A Review of Gene Therapy for Parkinson's Disease to Control Dopaminergic Neurons
Parkinson's disease (PD) is the most common neurodegenerative disorder after Alzheimer's disease. Both motor and nonmotor symptoms are caused by the gradual death of dopaminergic neurones in the substantia nigra and a reduction in dopamine (DA) levels. Early-phase clinical trials have shown that DA replacement gene treatments that raise DA transmitter levels, such as Adeno-associated virus (AAV)–glutamic acid decarboxylase (GAD) and L-amino acid decarboxylase (AADC) gene therapies, are safe and effective. Strategies for disease modification, which try to reduce the course of the disease, seem to be effective. These include treatments that target neurotrophic factors, downstream pathways, and midbrain dopaminergic neuronal factors, all of which have demonstrated promise in preclinical and clinical studies. Other possible gene treatments that target both glial cells and dopaminergic neurones may have similar results, but further study is required. The potential of neurotrophic factors, including Neurturin, Glial-cell-line-derived neurotrophic factor (GDNF), brain-derived neurotrophic factor (BDNF) and vascular endothelial growth factor (VEGF), to promote dopaminergic neurone survival is also being studied. Furthermore, intriguing approaches include gene treatments that target important downstream processes such endoplasmic reticulum (ER) stress, mitochondrial function, and the autophagy–lysosome pathway. As gene editing and delivery methods advance, new avenues for creating potent gene treatments for Parkinson's disease become available. A possible treatment for Parkinson's disease appears to be the CRISPR-Cas9 system, a ground-breaking technology developed in the past ten years that enables precise and instantaneous genome editing in almost any living species.
Abbreviations
PD: Parkinson’s Disease; AAV: Adeno-Associated Virus; GAD: Glutamic Acid Decarboxylase; AADC: L-Amino Acid Decarboxylase; GDNF: Glial-Cell-Line-Derived Neurotrophic Factor; BDNF: Brain-Derived Neurotrophic Factor; VEGF:
Vascular Endothelial Growth Factor; ER: Endoplasmic Reticulum; RBD: Rapid Eye Movement Sleep Behaviour Disorder; SAA: Synuclein Amplification Assays; ICD: Impulse Control Issues; LP: Lewy Pathology; SNPC: Substantia Nigra Pars Compacta; TD: Tremor Dominant; AR: Akinetic Rigid; PIGD: Postural Instability And Gait Disorder; SNc: Substantia
Nigra Pars Compacta; BG: Basal Ganglia; SPNs: Spiny Projection Neurones; GPi: Globus Pallidus; DBS: Deep Brain Stimulation; GAD: Glutamic Acid Decarboxylase; TH: Tyrosine Hydroxylase; GDNF: Glial Cell-Derived Neurotrophic Factor; BBB: Blood-Brain Barrier; UPR: Unfolded Protein Response; ER: Endoplasmic Reticulum; SNr: Substantia Nigra Pars Reticulata; D2-SPNs: D2-Receptor-Expressing Indirect Pathway Neurones.
Introduction
According to the most current Global Burden of Disease Study, Parkinson’s disease (PD) is one of the fastest-growing neurological disorders in terms of prevalence, disability, and death. Notably, Parkinson’s disease (PD) has a sluggish progression and a deceptive start. According to the revised International Parkinson and Movement Disorder Society research criteria for prodromal PD4, prodromal symptoms can be used to identify those who are at risk of developing Parkinson’s disease during this era. The prodromal phase is characterised by rapid eye movement sleep behaviour disorder (RBD), depression, olfactory loss, mild motor symptoms, and abnormal imaging markers of the cardiac sympathetic and presynaptic dopaminergic systems [1].
Parkinson’s disease (PD) is characterised by a complex interaction of genetic, behavioural, and environmental factors. Because heredity only accounts for 5-10% of Parkinson’s disease cases, the majority of persons who have been diagnosed have no family history of the condition [1].
Farming-related Parkinson’s disease risk has received a great deal of research, particularly in regard to pesticide exposure, which has low-dose cocktail effects. The type of crops and livestock used has a significant impact on pesticide and other agricultural exposures. Because of the great range of farming operations, a lot of recent studies suggest that it is critical to examine some farming techniques that might serve as a stand-in for diverse agricultural exposomes. However, as far as we are aware, there aren’t many studies looking at the association between PD risk and distinct farming techniques; most of them merely look at four to eighteen different types of crops and animal husbandry [2].
Synuclein amplification assays (SAA) may quantify αS in biological fluids, allowing for evaluation of pathophysiological mechanisms in “at risk” populations and Parkinson’s disease patients [3].
When PD patients are ON-medication (hyperdopaminergic), they may feel euphoria, well-being, impulse control issues (ICD), hypomania, and psychosis, but when they are OFF-medication (hypodopaminergic), they may experience worry, melancholy, weariness, lack of energy, and motivation. Neurotransmitters other than dopamine, such as serotonin, norepinephrine, acetylcholine, and adenosine, can also impact a variety of neuropsychiatric disorders. Non-motor symptoms frequently change independently and simultaneously with motor variations [4].
In the majority of areas, more than 95% of persons affected have idiopathic Parkinson’s disease. One of the most significant barriers to understanding and treating IPD is its biochemical heterogeneity. Although α-synuclein positive Lewy pathology (LP) promotes degeneration of dopaminergic neurones in the substantia nigra pars compacta (SNPC) in all IPD patients, the disease demonstrates significant clinicopathological variability. Affected persons exhibit varied constellations of motor and non-motor symptoms with varying rates of progress due to extensive neuronal dysfunction and loss of diverse anatomical distribution and severity [3, 5].
This phenotypic variation has fuelled the notion that IPD might be divided into numerous distinct disorders, each with its own underlying mechanisms and potential treatment responses. Previous attempts to differentiate IPD subtypes have been focused on clinical parameters, the most common of which are age of onset and motor or cognitive phenotype. Several phenotypical subtypes have been defined based on these categorisation criteria, including PD dementia (PDD), tremor dominant (TD), akinetic rigid (AR), and postural instability and gait disorder (PIGD). These divisions, however, give little information on the pathogenic heterogeneity of IPD. In reality, top-down tactics that attempt to link consensual phenotypic categories to specific underlying biological processes have yet to be successful, suggesting that alternative strategies are necessary. It has been proposed that a bottom-up strategy, which stratifies IPD based on distinct biological traits rather than clinical profiles, might be more successful in discovering disease subgroups that react well to certain therapies [5].
Etiology of PD
The major etiology of Parkinson’s disease is the loss of nigrostriatal dopaminergic innervation owing to neurodegeneration in the substantia nigra pars compacta (SNc). The pathogenic pathways include α-synuclein aggregates, mitochondrial dysfunction, oxidative stress, and neuroinflammation. Regardless of the specific cause, motor dysfunction or cognitive abnormalities in Parkinson’s disease develop when basal ganglia (BG) neuronal networks cease to operate correctly owing to dopamine depletion.
Dopamine is an important neuronal modulator that regulates the activity of spiny projection neurones (SPNs) in the striatum, the BG’s primary input component. There are two types of SPNs: D1-receptor-expressing direct pathway neurones (D1-SPNs), which project axons to BG output regions such as the internal segment of the globus pallidus (GPi) and the substantia nigra pars reticulata (SNr), and D2- receptor-expressing indirect pathway neurones (D2-SPNs), which regulate BG output via a multisynaptic pathway. Under normal physiological settings, empirical data shows that dopamine reduces D2-SPN activity while boosting D1- SPN activation. Following dopamine depletion, there is an imbalance in the activity of D1- and D2-SPNs, which leads to increased thalamic inhibition. This imbalance is thought to be the key contributor to the motor symptoms seen in Parkinson’s disease [6].
Replacement of Dopamine in the Midbrain Gene Therapy In some cases, healthcare providers administer dopamine receptor agonists. Deep brain stimulation (DBS) is considered after a comprehensive review for persons in advanced stages where traditional medication therapy has proven futile. While DBS has shown a significant increase in motor scores, ranging from 50% to 60%, it requires invasive surgical procedures and the long-term implantation of electrodes into patients’ brains, posing issues such as infection and electrode quality. A unique therapy paradigm uses stem cells to functionally repair the dopaminergic network [6].
Glutamic acid decarboxylase (GAD) catalyses the conversion of glutamate into GABA at human synapses, regulating excitability. In Parkinson’s disease, the GABA pathway, which suppresses hyperexcitability in the midbrain (including the substantia nigra), is compromised, resulting in the malfunction of nigrostriatal dopaminergic neurones, changes in associated brain circuitry, and, finally, movement impairment [7].
Downregulation of GAD1 gene expression has been detected in neurones of Parkinson’s disease patients. As a result, boosting GABA via gene therapy was thought to offer therapeutic promise. AAV-GAD treatment, which supplements the enzyme responsible for GABA production, was the first gene therapy to show efficacy. The first phase 1 clinical study of the AAV-GAD therapy took place between 2005 and 2007. In follow-up investigations, AAV-GAD gene therapy caused significant motor improvements in Parkinson’s disease patients, which were sustained for more than a year. The modifications included a distinct functional connectivity shift, indicating the creation of new connections between subthalamic nuclei and motor cortical areas, which was only seen in individuals who underwent GAD gene therapy [7, 8].
Dopamine is generated from the essential amino acid l-tyrosin in two processes, the tyrosine hydroxylase (TH) enzyme hydroxylates to l-dopa (l-3,4- dihydroxyphenylalanine), which is subsequently converted to dopamine by aromatic l-amino acid decarboxylase (AADC) [9].
In this issue of Science Translational Medicine, report the results of a preclinical study on a novel gene therapy approach that aims to restore the dopamine synthesizing machinery in parkinsonian monkeys [9].
Jarraya et al. developed a gene therapy protocol that involves introducing genes encoding key dopamine biosynthetic enzymes directly into the brain. This creates an artificial dopamine factory in neurones of the striatum, which receives the majority of dopamine released by the substantia nigra [8].
Gene Therapy for the Midbrain Neurotrophic Factor
GDNF
Glial cell-derived neurotrophic factor (GDNF) is crucial for the survival, maintenance, and regeneration of particular neuronal populations in adult brains [10]. This neurotrophic factor has shown favourable benefits in several animal models of Parkinson’s disease, but not in the α-synuclein model [11].
Glial cell-derived neurotrophic factor (GDNF) was first extracted from the supernatant of a rat glioma cell line and discovered to have a significant influence on the survival of midbrain dopaminergic neurones. In terms of possible therapy for neurodegenerative illnesses, GDNF has a relatively high specificity for dopaminergic neurones, making it a promising candidate for treating Parkinson’s disease, which is characterised by gradual depletion of dopaminergic cell populations in the midbrain [12]. Glial cells, such as microglia and astrocytes, release GDNF, a key neurotrophic factor. GDNF regulates neuroinflammation and is a therapeutic target for several neurological disorders [13]. Despite the limited therapeutic effect, the introduction of the GDNF family neurotrophic factor neurturin (NRTN) via an AAV2 vector was shown to be safe and well tolerated in PD patients [11, 14].
The existence of the blood-brain barrier (BBB) makes transport to the brain challenging, limiting its utility. To address this issue, we took use of the fact that bone marrow stem cell-derived macrophages may cross the BBB and get to areas of neuronal degeneration. Here, we describe the invention of a technique for brain delivery of GDNF using genetically engineered macrophages [15].
Neurturin
It is a natural structural and functional counterpart of GDNF. They also shed light on impairments in the condition of nigrostriatal neurones in advanced Parkinson’s disease, suggesting that severe axon-transport deficits restricted AAV2-NRTN bioactivity by restricting the protein accessible to the cell body. Thus, future efforts to treat neurodegenerative disorders with neurotrophic factors will need to target both the terminal fields and the cell bodies of degenerating neurones in order to maximise benefit [16]. The patients received AAV2-NRTN (CERE-120) injections bilaterally, stereotactically, and intraputaminal [17].
Cerebral Dopamine Neurotrophic Factor (CDNF)
CDNF modulates stress-induced unfolded protein response (UPR) signalling in the endoplasmic reticulum (ER) and supports protein homeostasis there. Because ER stress can produce dopaminergic degeneration in Parkinson’s disease, CDNF may be a therapeutic target [7].
CDNF protects and repairs dopamine neurones in Parkinson’s disease models in rodents and nonhuman primates. CDNF was evaluated on Parkinson’s patients in a Phase I-II experiment, and the research fulfilled its key goals of safety and tolerability after 12 months (Clinical trial number NCT03295786). CDNF has previously undergone broad toxicity assessments in non-human primates, and it has been evaluated satisfactorily in a Phase I-II clinical study in Parkinson’s patients. Importantly, CDNF’s long- lasting downregulation of Unfolded Protein Response (UPR) markers in the SOD1-G93A and TDP43 animal models, even after a single injection, suggests that the protective impact lasts a long time, indicating its potential therapeutic utility [18].
Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF)
In a PD animal model, MANF protects and repairs dopamine (DA) neurones. MANF is increased in response to ER stress, circumventing the overall reduction of protein synthesis. MANF protects ER-stressed cells and reduces UPR in several in vitro studies [19].
In rats with PD produced by 6-hydroxydopamine (6-OHDA), single doses of CDNF (Lindholm, et al., 2007) and MANF (Voutilainen et al., 2009) preserve and repair dopaminergic neurones [20].
MANF inhibited dopaminergic neurodegeneration induced by microglia activation in Parkinson’s disease via activating the AKT/GSK3β-Nrf-2 signalling pathway [21].
Brain-Derived Neurotrophic Factor (BDNF)
Neurogenesis is regulated by the brain-derived neurotrophic factor (BDNF) [22]. In situ hybridisation showed that dopaminergic neurones in the control substantia nigra pars compacta (SNpc) exhibit high levels of BDNF mRNA [23]. In MPTP-induced PD mice, BDNF overexpression reduced cognitive impairment and motor impairments. Following stereotaxic injection of BDNF-AAV, the brain’s BDNF protein levels dramatically rose [24].
Vascular Endothelial Growth Factor (VEGF)
The VEGF encapsulated nanospheres (NS) can go through the blood-brain barrier and shield the DA neurones [25]. VEGF may have analgesic effects because it suppresses the production of transient receptor potential vanilloid 1 (TRPV1). Thus, the advantages of VEGF-B and -C are supported by therapeutic data [26].
Gene Editing Therapy using CRISPR-Cas9
The CRISPR-Cas system was discovered in 1987 when it was discovered that bacteria, whenever they came into contact with phage DNA, inserted 32-nt (nucleotide) spacer sequences into 29-nt repeat sequences at CRISPR loci [27].
In CRISPR-Cas, there are two classes (Class 1 and Class 2), six kinds (I–VI) and 33 subtypes are distinguished [28]. Class 1 comprises multi-subunit effector complexes, whereas Class 2 comprises single protein effector modules. Additional study and analysis of the Class 2 CRISPR-Cas system was prompted by the discovery of two additional types and many subtypes. Of the two newly discovered and described CRISPR types, only the type VI systems were able to target RNA. One special characteristic of class 2 systems is that in some situations, the effector protein also participates in the processing of pre-crRNA (CRISPR RNA) [29].
Basic biological research is moving more quickly because to CRISPR-Cas9 technology, which enables accurate and efficient gene editing in practically every type of cell and creature. More and more teams are concentrating on identifying the molecular causes of neurological conditions and creating innovative treatments with the CRISPR-Cas9 technology [30].
A possible treatment for Parkinson’s disease appears to be the CRISPR-Cas9 system, a ground-breaking technology developed in the past ten years that enables precise and instantaneous genome editing in almost any living species. According to a number of recent papers, CRISPR-Cas9 presents the potential to expedite fundamental research, with an emphasis on clarifying the pathogenicity of neurological illnesses and generating novel treatments, namely for Parkinson’s disease. CRISPR-Cas9 technology is becoming more and more popular because it is more concise, adaptable, and economical than previous gene-editing techniques. By editing potential genes to produce suitable animal and cell line models, the CRISPR-Cas9 method greatly advances our knowledge of the illness. It could eventually be a useful and successful tool for gene therapy, which is thought to be a novel PD treatment approach (Figure 1) [27].
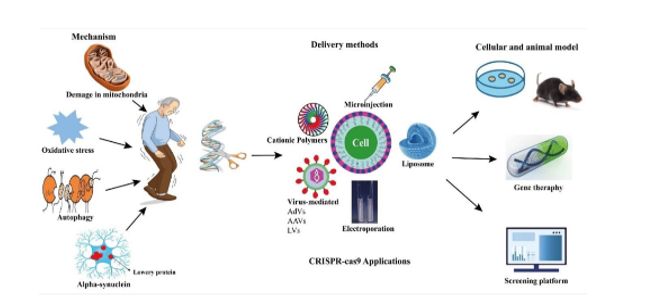
The two types of Parkinson’s disease are familial and sporadic; the latter accounts for only 10–15% of cases. Gene mutations in SNCA, Parkin, PINK1, DJ-1, and LRRK2 are the cause of the familial types. Autosomal-dominant types of Parkinson’s disease are caused by mutations in SNCA and LRRK2. In recent years, PD research has made extensive use of CRISPR-Cas9 technology to alter, knock out, or knock in genes associated with the illness. PD cellular models have mostly been created through in vitro research. To introduce the G2019S mutation in the LRRK2 gene, for instance, Vermilyea and colleagues used CRISPR-Cas9 on induced pluripotent stem cells (iPSCs) derived from marmosets, a non- human primate. They observed increased ROS generation, reduced neuronal survival, and neurite complexity—all characteristics linked to Parkinson’s disease—after the iPSCs were differentiated into dopaminergic neurones. Using human embryonic stem cells, CRISPR-Cas9 has also been successfully used to create in vitro models of Parkinson’s disease. In vivo models of Parkinson’s disease have also been created using CRISPR-Cas9. For example, CRISPR-Cas9- mediated genome editing produced Vps35 D620N knock-in mice, which exhibit certain important PD traits. In separate research, a conventional CRISPR-Cas9 system administered via AAV9 successfully knocked out the PINK1 and DJ-1 genes simultaneously in the monkeys’ substantia nigra. This method resulted in the development of behavioural and neuropathological changes, among other important characteristics of Parkinson’s disease [31, 32].
Conclusion
Since Parkinson’s disease (PD) mostly affects tiny, targeted brain regions, gene therapy may be an appropriate treatment option. By precisely delivering therapeutic genes to the afflicted areas, targeted treatments may be able to restore normal function or reduce the course of the disease. With the goal of altering the progression of the disease, a large number of gene therapy studies for Parkinson’s disease have already been carried out and many more are in progress. For the therapeutic treatment of Parkinson’s disease, the majority of gene treatments have not yet received approval. Inconsistencies in treatment effects and claimed side effects have been the cause of several failures.
“Restoring the DA pathway” is the ideal objective of PD therapy. Only certain gene treatments addressing midbrain factors are likely to do this, in addition to cell replacement therapy. Future treatment prospects include gene therapy using neurotrophic proteins including GDNF, BDNF, MANF, CDNF, and VEGF, as well as midbrain-specific developmental factors like Nurr1, Foxa2, and Pitx3. This strategy will become increasingly popular in the upcoming years as AAV vectors for gene therapy have been approved in both the US and Europe. When compared to adenoviral and lentiviral vectors, some AAV serotypes are favoured as therapeutic vectors due to their favourable safety profiles, robust cerebral tropism, effective and long-lasting induction of foreign gene expression, and minimal immunogenicity in the brain. Treatments involving persistent gene transfer, however, need careful consideration of the risk-benefit ratio. Notwithstanding the dangers, the absence of therapies that potentially halt Parkinson’s disease development suggests a substantial unmet therapeutic need. Key findings from preclinical and clinical trials—some of which were unsuccessful but might help shape the future of gene therapies for the treatment and management of CNS disorders—were emphasised in this review. Gene therapy might be utilised to meet the demands of specific patients in addition to the present therapeutic treatment paradigms that are pharmacologically based.
Acknowledgments
We would like to express our heartful gratitude to CMR Institute of Pharmacy. We are also thankful for staff who provided technical assistance and necessary background data during paper work.
Conflict of Interest
All the authors declared that there is no conflict of interest.
References
-
Chen X, Yaqi L, Yun S, Michael A, Xiang G (2025) Prodromal Parkinson’s disease and subsequent risk of Parkinson’s disease and mortality. npj Parkinson’s Disease 11(14): 8.
-
Pascal P, François B, Vincent B, Nicolas V (2025) Investigating Parkinson’s disease risk across farming activities using data mining and large-scale administrative health data. npj Parkinson’s Disease 11(13): 11.
-
Amgad D, Avital YV, Daniel A, Kyle BF, Omar SM (2025) Radiological markers of CSF α-synuclein aggregation in Parkinson’s disease patients. npj parkinson’s disease 11(7): 9.
-
Mari M, Emmanuelle S, Roberto I, Andrea K, Valérie F, et al. (2024) Deciphering the effects of STN DBS on neuropsychiatric fluctuations in Parkinson’s disease. npj parkinson’s disease 10(205): 6.
-
Irene HF, Lilah T, Dagny AS, Martina C, Sepideh M (2024) Mitochondrial complex I deficiency stratifies idiopathic Parkinson’s disease. nature communications 15: 18.
-
Xunyi Y, Xin J (2024) Shedding light on gene therapy of Parkinson’s disease in non-human primates Innovation (Camb) 5(2): 100581.
-
Jintae K, Yoon C (2024) Gene Therapy for Parkinson’s Disease Using Midbrain Developmental Genes to Regulate Dopaminergic Neuronal Maintenance Int J Mol Sci 25(22): 12369.
-
Bechir J, Sabrina B, Scott RG, Caroline J, Gilles B, et al. (2009) Dopamine Gene Therapy for Parkinson’s disease in a Nonhuman Primate without Associated Dyskinesia Science Translational Medicine 1(2).
-
Tomas B, Deniz K (2009) Gene therapy for dopamine replacement in Parkinson disease 1(2): 2ps2.
-
Shelley JA, Judy JW, Deborah KS, Neil UB, Nikunj KP (2013) GDNF, NGF and BDNF as therapeutic options for neurodegeneration. Pharmacology & Therapeutics 138(2): 155-175.
-
Susanne B, Johan P, Emilia G, Paivi P, Liina LN, et al. (2013) Gene therapy with AAV2-CDNF provides functional benefits in a rat model of Parkinson’s disease. brain and behaviour 15(2): 75-78.
-
Yousra EO, Jasper B, Barbara W, Nathalie C, Inez W (2023) Neurotrophic Factors as Regenerative Therapy for Neurodegenerative Diseases: Current Status, Challenges and Future Perspectives. Int J Mol Sci 24(4): 3866.
-
Gurpreet S, Anupama S, Vivek P, Saurabh S, Shashi BS (2023) Therapeutic potential of GDNF in neuroinflammation: Targeted delivery approaches for precision treatment in neurological diseases. Journal of Drug Delivery Science and Technology 87: 104876.
-
Amber L, Chadwick C, Aristide M, Nicolas P, Bradley E, et al. (2024) Phase 1b Safety and Preliminary Efficacy of Bilateral Intraputaminal Delivery of AAV2 GDNF (AB- 1005) in Participants with Mild or Moderate Parkinson’s Disease (N2.001). Neurology 102(17S).
-
Kc B, Qing Z, Guiming L, Syed ZI, James LR, et al. (2010) Macrophage-mediated GDNF delivery protects against dopaminergic neurodegeneration: a therapeutic strategy for Parkinson’s disease. Mol Ther 18(8): 1536-1544.
-
Raymond TB, Christopher DH, Yaping C, Alistair W (2011) Lamar Brown Bioactivity of AAV2-neurturin gene therapy (CERE-120): Differences between Parkinson’s disease and nonhuman primate brains 26(1): 27-36.
-
Christopher DH, Biplob D, Mehdi G, Kathie B, Jeffrey HK, et al. (2008) Transgene Expression, Bioactivity, and Safety of CERE-120 (AAV2-Neurturin) Following Delivery to the Monkey Striatum. molecular therapy 16(10): 1737-1744.
-
Francesca DL, Patrick L, Jinhan N, Liam B, Federica P, et al. (2023) CDNF rescues motor neurons in models of amyotrophic lateral sclerosis by targeting endoplasmic reticulum stress. Brain 146(9): 3783-3799.
-
Vera K, Ying YL, Larisa I, Paivi L, Mati K (2024) MANF regulates neuronal survival and UPR through its ER- located receptor IRE1α. cell reports 42(2): 112066.
-
Merja HV, Susanne B, Johan P, Paivi L, Atso R, et al. (2011) Chronic infusion of CDNF prevents 6-OHDA- induced deficits in a rat model of Parkinson’s disease. Experimental Neurology 228(1): 99-108.
-
Jing XZ, Kai GZ, Yan XY, Ling JJ, Wei FT, et al. (2023) Mesencephalic astrocyte-derived neurotrophic factor (MANF) prevents the neuroinflammation induced dopaminergic neurodegeneration. Experimental Gerontology 171: 112037.
-
Helen S, Jeffrey G, Adam M, Sudar P, Cara A (2005) Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Experimental Neurology 192(2): 348-356.
-
Howells DW, Porritt MJ, Wong JYF, Batchelor PE, Kalnins R, et al. (2000) Reduced BDNF mRNA Expression in the Parkinson’s disease Substantia Nigra. Experimental Neurology 166(1): 127-135.
-
Chang E, Jiongmei W (2021) Brain-derived neurotrophic factor attenuates cognitive impairment and motor deficits in a mouse model of Parkinson’s disease. brain and behavior 11(8): e2251.
-
Xian YM, Huang AQ, Afsar K, Zhang L, Xiao QS, et al. (2020) vascular endothelial growth factor-loaded poly- lactic-co-glycolic acid nanoparticles with controlled release protect the dopaminergic neurons in Parkinson’s rats. Chemical biology & drug design 95(6): 631-639.
-
Torsten F, Xu Y, Shiling Z, Alexander DCM, Brandon JY, et al.(2011) Vascular endothelial growth factor-B is neuroprotective in an in vivo rat model of Parkinson’s disease. Neuroscience Letters 496(1): 43-47.
-
Mujeeb UR, Muhammad B, Junaid AS, Ajeet K, Pierre LT, et al.(2022) CRISPR-Cas9-Based Technology and Its Relevance to Gene Editing in Parkinson’s Disease. Pharmaceutics 14(6): 1252.
-
Kira SM, Yuri I. Wolf, Jaime Iranzo, Sergey A. Shmakov, Omer S. Alkhnbashi, et al. (2019) Evolutionary classification of CRISPR–Cas systems: a burst of class 2 and derived variants. Nature Reviews Microbiology 18: 67–83.
-
Eugene VK, Kira SM, Feng Z (2017) Diversity, classification and evolution of CRISPR-Cas systems. Current Opinion in Microbiology 37: 67-78.
-
Lihong G, Yawei H, Ciqing Y, Suxiang L, Jiang D, et al. (2022) CRISPR-Cas9-Mediated Gene Therapy in Neurological Disorders. Molecular Neurobiology 59: 968-982.
-
Jintae K, Yoon CM (2024) Gene Therapy for Parkinson’s Disease Using Midbrain Developmental Genes to Regulate Dopaminergic Neuronal Maintenance. Int J Mol Sci 25(22): 12369.
-
Kuo HC, Cheng YH, Chih HOY, Chang HH, Han YL, et al. (2021) In vitro genome editing rescues Parkinsonism phenotypes in induced pluripotent stem cells-derived dopaminergic neurons carrying LRRK2 p.G2019S mutation. Stem Cell Research & Therapy 12(508).
- Late-Onset Myasthenia Gravis in a Patient with Recurrent Breast Cancer: A Case Report
- Covid-Induced Dystonia and Opsoclonus: A Case Report
- Generalized Tonic-Clonic Seizure in a Pediatric Patient with Sunflower Syndrome: A Case Report
- Comparison of Doppler Guided Seldinger Technique Versus Classic Palpatory Seldinger Technique for Radial Artery Cannulation-an Open Label Randomized Controlled Trial
- Brown Sequard Syndrome: Understanding the Complexities of Spinal Cord Injury
- Seepage and Swelling-Neuroendocrine Tumour Testis