Production of Corn; Effects of Manganese Application on Plant Parameters
Corn plants need a small amount of manganese (Mn) that plays an active role in plant metabolism. We determined plant parameters, relative water content (RWC), chlorophyll (Chl) content, Chl fluorescence, photosynthesis parameters, and yield parameters to justify the effects of Mn on corn plants. Different Mn concentrations (0, 0.2, 1.5 and 3 ppm) were applied as a foliar spray on leaves of corn plants. The Mn treatment increased plant’s height and leaf numbers. RWC, net photosynthesis rate (Pn) and photosynthetically active radiation (PAR), Chl content and Chl fluorescence increased in leaves of Mn-treated plants compared to that of Mn-untreated plants. In addition, Mn application increased the yield of corn plants. Taken together, the Mn concentration at 1.5 ppm as a foliar application might increase corn production.
Introduction
Corn (Zea mays) is a cereal grain and used as a food source for both humans and animals. Micronutrients improve the yield and the crop quality for cereals, corn, beans, forages, and oil seeds [1]. The deficiency of micronutrients reduces performance and profitability in the plant [2]. Also, Maize is a major cereal crop in the world and has economic value in livestock [3]. The Mn deficiency reduced dry matter production and yield but induced susceptibility to drought and heat stress that produces pale green or yellow patches on younger leaves [4]. Manganese (Mn) involves in plant metabolic processes such as respiration, photosynthesis, amino acid synthesis and hormone activation [5, 6]. It was reported that Mn is an essential mineral micronutrient and co-factors for antioxidant enzymes [7]. Physiological activities such as enzyme activity, absorption, translocation and oxidative stress might occur due to excessive of the Mn concentration in plant tissue [8]. Manganese plays a key role in photosynthesis and photolysis (light splitting) of water molecules and provides energy for photosynthesis. Therefore, Mn deficiency impairs photosynthesis [9] and lignin biosynthesis in the root of plants, which increase soil- born fungi following the lignin-induced reduction of pathogenic infection [10]. The application of Mn provides the resistant capacity in plants against not only various soil-borne diseases but also fungal leaf diseases [11, 12]. Manganese deficiency also weakens fatty acid production, which induces non-stomatal water loss and the heat load on leaves and increased transpirational water loss and lower water-use-efficiency [13]. To date, few pieces of research were accounted on micronutrients function on corn plants [14, 15, 16, 17, 18]. However, the effects of Mn on the physiological parameters of corn plants and yield were not properly justified under low fertile soil condition. Therefore, this study was taken to know whether Mn improves in improving plant physiological functions and sustaining yield of the corn plant in sandy soil.
Methodology
Hybrid corn variety of L41 was used in this study. Two seeds were placed in a hole on a pre-prepared seedbed and the spacing of 25 cm X 75 cm in between two holes was applied. After that, a healthy seedling was maintained to grow for experimental purposes. The soil was a predominantly of BRIS soil that contains more than 90% of a sand particle with reduced soil physical and chemical properties. The BRIS soil encompasses carbon (3.82%), nitrogen (0.14%), phosphorus (0.1%), potassium (0.03%), calcium (0.34%), magnesium (1.01%), manganese (0.008%) and pH of 4.5. The compost, contains of carbon (27.5%), nitrogen (1.7%), phosphorus (1.5%), potassium (1.0%), calcium (2.3%), magnesium (1.3%), manganese (0.04%) and pH of 6.5, was mixed with BRIS soil. Plant tissue contains 35.2 mg kg-1 DM of Mn. Four Mn (0, 0.2, 1.5 and 3 ppm) treatments with five replicates were arranged according to the completely randomized design. Care of plants [19, 20, 21] and different cultural practices were followed according to the previous studies [22]. The concentration of Mn was chosen based on the lower, optimum and maximum level according to previous information [23]. The leaves were counted numerically. The plant height was measured from the soil surface to the leaf tip of the top of the final leaf emerged with a measuring ruler alongside. The weight of fresh leaf (FW) was measured just after detached from the plants then turgid weight (TW) was determined after leaf was incubated in distilled water for 24 h to obtain a full turgidity. Dry weight (DW) of the leaf, dried at 60°C for 24 h in an oven, was measured. The following reaction was used to measure relative water content according to the previous studies [24, 25, 26]. RWC (%) = [(FW-DW) / (TW-DW)] x 100 An SPAD-502 portable chlorophyll meter (Minolta, Japan) was used to acquire a rapid estimation of chlorophyll (Chl) content in leaves of corn plants [27, 28, 29]. Five plants were selected and the second uppermost collared-leaf was used to determine Chl content. There were five measurements from each plant and a total of 25 measurements were taken for each treatment. Data were taken at 11 am up to 1 pm to avoid wetness condition that affects Chl reading. A portable Junior-PAM Chl-monitoring meter (Walz, Germany) was used to quantify Chl fluorescence from leaves of corn plants [26]. A second uppermost collared-leaf was used for taking the data in between 11 am to 1 pm. The maximum fluorescence level was recorded in this experiment.Net photosynthesis rate (Pn) was measured using a CI-340 portable photosynthesis meter (CID Biosciences, Inc.) according to Khairi et al. [26]. These data were taken in between 11 am to 1 pm. Five measurements were taken for each treatment. Plant height, the number of leaves, length and weight of cob were determined after the yield produced. Cob weight was taken by using digital weight machine for each treatment. Data were analyzed for differences of mean value among treatments by one factor ANOVA procedure and T-test using Minitab-16 and MS Excel software. Differences at P < 0.05 were considered significant.
Results and Discussion
Plant height and leaf numbers were presented in Figure 1.
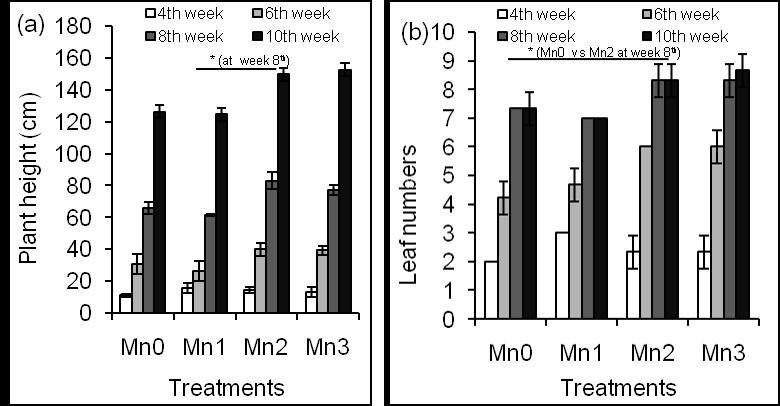
Figure1: Effects of different concentrations of Mn on plant height (a) and leaf numbers (b).
Weekly-data showed that the height of corn plant increased with increasing time (Figure 1a). At week 4th, Mn did not affect plant height in both of Mn-treated and Mn-untreated corn plants. Plant height between control (Mn0) and a lower dose of Mn (Mn1) was almost similar throughout the growing period. On the other hand, Mn2 treatment significantly increased plant height compared to Mn1 treatment at week sixth (P ≤ 0.003) and week eighth (P ≤ 0.001) where control and Mn1 showed no difference. Similar results were observed in the case of Mn3 treatment. The plant height was similar when data were compared between week 6th and week 10th. It suggests that higher concentrations of Mn affect plant height [30]. At week 4th, Mn did not affect leaf numbers in Mn-treated and Mn-untreated plants (Figure 1b).
However, Mn2 significantly increased numbers of the leaf at week 8th (P ≤ 0.016) and week 10th (P ≤ 0.016). The effects of Mn3 on leaf numbers were similar to Mn2 treatments. This result suggests that Mn might increase leaf numbers of corn plant. Relative water content significantly (P ≤ 0.003) decreased in leaves of Mn-untreated plants compared to leaves of Mn-treated plants (Figure 2a).
Relative water content increased with increasing Mn concentration and reached at the highest level when plants were treated with 3 ppm of Mn. The Mn2 treatment significantly (P ≤ 0.0002) increased RWC in plants compared to Mn0 treatment. Also, the RWC declined significantly (P ≤ 0.014) in plants when it was treated with Mn1 (0.2 ppm) compared to the Mn3 (3 ppm) treatment. Taken together, these results indicate that Mn2- treated plants might perform a better function related to the water content in the cell than Mn-untreated plants. Manganese plays a major role in improving stress tolerance through the increasing activity of Mn- superoxide dismutase. Higher RWC in Mn-treated plant improves fatty acid production [31]. Therefore, there is a possible that Mn deficiency plants showed susceptibility to both drought and heat stress. Because, Mn deficiency reduced the waxy content that increased transpirational water loss and lower water-use-efficiency [31].
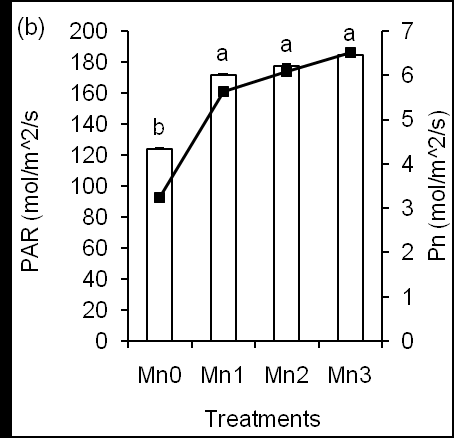
We measured the net Pn and PAR rate in leaves of corn plants under different Mn conditions (Figure 2b). We found that Mn treatment significantly (P ≤ 0.0001) increased Pn rate in plants compared to Mn-untreated plants (Figure 2b, line graph). This result suggests that foliar application of Mn might affect net photosynthesis rate. In addition, Mn1 treatment significantly (P ≤ 0.012) increased PAR in plants than Mn0- treated plants (Figure 2b, bar graphs). In the case of Mn2 and Mn3, Pn and PAR significantly increased in plants than Mn-untreated plants, but Pn and PAR were insignificantly different among different Mn treatments. Taken together, it suggests that Mn increases plant photosynthesis rate and functions on light related reaction, example, light antenna complexes function in photosystem II where Mn is required for water oxidation, which plays a major role in photosynthesis [9]. Mn provides energy during photosynthesis through the splitting of water molecules in photosystem II [32] and damaged the photosynthetic apparatus [33], which finally might affect the production of dry matter and yield in plants. Whether the Mn application affected light related parameters, the chlorophyll (Chl) content, Chl fluorescence in leaves were determined at different growing stages. The Mn treatment affected Chl content (Figure 3a), however, Mn- treated plants presented different Chl accumulations in leaves under different observation times (Figure 3a).
![Figure 3: Effects of different concentrations of Mn on Chl content (a) and Chl florescence (b). In addition, Chl content increased significantly in leaves of Mn-treated plants at week 8th (P ≤ 0.013) and 10th (P ≤ 0.012) than in leaves of Mn- untreated plants. These results suggest that Mn application increases Chl content in leaves of corn plants and support that Mn might function on the light antenna in photosystem II. Previously, Jahan et al. [34,35] stated that light-harvesting antenna of photosystem II increased chlorophyll content in Arabidopsis plants. In plants, Mn participates in the structure of photosynthetic proteins and enzymes [32]. Chl fluorescence is positively correlated with Chl content data. In addition, Mn application increased chlorophyll fluorescence in leaves of corn plants (Figure 3b). Mn- treated plants showed no difference (P ≤ 0.09) concerning Chl fluorescence accumulation during data taking time from 4th to 10th week but significantly higher than Mn- untreated plants (Figure 3b). These results indicate that Mn functions on Chl parameters probably through the photosystem II-water oxidizing system and photolysis of water molecules that provide energy for photosynthesis [9]. This result was consistent with Pn results in this study (Figure 2b). The yield and yield parameters were determined based on the weight and length of corn fruit, which were shown in Figure 4. The yield gradually increased with increasing Mn concentration (Figure 4a).](/fulltextimages/235/fig_3.png)
Figure 3: Effects of different concentrations of Mn on Chl content (a) and Chl florescence (b). In addition, Chl content increased significantly in leaves of Mn-treated plants at week 8th (P ≤ 0.013) and 10th (P ≤ 0.012) than in leaves of Mn- untreated plants. These results suggest that Mn application increases Chl content in leaves of corn plants and support that Mn might function on the light antenna in photosystem II. Previously, Jahan et al. [34, 35] stated that light-harvesting antenna of photosystem II increased chlorophyll content in Arabidopsis plants. In plants, Mn participates in the structure of photosynthetic proteins and enzymes [32]. Chl fluorescence is positively correlated with Chl content data. In addition, Mn application increased chlorophyll fluorescence in leaves of corn plants (Figure 3b). Mn- treated plants showed no difference (P ≤ 0.09) concerning Chl fluorescence accumulation during data taking time from 4th to 10th week but significantly higher than Mn- untreated plants (Figure 3b). These results indicate that Mn functions on Chl parameters probably through the photosystem II-water oxidizing system and photolysis of water molecules that provide energy for photosynthesis [9]. This result was consistent with Pn results in this study (Figure 2b). The yield and yield parameters were determined based on the weight and length of corn fruit, which were shown in Figure 4. The yield gradually increased with increasing Mn concentration (Figure 4a).
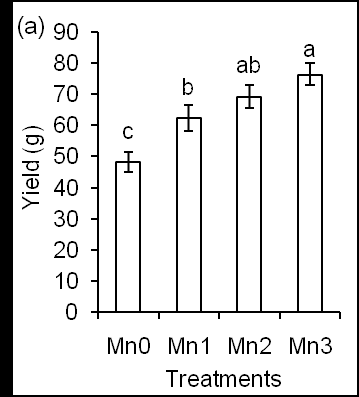
The yield was lower than usual corn production due to the BRIS soil. Yield significantly increased in Mn-treated plants than Mn- untreated plants. In addition, different Mn concentrations differentiate corn yield where a higher concentration of Mn treatment significantly increased (P ≤ 0.013) corn yield compared to the lower Mn concentration treatment. This result was consistent with Pn data (Figure 2b) that higher Pn rate contributes in increasing yield of corn plants. Consequently, Mn2 and Mn3 showed better performance regarding cob yield. Besides, the length of corn plant was also higher under Mn2 and Mn3 treatments (Figure 4b) and showed insignificantly larger size than that of Mn- untreated plants and 0.2 ppm Mn- treated plants. These results also support that 1.5 ppm Mn supports plant tissues for better enzyme activity, absorption, translocation and utilization of other mineral elements [8].In conclusion, this study shows that the Mn application enhanced RWC, PAR, and Pn rate in corn plants. The 1.5 ppm of Mn application increased yield as well as increasing light related parameters, Chl content and Chl fluorescence and photosynthesis rate. Therefore, Mn at 1.5 ppm could be applied as a foliar spray on leaves for sustainable corn production.
Acknowledgements
This work was supported by the faculty of Agriculture, Biotechnology and Food Sciences, Universiti Sultan Zainal Abidin, Kuala Terengganu, Malaysia.
References
-
Malakouti MJ (2007) Zinc is a neglected element in the life cycle of plants: A review. Middle East Russian Journal of Plant Science and Biotechnology 1(1): 1-12.
-
Fisher GEJ (2008) Micronutrients and animal nutrition and the link between the application of micronutrients to crops and animal health. Turkish Journal of Agriculture and Forestry 32(3): 221-233.
-
Harris D, Rashid A, Miraj G, Arif M, Shah H (2007) On- farm seed priming with zinc sulphate solution A cost effective way to increase the maize yields of resource poor farmers. Field Crops Resistance 110(2): 119- 127.
-
Alloway BJ (2008) Micronutrients and Crop Production: An Introduction. In: Alloway BJ (Ed) Micronutrient Deficiencies in Global Crop Production Springer 1-40.
-
Inani N, Nozulaidi M, Khairi M, Abdulkadir AB, Jahan MS (2015) Glutathione functions on physiological characters of corn plants to enhance Mn-induced corn production. Pertanika Journal Tropical Agriculture Sciences 38(4): 509-581.
-
Lidon FC, Barreiro M, Ramalho J (2004) Manganese accumulation in rice: Implications for photosynthetic functioning. Journal of Plant Physiology 161(11): 1235-1244.
-
Cakmak I (2000) Role of zinc in protecting plant cells from reactive oxygen species. New Phytology 146: 185-205.
-
Lei Y, Korpelainen H, Li C (2007) Physiological and biochemical responses to high Mn concentrations in two contrasting Populus cathayana populations. Chemosphere 68(4): 686-694.
-
Hakala M, Rantamaki S, Puputti EM, Tyystjarvi T, Tyystjarvi E (2006) Photoinhibition of manganese enzymes: insights into the mechanism of photosystem II photoinhibition. Journal of Experimental Botany 57(8): 1809-1816.
-
Marschner H (2012) Marschner’s Mineral Nutrition of Higher Plants (3rd ed). Elsevier Science.
-
Heine G, Max JFJ, Fuhrs H, Moran-Puente DW, Heintz D, et al. (2011) Effect of manganese on the resistance of tomato to _Pseudocercospora fuligena_. Journal of Plant Nutrition and Soil Science 174(5): 827-836.
-
Yao Q, Osborne L, Rengel Z (1998) Micronutrient deficiency changes activities of superoxide dismutase and ascorbate peroxidase in tobacco plants. Journal of Plant Nutrition 21(7): 1427-1437.
-
Hebbern CA, Laursen KH, Ladegaars AH, Schmidt SB, Pedas P, et al. (2009) Latent manganese deficiency increases transpiration in barley (_Hordeum vulgare_). Physiologia Plantarum 135(3): 307-316.
-
Xue Y, Yue S, Zhang W, Liu D, Cui Z, et al. (2014) Zinc, Iron, Manganese and Copper Uptake Requirement in Response to Nitrogen Supply and the Increased Grain Yield of Summer Maize. PLoS One 9(4): e93895.
-
Munirah N, Khairi M, Nozulaidi M, Jahan MS (2015) The effects of zinc application on physiology and production of corn plants. Australian journal of basic and applied sciences 9(2): 362-367.
-
Munirah N, Jahan MS, Nashriyah M (2015) N- acetylcysteine and Zn regulate corn yield. Science Asia 41: 246-250.
-
Syuhada N, Jahan MJ (2016) Glutathione functions on physiological characters to increase copper-induced corn production. Russian Agricultural Science 42(1): 5-10.
-
Syuhada N, Jahan MS, Nashriyah M, Khairi M, Nozulaidi M, et al. (2014) Application of copper increased corn yield through enhancing physiological functions. Australian Journal of Basic and Applied Sciences 8(16): 282-286.
-
Sarwar MJ, Khanif YM (2005) Techniques of water saving in rice production in Malaysia. Asian Journal of Plant Sciences 4(1): 83-84.
-
Sarwar MJ, Khanif YM (2005b) The effect of different water levels on rice yield and Cu and Zn concentration. Journal of Agronomy 4(2): 116-121.
-
Sarwar MJ, Khanif YM, Syed Omar SR, Sinniah UR (2004) The effect of different water regimes on yield and bioavailability of Phosphorus in rice production in Malaysia. The Malaysian Journal of Soil Science 8: 53-62.
-
Jahan MS, Khanif YM, Sinniah UR, Nozulaidi MBN, Khairi MBCL (2012) Bioavailability of soil nitrogen in low water input rice production. Journal of Sustainability Science and Management 7(2): 207- 212.
-
Jahan MS, Nordin MNB, CheLah MKB, Khanif YM (2013) Effects of water stress on rice production: bioavailability of potassium in soil. The Journal of Stress Physiology & Biochemistry 9(2): 97-107.
-
Soltangheisi A, Rahman ZA, Ishak CF, Musa HM, Zakikhani H (2014) Interaction effects of zinc and manganese on growth, uptake response and chlorophyll content of sweet corn (_Zea mays_ var. _saccharata_). Asian Journal of Plant Science 13(1): 26- 33.
-
Chelah MKB, Nozulaidi MN, Musliania MI, Khanif YM, Jahan MS (2011) Composting increases BRIS soil health and sustains rice production on BRIS Soil. Science Asia 37: 291-295.
-
Khairi M, Nozulaidi M, Afifah A, Jahan MS (2015) Effect of various water regimes on rice production in lowland irrigation. Australian Journal of Crop Science 9(2): 153-159.
-
Khairi M, Nozulaidi M, Jahan MS (2015) Effects of different water levels on physiology and yield of salinity rice variety. Australian Journal of Basic and Applied Sciences 9(2): 339-345.
-
Khairi M, Nozulaidi M, Jahan MS (2016) Effects of flooding and alternate wetting and drying on the yield performance of upland rice. The Pertanika Journal of Tropical Agricultural Science 39(3): 299-309.
-
Nozulaidi M, Khairi M, Jahan MS (2015) Effects of different salinity levels on rice production. Australian Journal of Basic and Applied Sciences 9(5): 524-530.
-
Gavalas NA, Clark HE (1971) On the role of manganese in photosynthesis; kinetics of photo inhibition in manganese-deficient and 3-(4- Chlorophenyl)-1, 1-dimethylurea-inhibited Euglena gracilis. Plant Physiology 47(1): 139-143.
-
Buchanan B, Grusen W, Jones R (2000) Biochemistry and molecular biology of plants. Amer Soc Plant Physiol Maryland 1367.
-
Mukhopadhyay M, Sharma A (1991) Manganese in cell metabolism of higher plants. Botany Review 57(2): 117-149.
-
Jahan MS, Nozulaidi M, Khairi M, Mat N (2016) Light- harvesting complexes in photosystem II regulate glutathione-induced sensitivity of Arabidopsis guard cells to abscisic acid. Journal Plant Physiology 195: 1- 8.
-
Jahan MS, Nozulaidi M, Khandaker MM, Afifah A, Husna N (2014) Control of plant growth and water loss by a lack of light-harvesting complexes in photosystem-II in _Arabidopsis thaliana ch1-1_ mutant. Acta Physiologia Plantarum 36(7): 1627-1635.
-
Jahan MS, Khanif YM, Sinniah UR (2013) Effects of low water input on rice yield: Fe and Mn bioavailability in soil. Pertanika Journal of Tropical Agricultural Science 36(1): 27-34.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note