified Bacterial Lipopolysaccharide. Journal of Clinical & Experimental Immunology 7(3): 476-484.
-
ILAR (1996) Guide for the care and use of laboratory animals. The National Academies Press, Washington 11(8): 131.
-
Ochei J, Kolhatkar A (2006) Euthanasia of Animals by Cervical Dislocation. In: Ochei J (Eds.), Theory and Practice of Medical Laboratory Science. Tata McGraw- Hill publishing Company Limited, New Delhi, pp: 1213- 1230.
-
Chiswick EL, Duffy E, Japp B, Remick D (2012) Detection and Quantification of Cytokines and Other Biomarkers. Methods Mol Biol 844: 15-30.
-
Shott S (1990) Statistics for health professionals. Saunders, W.B., Co. Philadelphia, pp: 313-336.
-
Peterson E, Kaur P (2018) Antibiotic Resistance Mechanisms in Bacteria: Relationships between Resistance Determinants of Antibiotic Producers, Environmental Bacteria, and Clinical Pathogens. Front Microbiol 9: 2928.
-
Kumar V, Shriram V, Mulla J (2013) Antibiotic resistance reversal of multiple drug resistant bacteria using Piper longum fruit extract. J App Pharm Sci 3(3): 112-116.
-
Kyo E, Uda N, Kasuga S, Itakura Y (2001) Immunomodulatory effects of aged garlic extract. J Nutr 131(3S): 1075S-1079S.
-
Wang X, Quinn PJ (2010). Endotoxins: Lipopoly saccharides of Gram-Negative Bacteria. In: Endotoxins: Structure, Function and Recognition. Subcellular Biochemistry, 53: 3-25.
-
Gardner ST, Assis VR, Smith KM, Appel AG, Mendonça MT, et al. (2020) Innate immunity of Florida cane toads: how dispersal has affected physiological responses to LPS. J Comp Physiol B 190(3): 317-327.
-
Ogunlana OO, Ogunlana OE, Ugochukwu SK, Adeyemi AO (2018) Assessment of the ameliorative effect of ruzu herbal bitters on the biochemical and antioxidant abnormalities induced by high fat diet in wistar rats. International Journal of Pharmacology 14(3): 329- 341.
-
Lee SH, Lee E, Ko YT (2012) Anti-inflammatory effects of a methanol extract from Pulsatilla koreana in lipopolysaccharide-exposed rats. BMB Rep 45(6): 371- 376.
-
Mirzaee S, Drewniak A, Sarrami-Forooshani R, Kaptein TM, Gharibdoost F, et al. (2015) Herbal medicine IMOD suppresses LPS-induced production of proinflammatory cytokines in human dendritic cells. Front Pharmacol 6: 64.
-
Park JY, Kwon YW, Lee SC, Park SD, Lee JH, et al. (2017) Herbal formula SC-E1 suppresses lipopolysaccharide- stimulated inflammatory responses through activation of Nrf2/HO-1 signaling pathway in RAW 264.7 macrophages. BMC Complement Altern Med 17(1): 374.
-
Ogunlana OO, Ogunlana OE, Adekunbi TS, Adetuyi BO, Adegboye BE, et al. (2020) Anti-inflammatory Mechanism of Ruzu Bitters on Diet-Induced Nonalcoholic F a t t y Liver Disease in Male Wistar Rats. Evidence-Based Complementary and Alternative Medicine 2020: 8.
-
Won TW (2013) Fenofibrate, a PPAR-α agonist, blocks lipopolysaccharide-induced inflammatory pathways in mouse liver. Korean J Hepatobiliary Pancreat Surg 17(3): 89-108.
-
Galley HF, Nelson SJ, Dubbels AM, Webster NR (1997) Effect of Ciprofloxacin on the accumulation of interleukin-6, interleukin-8, and nitrite from a human endothelial cell model of sepsis. Crit Care Med 25(8): 1392-1395.
-
Iyer SS, Cheng G (2012) Role of Interleukin 10 Transcriptional Regulation in Inflammation and Autoimmune Disease. Crit Rev Immunol 32(1): 23-63.
-
Sabat R, Grütz G, Warszawska K, Kirsch S, Witte E, et al. (2010) Biology of interleukin-10. Cytokine Growth Factor Rev 21(5): 331-344.
-
O’Garra A, Barrat FJ, Castro AG, Vicari A, Hawrylowicz C, et al. (2008) Strategies for use of IL-10 or its antagonists in human disease. Immunol Rev 223: 114-131.
-
Siewe L, Bollati-Fogolin M, Wickenhauser C, Krieg T, Müller W, et al. (2006) Interleukin‐10 derived from macrophages and/or neutrophils regulates the inflammatory response to LPS but not the response to CpG DNA. Eur J Immunol 36(12): 3248-3255.
-
Roncarolo MG, Gregori S, Battaglia M, Bacchetta R, Fleischhauer K, et al. (2006) Erratum: Interleukin- 10-secreting type 1 regulatory T cells in rodents and humans. Immunol Rev 212: 28-50.
-
Lee JY, Sullivan KE (2001) Interferon-gamma and lipopolysaccharide interact at the level of transcription to induce tumor necrosis factor-alpha expression. Infect Immun 69(5): 2847-2852.
-
Kak G, Raza M, Tiwari BK (2018) Interferon-gamma (INF-ƴ): exploring its implications in infectious diseases. Biomol Concepts 9(1): 64-79.
-
Déry RE, Bissonnette EY (1999) IFN-gamma potentiates the release of TNF-alpha and MIP-1alpha by alveolar macrophages during allergic reactions. Am J Respir Cell Mol Biol 20(3): 407-412.
-
Ahmad W, Jantan I, Kumolosasi E, Bukhari SN (2015) Immunostimulatory effects of the standardized extract of Tinospora crispa on innate immune responses in Wistar Kyoto rats. Drug Des Devel Ther 9: 2961-2973.
-
Peng JH, Hu YY, Cheng Y, Han C, Xu LL, et al. (2008) Effect of Jianpi huoxue decoction on inflammatory cytokine secretion pathway in rat liver with lipopolysaccharide challenge. World J Gastroenterol 14(12): 1851-1857.
-
Zhang Q, Lei HM, Wang PL, Ma ZQ, Zhang Y, et al. (2017) Bioactive Components from Qingwen Baidu Decoction against LPS-Induced Acute Lung Injury in Rats. Molecules 22(5): 692.
- Epidemiological and Clinical Aspects of Intestinal Parasitoses Among Students in the City of Bocaranga, Central African Republic
- Artificial Intelligence Empowers Global Infectious Disease Prevention and Control: Opportunities and Challenges
- Factors that Affect the Incidence of Babesia and Blood Donor Testing in Select States: A Regression Analysis
- Neuro-TB: The Battle between Tuberculosis and the Nervous System
- The Biological and Health Implications of Cat Fleas (Ctenocephalides felis): Assessing Zoonotic Risks and Hygiene Strategies
- Biostatistical Analysis of Medicinal Plants for Treating Schizophrenia