Linking Compost Residue to ABAGE in Plants - a Short Note
Compost residue refers to legacy compost present in the soil from what remains after microbial and plant utilization and losses. These losses may be nutrients and other non-nutrients that migrate out of the soil or vaporize. Compost is a brown to dark-brown humic material derived from the biological decomposition of organic matter. Compost contains humic and nonsubstances, which include fulvic acid, humic acid and humin, nutrient elements, and macromolecules such as nucleic acids, carbohydrates, proteins, fats and lipids, and various beneficial microbiome communities. Following soil amendments with compost, some of these substances remain as residues at the end of the growing season and form complexes with the soil organic matter. Abscisic acid glucose ester (ABAGE) is one of the catabolites of ABA formed by conjugation of small molecules such as glucose and catalyzed by the ABA O-glycosyl transferase enzyme. ABAGE is proposed to be the storage or transport forms of ABA. ABAGE is hydrolyzed by BGLU18 to free ABA under stress conditions.
Introduction
Composting of organic waste prevents irresponsible waste disposal that causes poor sanitation and environmental pollution, as these can lead to major human health issues such as outbreaks of cholera, diarrhoea, hepatitis A, typhoid, and dysentery among others [1]. These organic wastes, including manure, food processing, farm and municipal solid wastes are rich sources of plant nutrients and plant growth promoting compounds [2, 3, 4]. As such, compost – a brown to black humic-like substance rich in nutrient elements, and macromolecules such as nucleic acids, carbohydrates, proteins, fats and lipids, and various beneficial microbiome communities – has become an accepted organic soil amendment for the enhancement of soil physical, chemical, and microbiological properties [3, 4]. Typically, compost improves soil structure and ecosystem services such as the addition of organic matter to improve soil structure, water retention capacity, aeration and diversity and distribution of beneficial soil microbiome [3, 5]. Not all the compost applied to soil is utilized by plants due to the slow breakdown and release of nutrients. Therefore, continuous application of compost leaves significant amounts of nutrient and non- nutrient chemical residues that can potentially improve the resilience of plants and may directly or indirectly help slow down global climate change by increasing soil carbon storage capacity [6]. Although compost is used globally and has gained global attention, its enhancement of plant resilience is not clearly elucidated. This entry is a description of tomato (Solanum lycopersicum ‘Scotia’) plant resilience to water- deficit stress as influenced by long-term soil organic matter build-up from compost residue and impact on changes in plant abscisic acid glucose ester (ABAGE) content. ABAGE is a major conjugate and metabolically inactive form of ABA.
Materials and Methods
The study was performed at the Department of Plant, Food, and Environmental Sciences, Faculty of Agriculture, Dalhousie University, Nova Scotia, Bible Hill between February and May 2023. The soils were obtained from a 5-year organic field research plot in Agaard Farms, Brandon, Manitoba, Canada between fall 2015 and winter 2020. The soil type was Orthic Black Chernozem solum on moderate to strong calcareous, loamy morainal till of limestone, granitic and shale origin within the Newdale soil series [7]. The compost was obtained from the City of Brandon municipal solid waste management facility for the study.
Field History
A comprehensive analysis of the compost and the location of soil bio-physicochemical properties was previously published by Abbey, et al. [3, 8, 9]. The compost application rate was approximately 2 t/ha at a bulk density of 650 kg/m3. In brief, the compost was applied every fall to annually assigned plots (AN-soil) and every other year to biennially assigned plots (BI-soils). The control plots (C-soils) did not receive compost throughout the five years of the research. At the end of the fifth year, 20 kg of soil samples were dug randomly with a shovel at a depth of 0 - 30 cm from five locations per plot and were bulked to obtain triplicate samples of 100 kg of soil per treatment. The soil samples were packaged and shipped to the Department of Plant, Food, and Environmental Sciences for cold storage at 4oC until use.
Planting and Water-Deficit Stress Treatment
Fifteen-cm diameter plastic pots were filled with 1 kg of AN-soil, BI-soil and C-soil and each pot was placed in a 20-cm diameter plastic saucer. The soil was watered to field capacity and allowed to stand for 24 hr before transplanting one 5-week-old tomato cv. Scotia seedling into each pot. The corresponding plants were labeled AN-plant, BI-plant and C-plant. The potted-plants were arranged in a completely randomized design with four replications in a Biotronette Mark II Environmental Chamber (Lab-Line Instruments Inc., Melrose Park, ILL, USA) at 24°/20°C day/night temperature cycle and 12/12 hr day/night light cycle. The plants were watered to field capacity every 3 days for 4 weeks before the water-deficit stress treatment. The water deficit lasted for 8 days when all the plants showed clear signs of wilting, especially the C-plants which were visually completely wilted. The potted plants were rearranged weekly to offset unexpected variations in microclimate. The soil moisture contents at the time of cessation of watering were 41%, 35.4% and 30.5% field capacities for the AN-soils, BI-soils and C-soils, respectively; while the respective soil moisture content at the end of the water-deficit stress were 5.1%, 2.9% and 0.7%.
Video-Plant Water Stress Recovery
The wilted plants were re-watered to their respective soil field capacities after 8 days of water-deficit stress and a 10-min video was recorded using a DS126801 digital Canon camera (Canon Inc., Tokyo, Japan) to show the timely recovery of the plants under room temperature conditions of 22oC.
Data
Soil samples were sent to the Prince Edward Island Analytical Laboratory, Charlottetown for organic matter analysis using the combustion method #S/N:20014 as detailed in Laboratory manual #SFL 49P. Soil pH was determined by scooping 10 cm3 of dried, ground soil into a 50 mL vial and reverse osmosis water was added at 1:1 ratio (v/v). The mixture was mixed thoroughly before measuring the pH. All data were subjected to a one-way analysis of variance (ANOVA) using Minitab vers. 21 (Minitab, Inc., State College, Pennsylvania, USA). Tukey’s honestly significant difference post-test was used to separate treatment means at P<0.05.
Results and Discussion
History
Previous work by Abbey, et al. [3] on the same AN- soil, BI-soil and C-soil bio-physicochemical composition indicated high soil nutrients and organic matter content in AN-soil followed by the BI-soil and the least in the C-soil. The high organic matter significantly (P<0.05) increased soil bulk density, porosity, and water-holding capacity. Estimated nitrogen release in the AN-soil was higher than in the BI-soil and the C-soil. Microbiome assessment indicated that Deltaproteobacteria, Bacteroidetes Bacteroidia, and Chloroflexi Anaerolineae were overrepresented in the AN- soil and the BI-soil compared to the C-soil. Also, there was a strong positive correlation (r = 0.89) between AN-soil and 18 microbial metabolic pathways out of 205 that were detected.
Residual Active Organic Matter
The impact of residual compost on soil health can be determined using many indicators. However, one of the best soil health indicators is soil organic matter (SOM) content.
Soil organic matter, measured as total soil carbon, represents the amount of carbon compounds in the soil that are derived from living and dead organisms [10]. High SOM is positively associated with high soil organic carbon (SOC) [11, 12]. SOM can be found at different stages of decomposition in the soil where it directly influences most of the important soil properties and ecosystem services like soil fertility, soil nutrient cycling, soil water storage and infiltration, as well as soil microbial communities and their activities [3, 6, 9]. Therefore, after 5 years of annual compost application to the AN-soil, it is expected that residual SOM will increase significantly (P<0.01) in AN-soil compared to BI-soil and C-soil, as presented in Table 1. This SOM comprised a stable fraction – humus – which is resistant to decomposition and stores carbon; and an active fraction, which is recently formed (1-5 years) and can be decomposed to become readily available to plants. This makes the active SOM an important fraction that consisted of decomposing OM with a positive impact on soil structure and ecosystem services [13].
| Soil treatment | Total OM (%) | Active OM (µg/g) | CEC (Meq/100g) |
|---|---|---|---|
| AN-soil | 8.0a | 764.5a | 47.5a |
| BI-soil | 4.75bc | 572.1b | 38.0b |
| C-soil | 3.85c | 371.5c | 34.2c |
| Significant level at α = 0.05 | ** | *** | * |
Table 1: Residual organic matter (OM) and cation exchange capacity (CEC) of soils previously treated with compost at different fr
1 AN-, BI- and C-soils are soils with residual compost from the previous five years of treatment with annual, biennial and control (no compost) application, respectively. Differences in alphabetical letters within a column represent differences in treatment means at P≤0.05. *, , * denotes P-values at 0.05, 0.01 and 0.001, respectively, from the analysis of variance.
It is therefore expected that soils with high compost residue will have high SOM that will inevitably improve the fluxes and composition of soil gas, modulate soil temperature, cycle soil carbon and nutrients, cycle water, natural decomposition and recycling of organic debris and the beneficial microbiome communities and their functions [14, 15]. Furthermore, land use systems and management practices in addition to climatic and edaphic factors can significantly impact soil carbon dynamics [16, 17]. The trend of the active SOM was AN-soil > BI-soil > C-soil (Table 1), which culminated in a correspondingly higher ability of the AN-soil to hold and exchange nutrients (CEC) although pH did not significantly (P>0.05) vary. According to Solly, et al.
[10], effective CEC and SOC content are strongly correlated when pH > 5.5. The pH of the soils in the present study ranged between 7.4 and 7.6, which can explain the strong association between the active SOM and the CEC values as SOC directly correlates with SOM. The reason is that the surfaces of SOM have negative charges and are sites for cation exchange and as such, the higher the CEC, the higher the soil fertility [6, 18, 19, 20]. Therefore, the high SOM in the AN-soils suggests better SOC preservation and high sorptive capacity for high CEC [21]. On the other hand, anion exchange capacity increases with decreasing pH<5.5 as the charged surfaces of the SOM become more positive due to the protonation of functional groups [22]. This suggests that the higher residual compost in the AN-soils of pH 7 led to higher SOM and CEC such as calcium, potassium, and magnesium compared to the BI-soil with the least in the C-soil. This also made the AN-soil more fertile for higher plant growth and resilience compared to the BI-soil and the C-soil as shown in Figure 1 below.
Plant Growth and Water-Deficit Stress Response
Figure 1 shows the variation in tomato ‘Scotia’ plant growth with and without water-deficit stress in soils with different levels of compost residue. Plant growth markedly varied according to the level of compost residue with the growth trend AN-plants > BI-plants > C-plants. Insufficiency in soil residual nutrients, particularly nitrogen [3, 9], led to a drastic increase in leaf paleness (chlorosis) and reductions in the growth of the BI-plants and C-plants even when the plants were watered regularly to field capacity. Watering cessation for 8 days caused severe wilting in the C-plants followed by the BI-plants and moderate in the AN-plants. This visual observation Figure 1 was proof that an increase in the level of residual soil compost can remarkably increase plant resilience to water-deficit stress.
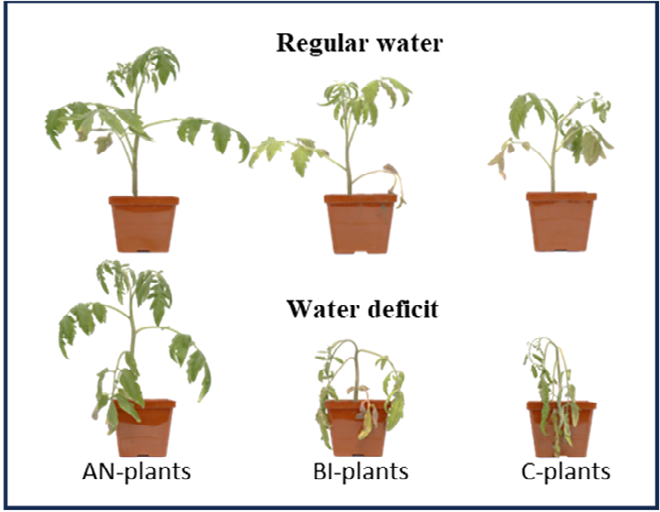
Figure 1: Visual differences in tomato (Solanum lycopersicum ‘Scotia’) plants (n = 9) growth under regular water and water-deficit stress as influenced by soil residual compost. AN-, BI- and C-plants denote plants grown in soils with residual compost effect from the previous five years of treatment with annual, biennial and control (no compost) application, respectively.
Plants are sessile and are therefore exposed to varied environmental stresses – mainly climate and edaphic-related stresses – which warrants prompt response and use of adaptive strategies to enable them to survive and proliferate. Management techniques such as irrigation, fertilization, soil amendments, and pest and disease management also help to improve plant resiliency and rapid response to environmental stresses [23, 24, 25]. As evidenced in Figure 1, an increase in residual soil compost had tremendous benefits to the AN-plants compared to the BI-plants and C-plants. Obviously, the annual application of compost in the long- term seemed to have left residues of its content comprising humic and non-humic substances in the soil. These residues impact soil health including essential plant nutrients, and the growth and development of subsequent plants [25]. Ultimately, the compost residue did not only increase plant growth but improved plant tolerance to water-deficit stress.
Biosynthesis of ABAGE
ABA is an important signaling phytohormone that is involved in plant development and stress physiology [26, 27]. Typically, ABA controls stomatal opening and closure, gene upregulation, and osmolyte synthesis. De novo biosynthesis of ABA in plastids begins with the conversion of 9-cis- epoxycarotenoids to xanthonin catalyzed by 9-neoxanthin cis-epoxicarotenoid dioxygenase (NCED) [27, 28] as shown in Figure 2. The biosynthesized ABA can be inactivated through several routes. Firstly, ABA 8′-hydroxylase (CYP707A) cleaves ABA into 8′-OH-ABA and the products are immediately converted to the primary products, dihydrophaseic acid (DPA) and phaseic acid (PA).
Secondly, ABA can be conjugated by glucose catalyzed by the ABA O-glycosyl transferase enzyme to form an inactive ABAGE, which is then stored in the vacuole or transported through the xylem into potential sinks [29, 30]. Under considerable osmotic stress, ABAGE is cleaved by beta- glucosidase 18 (BGLU18) enzyme into free and active ABA in response to plant stress [31]. This mechanism occurs in almost all plants including tomato and can explain the alterations in ABAGE content of the regularly watered and water-deficit stressed plants as described below.
Residual Compost Alteration of ABAGE Under Water-Deficit
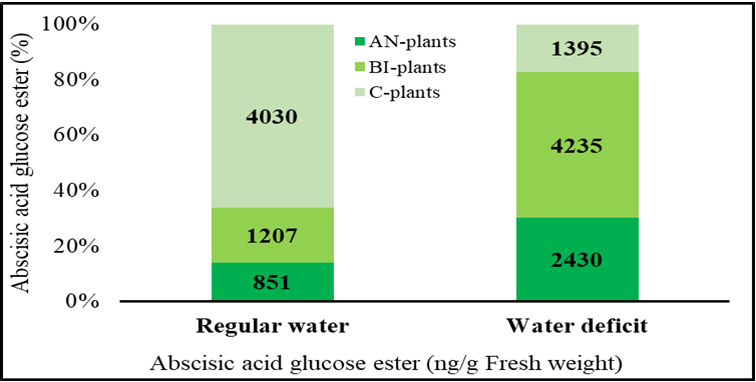
The variations in the levels of residual compost in the soils can be associated with the remarkable differences in the different contents of ABAGE in the plants (Figure 3). When water was not limiting, the ABAGE content in the C-plants was about 3.7-folds and 2.3-folds higher than those of the AN-plants and the BI-plants, respectively. This suggests a higher amount of inactive and stored ABA in plants grown in the nutrient-insufficient soils i.e., the BI-plants and the C-plants. Total ABA from active (i.e., cis-ABA) and inactive ABA
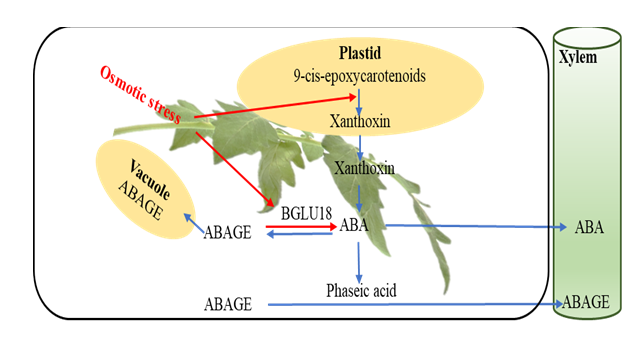
Figure 3: Abscisic acid glucose ester (ABAGE) content of tomato (Solanum lycopersicum ‘Scotia’) plants (n = 9) under regular water and water-deficit stress as influenced by soil residual compost. The bars show the percentage differences in ABAGE, numbers within the bars show the actual value of ABAGE in ng/g fresh weight. AN-, BI- and C-plants denote plants grown in soils with residual compost effect from the previous five years of treatment with annual, biennial and control (no compost) application, respectively.
(i.e., ABAGE, trans-ABA, DPA, PA and neo-PA) were previously reported to be high in abiotically stressed plants [27, 28]. As such, it can be surmised that free and active ABA and its catabolites including ABAGE could be present in high amounts in the C-plants and the BI-plants. Under water- deficit stress conditions, ABAGE is hydrolyzed by BGLU18 to free and active ABA [31]. Therefore, it seems most of the ABAGE in the C-plants were hydrolyzed to ABA leading to low levels of ABAGE compared to the AN-plants and BI-plants in response to water-deficit stress (Figure 3). The increase in ABAGE in the AN-plants and BI-plants under water-deficit stress conditions suggests an increased catabolism of de novo biosynthesized ABA and subsequent conjugation to ABAGE [27, 28, 31]. This means the imposition of environmental stress such as water-deficit stress instigates plant response via an internal network of signaling compounds including ABA to cause regulation of genes, metabolic reprogramming, and crosstalk of phytohormones for plant growth adjustment and ultimately, survival and adaptation.
Video-Plant Water Stress Recovery
A 10-min video of plant recovery from water-deficit stress in the different soil treatments is presented. Plants recover from water stress when the stress is transient. Otherwise, a permanent wilting point is reached where the plant will not recover after re-watering due to complete cell death [32]. Although Figure 1 visually showed the extent of plant wilting in the different soils with the C-plants showing the worse effect, they recovered after re-watering. However, the time and speed of recovery varied with the level of residual compost in the different soils. The fastest recovery time was recorded by the AN-plants, followed by the BI- plants, and a delay in recovery by the C-plants. The variation in the rate of plant recovery from water-deficit stress can therefore be attributed to the level of soil compost residue. The higher the compost residue present in the soil, the faster the recovery time, probably because the plant was more resilient and healthier compared to plants in soils with low to no compost residue.
Conclusions and Prospects
Legacy compost refers to the residue from soil amendments with compost that remains in the soil after long- term frequent application and utilization by plants, microbial decomposition, and losses through various channels such as soil erosion, run-off, leaching, and vaporization. The results showed the positive impact of compost residue on soil health, plant growth, and resilience to water-deficit stress through the enhancement of soil bio-physiochemical properties and ecosystem services. The overall importance of compost on soil organic carbon and cation exchange capacity was amply demonstrated. The results proved that soils without a history of compost amendment do not benefit plants but rather, reduce productivity and increases the vulnerability of plants to not only water-deficit stress but possibly, to other abiotic and biotic stresses as well. Consequently, soil amendment with compost can be adopted by farmers with the prospects of improving or maintaining soil health for sustainable farming, and to increase plant resilience and productivity. This will have added benefits of increasing soil carbon capture and storage capacity and thereby, contribute to mitigation of greenhouse gas emissions and slowing down climate change.
Author Contributions
Conceptualization, L.A.; methodology, L.A.; validation, L.A., R.O., S.C.; formal analysis, L.A., R.O; investigation, L.A., R.O., E.B.N.; resources, L.A.; data curation, L.A., S.C.; writing—original draft preparation, L.A.; writing—review and editing, R.O., S.C., E.B.N; Video, R.O.; supervision, L.A.; project administration, L.A.; funding acquisition, L.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by The Compost Council of Canada, Ontario and Manitoba Conservation and Water Stewardship award.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
-
Shah AV, Srivastava VK, Mohanty SS, Varjani S (2021) Municipal solid waste as a sustainable resource for energy production: State-of-the-art review. J Envir Chem Eng 9(4): 105717.
-
Pergola M, Persiani A, Palese AM, Di Meo V, Pastore V, et al. (2018) Composting: The way for a sustainable agriculture. Applied Soil Ecol 123: 744-750.
-
Abbey L, Yurgel SN, Asunni AO, Ofoe R, Ampofo J, et al. (2022) Changes in soil characteristics, microbial metabolic pathways, TCA cycle metabolites and crop productivity following frequent application of municipal solid waste compost. Plants 11(22): 11223153.
-
Singh YP, Arora S, Mishra VK, Singh AK (2022) Synergizing microbial enriched municipal solid waste compost and mineral gypsum for optimizing rice-wheat productivity in sodic soils. Sust 14(13): 14137809.
-
Waqas M, Hashim S, Humphries UW, Noor R, Naseem A, et al. (2023) Composting processes for agricultural waste management: a comprehensive review. Processes 11(3): 11030731.
-
Fu B, Chen L, Huang H, Qu P, Wei Z (2021) Impacts of crop residues on soil health: a review. Envir Poll Bioavail 33(1): 164-173.
-
MAFRD (2010) Soil Series Description. Manitoba Agriculture, Food and Rural Initiatives pp: 208.
-
Abbey L, Ofoe R, Gunupuru LR, Ijenyo M (2021) Variation in frequency of CQA-tested municipal solid waste compost can alter metabolites in vegetables. Food Res Intl 143: 110225.
-
Abbey L, Ijenyo M, Spence B, Asunni AO, Ofoe R, et al. (2021) Bioaccumulation of chemical elements in vegetables as influenced by application frequency of municipal solid waste compost. Can J Plant Sci 101(6): 967-983.
-
Solly EF, Weber V, Zimmermann S, Walthert L, Hagedorn F, et al. (2020) A critical evaluation of the relationship between the effective cation exchange capacity and soil organic carbon content in Swiss forest soils. Front Glob Change 3: 98.
-
Metting FB, Smith JL, Amthor JS (1999) Science needs and new technology for soil carbon sequestration. In Izaurralde RC, at al. Carbon Sequestration in Soils: Science, Monitoring and Beyond Battelle Press: Columbus, OH, USA, pp: 1-34.
-
Akpa SIC, Odeh IOA, Bishop TFA, Hartemink AE, Amapu IY (2016) Total soil organic carbon and carbon sequestration potential in Nigeria. Geoderma 271: 202- 215.
-
Brady NC, Weil RR (2008) The Nature and Properties of Soils 14th edn. Publisher: Prentice Hall, Upper Saddle River NJ USA, pp: 650.
-
Rezig FAM, Mubarak AR, Ehadi EA (2013) Impact of organic residues and mineral fertilizer application on soil – crop system: II soil attributes. Arch Agron Soil Sci 59(9): 1245-1261.
-
Abbasi MK, Mushtaq A, Tahir MM (2009) Cumulative effects of white clover residues on the changes in soil properties, nutrient uptake, growth and yield of maize crop in the sub-humid hilly region of Azad Jammu and Kashmir, Pakistan. Afr J Biotechnol 8(10): 2184-2194.
-
Post WM, Kwon KC (2000) Soil carbon sequestration and land-use change: processes and potential. Glob Change Biol 6: 317-327.
-
Nunes MR, Veum KS, Parker PA, Moorman TB, wills SA, et al. (2021) The soil health assessment protocol and evaluation applied to soil organic C. Soil Sci Soc Am J 85(4): 1196-1213.
-
Zhao XL, Yuan GY, Wang HY, Lu D, Chen X, et al. (2019) Effects of full straw in corporation on soil fertility and crop yield in rice-wheat rotation for silty clay loamy cropland. Agron 9(3): 133.
-
Tan CJ, Cao X, Yuan SA, Wang W, Qiao B, et al. (2015) Effects of long-term conservation tillage on soil nutrients in sloping fields in regions characterized by water and wind erosion. Sci Rep 5(1): 17592.
-
Ali A, Ghani MI, Haiyan D, Iqbal M, Cai Z, et al. (2020) Garlic substrate induces cucumber growth development and decreases Fusarium wilt through regulation of soil microbial community structure and diversity in replanted disturbed soil. Int/ J Mol. Sci 21(17): 6022.
-
Bailey VL, Bond-Lamberty B, DeAngelis K, Grandy AS, Hawkes CV, et al. (2018) Soil carbon cycling proxies: understanding their critical role in predicting climate change feedbacks. Glob Change Biol 24(3): 895-905.
-
Zhao N, Ju F, Song Q, Qi Z, Ling H (2022) Quantitative assessment of the contribution of soil organic matter functional groups and heteroatoms to PAHs adsorption based on the COSMO-RS model. Sci Total Envir 846: 157415.
-
Mariani L, Ferrante A (2017) Agronomic management for enhancing plant tolerance to abiotic stresses—drought, salinity, hypoxia, and lodging. Horticulturae 3(4): 52.
-
Bulgari R, Franzoni G, Ferrante A. Biostimulants (2019) application in horticultural crops under abiotic stress conditions. Agron 9(6): 306.
-
Giller KE, Hijbeek R, Andersson JA, Sumberg J (2021) Regenerative agriculture: an agronomic perspective. Outlook on Agric 50(1): 13-25.
-
Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR (2010) Abscisic acid: Emergence of a core signaling network. Ann Rev Plant Biol 61: 651-679.
-
De Ollas C, Hernando B, Arbona V, Gómez Cadenas A (2013) Jasmonic acid transient accumulation is needed for abscisic acid increase in citrus roots under drought stress conditions. Physiol Plant 147(3): 296-306.
-
Shehzad I, Xiukang W, Iqra M, Muhammad K, Iqra K, et al. (2022) Phytohormones Trigger Drought Tolerance in Crop Plants: Outlook and Future Perspectives. Front Plant Sci 12: 799318.
-
Dong T, Park Y, Hwang I (2015) Abscisic acid: biosynthesis, inactivation, homoeostasis and signalling. Essays Biochem 58: 29-48.
-
Ondzighi Assoume CA, Chakraborty S, Harris JM (2016) Environmental nitrate stimulates abscisic acid accumulation in Arabidopsis root tips by releasing it from inactive stores. The Plant Cell 28: 729-745.
-
Han Y, Watanabe S, Shimada H, Sakamoto A (2020) Dynamics of the leaf endoplasmic reticulum modulate β-glucosidase-mediated stress-activated ABA production from its glucosyl ester. J Exp Bot 71(6): 2058-2071.
-
Shao HB, Chu Li-Ye, Jaleel CA, Zhao CX (2008) Water- deficit stress-induced anatomical changes in higher plants, Comptes Rendus Biologies 331(3): 215-225.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Building Living Structures – The Use of Space and Time for their Development