Synaptic Diversity and Quantification of Intra-Hippocampal Contextual Memory
The hippocampal neurons seem to process both space and time information within a specific episode. However, the integrated system how to sustain a piece of specific memory or what associates the memory fragment each other is largely unknown. Bliss & Lømo (1973) reported long-term potentiation (LTP) in rabbit hippocampus, which has been considered as a cellular model of hippocampal memory. Hayashi, et al. (1999) clearly showed that the synaptic delivery of AMPA receptor is a molecular mechanism of LTP. Although the molecular mechanism of in vivo hippocampal learning had been unknown, we showed that learning-dependent synaptic delivery of AMPA receptors into the CA3-CA1 synapses is required for contextual learning. More importantly, contextual learning not only induces synaptic delivery of AMPA receptors but also strengthens GABAA receptor-mediated inhibitory synapses onto the CA1 neurons. Although each CA1 neurons showed different strength of the excitatory/inhibitory synapses in untrained animals, contextual learning clearly diversified the strength of excitatory/inhibitory synapses. Entropy analysis of the diversity may allow us to quantify the amount of information in trained animals.
Introduction
The hippocampus plays a central role to form new episodic memory in various species including humans [1]. The hippocampal neurons seem to process variety of information, such as spatial location [2], temporal information [3] and emotional state [4] within specific episodes [5]. However, the basic rules such as how to sustain a piece of specific memory and what associates the memory fragment each other are completely unknown.
Our Findings and Hypothesis
Contextual Training
Stressful experience sometimes forms strong episodic memory in the hippocampus [6], which is useful to avoid similar risk in future. As a learning paradigm, we used an inhibitory avoidance (IA) task. In the task, we placed a rat in an illuminated box. When the rat entered into the dark side of box, we applied brief electrical foot shock in the dark side. Even one-time foot shock experience, the rat quickly learns the experienced episode. The rats were returned to the home cage, then 30 min after, we placed the rat in the illuminated box again. We measured the latency in the illuminated box as a learning performance (Figure 1).
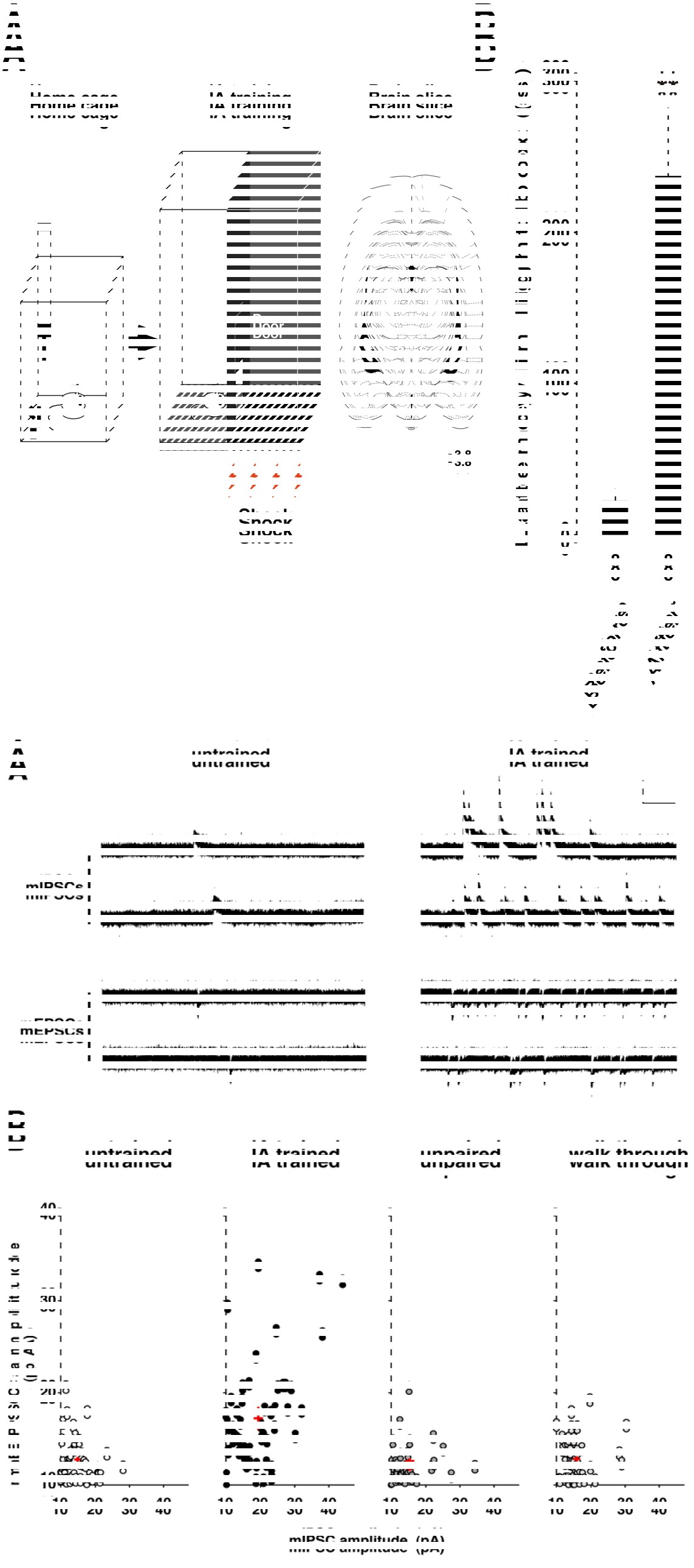
Figure 1 A: Schema of inhibitory avoidance (IA) task. Rats were experienced an episode with an electrical foot shock.
B: After the experience, the rats showed longer latency in the illuminated side of the box [7, 8]. After the training, we made acute brain slices to analyze synaptic function using slice patch-clamp technique [7, 8]. Although some aspect of the training- induced synaptic plasticity is similar to the tetanus stimuli-induced LTP [9, 10], we found many novel aspects of plasticity in trained animals.
Contextual training creates synaptic diversity
For example, to analyze the plasticity, we recorded the postsynaptic response induced by a single vesicle of GABA (mIPSC) or glutamate (mEPSC) in the presence of a Na+ channel blocker (Figure 2A). The means of mEPSC amplitude and mIPSC amplitude were calculated in each neuron, and the data was plotted in Figure 2B. Although the amplitudes were low and showed a narrow distribution range in untrained unpaired, or walk-through rats, IA training not only strengthened AMPA receptor- mediated excitatory inputs, but also increased GABAA receptor-mediated inhibitory synaptic inputs in CA1 neurons (Figure 2B). These results suggest that the training promoted the diversity of excitatory and inhibitory inputs in CA1 neurons.
Learning Requires the Plasticity at Excitatory Synapses
Question arises as to whether the learning requires the synaptic plasticity. We found bilateral CA1 expression of GluA1-c-tail or MPR-DD, an AMPA receptor-delivery blocker, successfully impaired the learning [8], suggesting the contextual learning requires the synaptic delivery of GluA1 containing AMPA receptors. Moreover, recent chromophore-assisted light inactivation technique (CALI) further demonstrated that optical inactivation of synaptic AMPA receptors can erase acquired memory [11], These results further showed that newly delivered GluA1- containing AMPA receptors contribute to forming contextual memory.
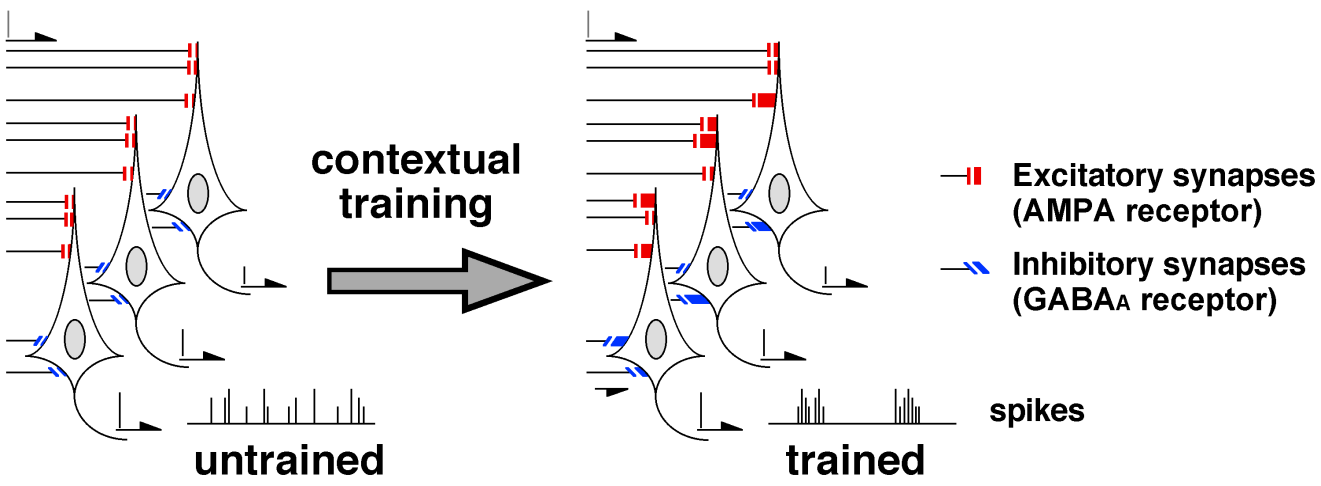
As to the inhibitory synaptic plasticity, nicotinic α7 receptor antagonist is known to block the strengthening of GABAA receptor-mediated synapses [7, 12]. Since pharmacological blockade of the plasticity in bilateral CA1 impaired the learning, contextual learning seems to require the inhibitory synaptic plasticity in the CA1. Moreover, optogenetic enhancement and suppression technique further revealed the role of the GABAergic inhibitory synapses in learning [13]. Taken together, I hypothesized that the contextual learning requires the training-induced synaptic plasticity at both excitatory and inhibitory synapses in CA1 pyramidal neurons (Figure 3).
Learning Requires the Plasticity at Inhibitory Synapses
Entropy Analysis of the Synaptic Diversity
The learning-induced synaptic diversity can be analyzed mathematically. By calculating the appearance probability of each point of Figure 2B, data from each neuron can be converted to self-entropy (bit) using the information theory of Ralph Hartley and Claude Shannon CE [17, 18]. A point with high appearance probability (around the mean level) indicates low self-entropy, while a point with very rare probability (a deviated point) indicates high self-entropy. Compared with untrained rats, the self-entropy per neuron was clearly increased in IA-trained rats but not in unpaired or walk-through rats [7]. This analysis suggests a learning-induced increase in the amount of information in CA1 neurons. Although each neuron showed different self-entropy level, we calculated self-entropy level per single CA1 neuron after the training [19]. By multiplying 4.0 x 105, the total number of CA1 neurons in rats [20], we could estimate a possible amount of intra-CA1 self-entropy after the training. Synaptic inputs regulate neuron firing according to the all-or-none principle [21]. As neurons are considered an all-or-none device [22, 23] , one neuron can handle 1-bit of memory per clock cycle (log2 2 = 1 bit) according to a principal formula of information theory [17, 18]. Based on the principle, computational theory proposed a role of the hippocampus as a kind of memory device [24]. Our previous finding of a logarithmic correlation between the number of cells blocking plasticity and learning performance [8] may provide evidence that each CA1
neuron transmits binary data, forming a contextual memory, such as what, where, or when.
Entropy Analysis of Genome DNA
Since many diverse features contain entropy, the synapse is not a solo factor to be analyzed. For example, 3.1 x 109 base pairs of the human genome quaternary system are identical to 6.2 x 109 bits in a binary system [25, 26, 27]. However, 6.2 x 109 bits contain only 755 megabytes (1 byte = 8 bits), the amount is not more than one hour of short movies. Although postnatal maternal care affect the offspring epigenetic programming [28], postnatal experience cannot affect the genomic quaternary information. Because our brains process much more information per day, an understanding of genome information cannot be sufficient to reveal the learning systems.
Entropy Analysis of Firing In Vivo
The diversified excitatory/inhibitory synapses after training may create a variety of ripple-like on/off firing in hippocampal CA1 (Figure 3). In fact, hippocampal ripple- like events seem to exhibit diverse features [15, 29] and phase-locked (≈ 180°) with theta waves [16]. Interestingly, all the published ripple-like events from many research groups were not identically same. Since selective suppression of hippocampal ripples impairs spatial memory in dorsal CA1 [30], the ripple-like firing may code a piece of experienced information in the CA1.
Conclusion
Our results, together with the previous studies, we hypothesized that the excitatory/inhibitory synaptic plasticity creates a variety of ripple-like events to process the experienced context by using the theta phase-locked processing system in the CA1. We are going to analyze the diversity of the ripple-like events in trained animals, that would be necessary to decode intra-CA1 information.
References
-
Scoville WB, Milner B (1957) Loss of recent memory after bilateral hippocampal lesions. J. Neurol Neurosurg. Psychiatry 20(1): 11-21.
-
Wills TJ, Cacucci F, Burgess N, O'Keefe J (2010) Development of the hippocampal cognitive map in preweanling rats. Science 328:1573-1576.
-
Mitsushima D, Takase K, Funabashi T, Kimura F (2009) Gonadal steroids maintain 24-h acetylcholine release in the hippocampus: organizational and activational effects in behaving rats. J Neurosci, 29: 3808-3815.
-
Chen G, Wang LP, Tsien JZ (2009) Neural population- level memory traces in the mouse hippocampus. PLoS ONE 4(12): e8256.
-
Gelbard-Sagiv H, Mukamel R, Harel M, Malach R, Fried I (2008) Internally generated reactivation of single neurons in human hippocampus during free recall. Science 322:96-101.
-
Whitlock JR, Heynen AJ, Shuler MG Bear MF (2006) Learning induces long-term potentiation in the hippocampus. Science 313(5790): 1093-1097.
-
Mitsushima D, Sano A, Takahashi T (2013) A cholinergic trigger drives learning-induced plasticity at hippocampal synapses. Nat Commun. 4:2760 doi: 10.1038/ ncomms3760.
-
Mitsushima D, Ishihara K, Sano A, Kessels HW, Takahashi T (2011) Contextual learning requires synaptic AMPA receptor delivery in the hippocampus. Proc Natl Acad Sci USA, 108: 12503-12508
-
Bliss TV, Lømo T (1973) Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J Physiol 232(2) :331–356.
-
Hayashi Y, Shi SH, Esteban JA, Piccini A, Poncer JC, et al. (2000) Driving AMPA receptors into synapses by LTP and CaMKII: requirement for GluR1 and PDZ domain interaction. Science 287(5461): 2262-2267.
-
Takemoto K, et al. (2017) Optical inactivation of synaptic AMPA receptors erases fear memory. Nat Biotechnol. 35: 38-47. PFD
-
Townsend, et al. (2016) α7-nAChR agonist enhances neural plasticity in the hippocampus via a GABAergic circuit. J Neurophysiol 116(6): 2663-2675.
-
Chen SX, Kim AN, Peters AJ, Komiyama T (2015) Subtype-specific plasticity of inhibitory circuits in motor cortex during motor learning. Nat Neurosci. 18: 1109-1115.
-
Fisahn A, Pike FG, Buhl EH, Paulsen O (1998) Cholinergic induction of network oscillations at 40 Hz in the hippocampus in vitro. Nature 394: 186-189.
-
O'Keefe J, Nadel L (1978) The Hippocampus as a Cognitive Map, Oxford University Press pp: 150.
-
Csicsvari J, Hirase H, Czurkó A, Mamiya A, Buzsáki G (1999) Oscillatory coupling of hippocampal pyramidal cells and interneurons in the behaving rat. J Neurosci 19(1): 274-287.
-
Shannon CE (1948) A mathematical theory of communication. Bell System Technical Journal 27: 379–423, 623-656.
-
Hartley RVL (1928) Transmission of Information. Bell System Technical Journal 7(3): 535–563.
-
Ono K, Mitsushima D (2017) Learning creates diversity of excitatory and inhibitory synapses in the hippocampal CA1: a possible amount of information at a single synapse. J Physiol Sci. 67 suppl 1
-
Andersen P, Morris R, Amaral D, Bliss T, & O'Keefe J (2006) The Hippocampus Book. Oxford University Press USA.
-
Kandel ER, Schwartz JH, Jessell TM, Siegelbaum SA. Hudspeth AJ (2013) Principles of Neural Science. Fifth edition, the McGraw-Hill Companies Inc. 2013.
-
Cannon WB (1922) Biographical Memoir: Henry Pickering Bowdich. National Academy of Sciences 17, 181–196.
-
Wiener N (1961) Cybernetics: or control and communication in the animal and machine. MIT press, Cambridge, MA, 2nd Ed.
-
Marr D (1977) Simple memory: a theory for archicortex. Philos Trans R Soc B Biol Sci 262(841): 23-81.
-
Watson JD, Crick FHC (1953) Genetical implications of the structure of deoxyribonucleic acid. Nature 171: 964-967.
-
Nirenberg M, Leder P (1964) RNA code words and protein synthesis. Science 145: 1399-1407.
-
Collins FS (2004) Finishing the euchromatic sequence of the human genome. Nature 431: 931-945.
-
Weaver IC, Cervoni N, Champagne FA, D'Alessio AC, Sharma S, et al. (2004) Epigenetic programming by maternal behavior. Nature Neuroscience 7(8): 847- 854.
-
Ishikawa J, Mitsushima D (2016) Real-time change of neural activity in hippocampal CA1 after the experienced episodes: restraint stress and first encounters with female, male, and object. Soc. Neurosci Abstr. San Diego,
-
Girardeau G, Benchenane K, Wiener SI, Buzsáki G, Zugaro MB (2009) Selective suppression hippocampal ripples impairs spatial memory. Nat Neurosci 12(10): 1222–1223,
- A Review of Gene Therapy for Parkinson's Disease to Control Dopaminergic Neurons
- Late-Onset Myasthenia Gravis in a Patient with Recurrent Breast Cancer: A Case Report
- Covid-Induced Dystonia and Opsoclonus: A Case Report
- Generalized Tonic-Clonic Seizure in a Pediatric Patient with Sunflower Syndrome: A Case Report
- Comparison of Doppler Guided Seldinger Technique Versus Classic Palpatory Seldinger Technique for Radial Artery Cannulation-an Open Label Randomized Controlled Trial
- Brown Sequard Syndrome: Understanding the Complexities of Spinal Cord Injury