The Effect of Seed-Priming by Cobalt-Diglycyrrhizinate on Wheat (Triticum Aestivum L.) Genome
The aim of our study was to monitor the genomic changes of wheat (Triticum aestivum L.) during seed priming with natural elicitor cobalt-diglycyrrhizinate by RAPD markers. The genomic response to wheat seed priming was analysed with respect to the elicitor concentration (four different concentrations), the type of primer used (six random primers) and the type of responsive tissue (roots and leaves of in vitro seedlings). The level of genome response to seed priming was statistically dependent on elicitor concentration (P<0.001). The type of used primer has proven to have an amplification effect (P<0.001) on generating amplicons in the context of elicitor concentration. The genomic response to seed priming was tissue-specific. The priming effect was observed to a greater extent in leaves than in roots (P<0.001). Our results proved the impact of seed priming by natural elicitor, cobalt-diglycyrrhizinate on wheat genome polymorphism.
Introduction
Seed priming has emerged as a promising strategy in modern stress management because it enables the seedling to cope with environmental stresses. It affects biochemical and physiological processes leading to higher stress tolerance ability in the seedlings [1]. In plant defense, priming is defined as a physiological process by which a plant prepares to respond to imminent abiotic stress more quickly or aggressively [2]. The various approaches of seed priming include hydropriming, osmopriming, chemical priming, hormonal priming, biological priming, redox priming, solid matrix priming, etc. Seed-priming with plant hormone and chemical compounds (elicitors) is found to be effective in assuaging the damaging effects of abiotic stress in plants [1, 3]. Elicitors are natural or synthetic compounds, which activate chemical defense in plants by means of production of phenolic compounds and activation of various defense-related enzymes in plants [4]. Treating plants with elicitors in the absence of pathogen can promote plant resistance [5, 6].
Elicited plants or “primed plants” display faster activation of the various cellular defense responses to biotic or abiotic stresses [5, 7]. The effect of priming includes accumulation of phytoalexins, increased gene expression and accumulation of defense-related proteins and response oxidative stress [1, 8]. Priming treatments were found to induce DNA replication and significant changes in proteins synthesis [9, 10, 11, 12]. Quantitative real‐time PCR analysis showed that there was significant variation in gene expression during seed priming (0–48 h). The expression of genes associated with stress appeared at the early imbibition stage (24 h), while those associated with carbohydrate metabolism, protein synthesis and signaling increased at the late imbibition stage (48 h) [13]. The exact molecular mechanism behind priming is not completely known. Molecular markers may be better to control the priming process, but few studies have been conducted to determine which molecular markers are specific to seed priming [13].
Glycyrrhiza glabra L. commonly known as licorice is an important medicinal plant of many therapeutic values [14, 15, 16]. Due to its remarkable biological activities has licorice wide range of applications as a source of bio- based elicitors with promising application in crop protection as potential alternatives to synthetic fungicides [16, 17, 18]. The extract of licorice exhibits the effective antibacterial and antifungal activity due the presence of phytoterapeutical components [16, 19]. It has been defined a significant stimulation and fungi toxic activity of the copper complexes of glycyrrhizic acid [20, 21].
The aim of our study was to monitor the genomic changes of wheat (Triticum aestivum L.) during seed priming with natural elicitor cobalt-diglycyrrhizinate by RAPD markers.
Application of the RAPD markers, which are among the widely used molecular markers in plants, does not need any prior information about the target sequences on the genome and the assay is simple and fast [22]. During RAPD-PCR, amplification takes place when two hybridization sites are similar to each other and in opposite direction. These amplified fragments are totally dependent on the length and size of both the target genome and the primer. Each fragment is derived from a region of the genome that contains two short segments in inverted orientation, on opposite strands that are complementary to the primer and sufficiently close together for the amplification to work [23].
RAPD markers have been successfully used in wheat germplasm evaluation, because of their many advantages. RAPD gained importance due to its simplicity, efficiency and non-requirement of sequence information. RAPD provides virtually limitless set of descriptors to compare individual plants and populations [24]. RAPD markers have been widely used I genotyping studies of wheat [22, 24, 25, 26, 27, 28].
Material and Methods
Plant Material
The grains of Triticum aestivum L. variety Dustlik, originated from Uzbekistan were used in the study. Grains were surfaced sterilized in 0.1% solution of mercuric chloride during 5 minutes, followed by sterilization in 70% ethanol for 5 min and rinsed in sterile distilled water three times. Consequently, grains were immersed in the solution of the plant-based elicitor, cobalt- diglycyrrhizinate [29, 30]. Of various concentrations (1%; 0.1%; 0.01% and 0.001%) during 2 hours. Different concentrations of the agent were prepared using the sterile distilled water. The procedure of seeds material preparation and fungicide stock solutions was carried out in flow laminar cabinet.
Establishment of in Vitro Cultivation
The grains were placed on Murashige &Skoog plant growth medium under aseptic conditions, five grains per tissue culture vessel [31]. In total, twenty grains per tested variant were used. The cultivation has been carried out under controlled conditions (photoperiod 16/8 h day/night; 23°C/20°C and light intensity 50 µmol/m2/s). The germination of grains started after two days of cultivation.
Genomic DNA Isolation
Pooled sample of 10 five-days old seedlings (first rinsed with water) were used to isolate the genomic DNA from roots and leaves separately by the Saghai-Maroof et al. methods [32]. The pool of tissues from ten plants was used for genomic DNA isolation. The quality and concentration of isolated DNA was checked by Nanophotometer P360 (Implen).
PCR-RAPD amplification was performed in 20-µl of total volume reaction mixture containing 20 ng of genomic DNA, 1 × PCR buffer containing 0.8 M Tris-HCl,
0.2 mM dNTP mix (Invitrogen) and 0.4 µM of random primers. In total 6 RAPD primers were used (Tab. 1). The PCR amplification protocol consisted of initial denaturation at 940 C for 2 min; 45 cycles of denaturation at 940C for 1 min, annealing at 360 C for 1 min, extension at 720C for 2 min and final extension for 7 min at 720 C. The PCR-RAPD products were separated on 1.5% agarose gel together with 1Kb DNA ladder (Bio-Rad) running in 1×TBE buffer at a constant power 90 V for 1 hour. The optimization reactions were performed and amplification reactions were done in duplicates.
| Primer | Sequence (5'-3') |
| OPB-01 | GTTTCGCTCC |
| OPB-05 | TGCGCCCTTC |
| OPB-06 | TGCTCTGCCC |
| OPB-09 | TGGGGGACTC |
| OPB-11 | GTAGACCCGT |
| OPB-12 | CCTTGACGCA |
Table 1: Sequences of RAPD primers used in seed- priming study. 4.4.
Table 1: Sequences of RAPD primers used in seed- priming study. 4.4. Data Analysis Agarose gels were visualized in the G-Box Syngene electrophoresis documentation system. For the recording of individual tracks fragments profiles were gels analyzed by the GeneTools software (Syngene). Each fragment is characterized by quantity and volume of its profile in pixels. Profiles are recorded on the basis of set threshold value in which the analysis is carried out. The genomic changes as a response to seed priming with the elicitor were evaluated by comparing the DNA fragment profiles of individual lines of electrophoreograms. The polymorphism could be observed in (a) the amount, (b) spatial distribution and (c) the intensity of amplified fragments represented by the covered area of individual peaks. Data were subjected to analysis of variance (ANOVA, P<0.01) and the means were compared by Tuckey multiple range test using software Statgraphics version 5.0.
Results
The genomic response to wheat seed priming by plant- based elicitor, cobalt-diglycyrrhizinate was analysed with respect to the elicitor concentration (four different concentrations), the type of primer used (six random primers) and the type of responsive tissue (roots and leaves of in vitro seedlings). The number and position of individual RAPD amplicons were evaluated.
The level of genome response to seed priming was statistically dependent on elicitor concentration (P<0.001). Increasing concentration of applied elicitor, positively affected the occurrence of random amplicons (Fig. 1). However, the most of polymorphic RAPD fragments were observed at elicitor concentration of 0.001% which was almost comparable to control.
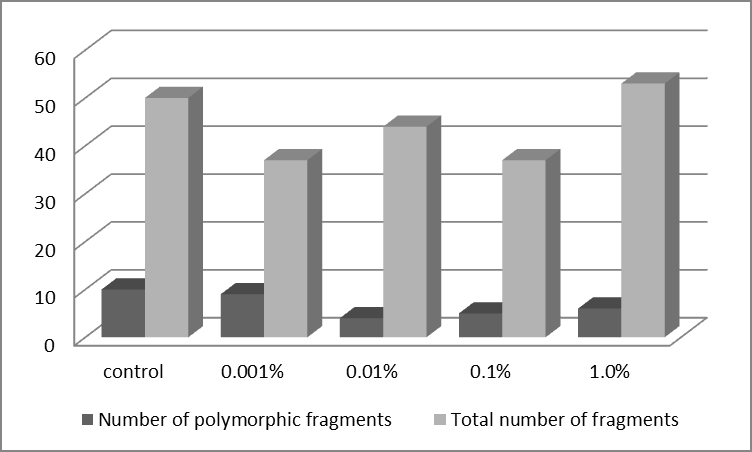
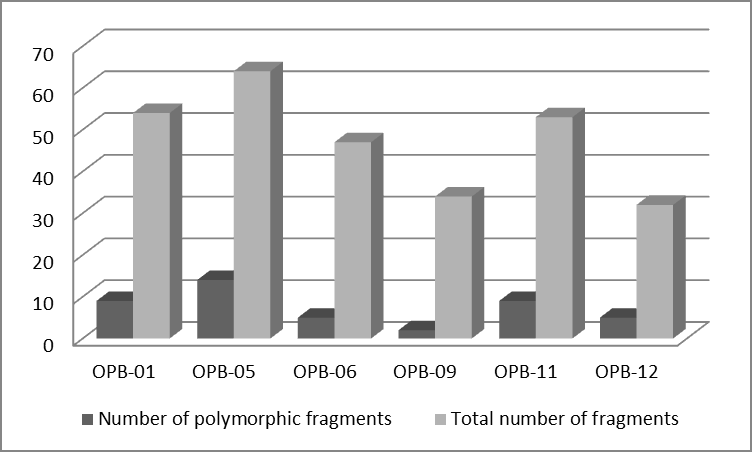
Polymorphic loci are unique in size, resulting in a shift in their localization to gels as compared to other monomorphic fragments. From this point of view is interesting observed polymorphism in polymorphic fragments occurrence at individual concentrations of seed priming agent. Number of polymorphic amplicons increased depending on the increasing concentration (0.01% - 0.1% - 1.0%) of elicitor. However, as was already The type of used primer has proven to have an amplification effect (P<0.001). Total number of amplified fragments as well as the number of polymorphic ones was observed with primer OPB-05 (Fig. 2). All six used decamer primers provided the amplification of repeatable fragments.
mentioned, the most of polymorphic RAPD fragments were observed at the lowest concentration (0.001%).
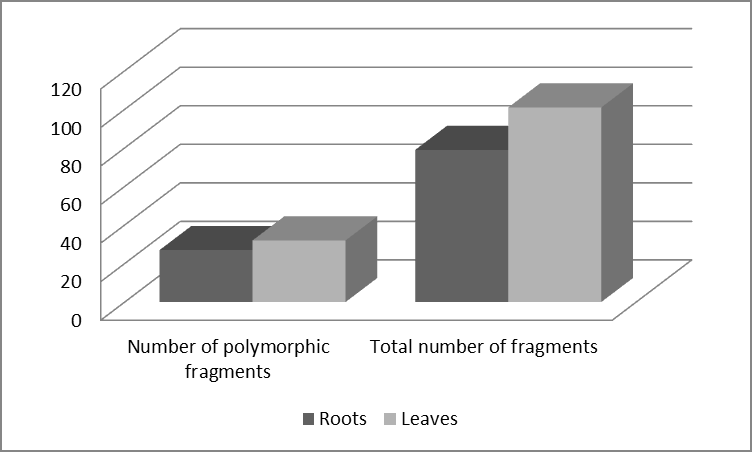
The genomic response to seed priming by plant-based elicitor was tissue-specific. This response was statistically significant (P<0.001). The priming effect was observed to a greater extent in leaves than in roots (Figure 3).
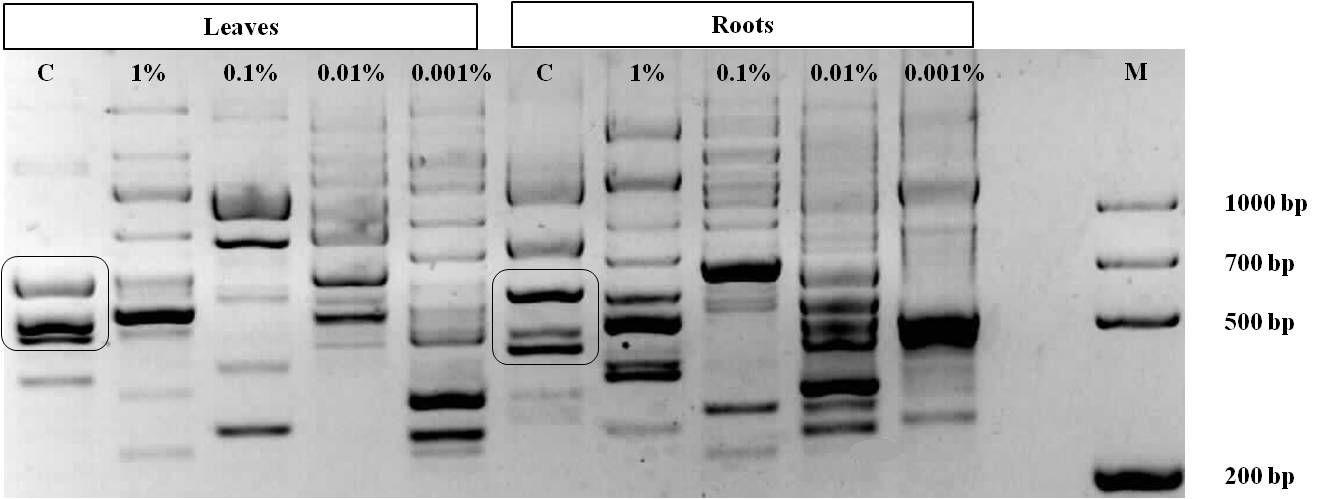
The distinguishable pattern profile of leaves and roots samples is obvious (Fig. 4) as well as the pattern of genomic response to different concentrations of applied elicitor. DNA fingerprinting of RAPD markers correlated not only with the concentration of seed priming solution but also with type of tissue where the effect was detected.
Discussion
Seed priming offers a smart, effective and realistic option for effective plant protection. Seed-priming enables the seedling to cope with environmental stresses. It affects biochemical and physiological processes leading to higher stress tolerance ability in the seedlings [1]. For successful application of seed priming by plant-derived elicitors in agriculture it is necessary to understand their effects on plant genome. Several chemicals were employed to bring about priming in various crops [2].
The results have shown that seed priming with the plant-derived elicitor affected the genome response of wheat. DNA fingerprinting generated by RAPD polymorphism has sensitively reflected to different concentrations of cobalt-diglycyrrhizinate, while this response was tissue-specific, what has been confirmed by several experiments. During the priming treatment was observed the increase of DNA content (4C DNA) in the embryonic root tip cells of pepper. Priming treatments were found to induce DNA replication in the embryo root meristems. Significant difference was observed in DNA content in priming seeds when compared with the unprimed seeds [10].
Randomly amplified polymorphic DNA markers are characterized by high level of observed polymorphism [22, 23]. In our study we confirmed high levels of polymorphism presented between primer binding sites. Depending on the elicitor concentration, the values of polymorphism ranged from 76% (0.001% elicitor concentration) to 91% (0.01% elicitor concentration). Depending of the used decamer primer, these values varied from 83% (primers OPB-01 and OPB-011) to 94% (primer OPB-09). The primer OPB-09 reached 100% polymorphism in evaluation of molecular relationships of 33 wheat accessions [26].
We can state that all six used primers were polymorphic. And finally, depending of the analyzed tissue, the polymorphism reached 66% in roots samples and 68% in leaves samples. Despite the dominant character of RAPD markers, they have been successfully used in wheat germplasm evaluation [22, 24, 25, 26, 27, 28]. Among other things, it is also because RAPD markers provide a wide range of descriptors to compare generated DNA finger printings of individual plants, which has been proven by our study too [24].
Observed significant effect of seed priming with cobalt-diglycyrrhizinate reflected genomic changes of wheat genome which can be detected by RAPD molecular markers. These changes located in primer binding sites create polymorphism [25]. The changes induced by elicitors treatment might be due to differentially expressed plant genes that do not have to relate directly to plant defense mechanism, however regulate plants ontogenesis or nutrient metabolism and consequently are implemented in plant response to pathogen as well as of different abiotic factors [3, 9, 18, 19]. For example, expression analyses of Rice Drought-responsive genes (RD1 and RD2) genes showed upregulated expression in the plants raised from primed seeds under drought stress [1].
There are several parameters which could be tested in order to improve seed priming effect in plant defence, such as contact time with elicitor, the concentration of elicitors which differ with the type and nature of elicitor [1, 4, 33, 34]. As we observed the number of polymorphic amplicons increased depending on the concentration (0.01% - 0.1% - 1.0%) of elicitor. Observed tissue specific reaction to seed priming can be connected with biochemical and physiological processes which take place during this process [1].
Conclusion
The present study was aimed to determine the effect of seed priming with plant-based elicitor on wheat genome. Our results showed that seed priming with cobalt-diglycyrrhizinate significantly affected the genomic response of wheat seedling, while this response was tissue-specific. The cobalt-diglycyrrhizinate has induced polymorphism that can be detected by random polymorphic markers. Therefore, exogenously applied plant-derived elicitors may provide promising alternative tool of environment-friendly plant protection system. However, knowledge of the mechanism of action of these elicitors is a necessary part of their full application in agricultural practice.
Acknowledgment
The project Buiding "AgroBioTech" Research Centre ITMS 26220220180 (50%) and the project APVV-15-0156 (50%).
References
-
Samota MK, Sasi M, Awana M, Yadav OP, Amitha Mithra SV, et al. (2017) Elicitor-Induced Biochemical and Molecular manifestations to improve drought tolerance in rice (Oryza sativa L.) through seed- priming. Frontiers in Plant Science 8: 934.
-
Jisha KC, Vijayakumari K, Puthur JT (2013) Seed priming for abiotic stress tolerance: an overview. Acta Physiologiae Plantarum 35(5): 1381-1396.
-
Lavanya SN, Niranjan-Raj S, Amruthesh KN (2016) Transcript profiling of vital defense proteins upregulated during 3, 5-Dichloroanthranilic acid (DCA) mediated induced resistance against pearl millet downy mildew disease. International Journal of Life Sciences 5(1): 46-54.
-
Thakur M, Sohal BS (2013) Role of elicitors in inducing resistance in plants against pathogen infection: a review. International Scholarly Research Notices Biochemistry 2013: 10.
-
Garrido-Gala J, Frias V, Perdices-Hoyo M, Hernandez- Ros JM, Rodrigez-Olmos A, et al. (2014) Natural elicitors of plant defense response in strawberry. Journal of Berry Research 4(1): 37-45.
-
Oliviera MDM, Varanda CMR, Felix MRF (2016) Induced resistance during the interaction pathogen x plant and the use of resistance inducers. Phytochemistry Letters 15: 152-158.
-
Burketova L, Trda L, Ott PG, Valentova O (2015) Bio- based resistance inducers for sustainable plant protection. Biotechnology Advances 33(6): 994-1004.
-
Wiesel L, Newton AC, Elliott I, Booty D, Gilroy EM, et al. (2014) Molecular effects of resistance elicitors from biological origin and their potential for crop protection. Frontiers in Plant Science 5: 655.
-
Lanterii S, Portis E, Bergervoet HW, Groot SPC (2000) Molecular markers for the priming of pepper seeds (Capsicum annuum L.). Journal of Horticultural Science and Biotechnology 75(5): 607-611.
-
Pradeepkumar GB (2013) Biochemial and Molecular Basis of Seed Priming in Soybean Seed. Thesis, pp: 76.
-
Varier A, Vari AK, Dadlani M (2010) The sub cellular basis of seed priming. Current Science 99(4): 450- 456.
-
Gong F, Wu X, Wang W (2013) Comparative proteomic identification of embryo proteins associated with hydropriming induced rapid- germination of maize seeds. Plant Omics Journal 6(5): 333-339.
-
Cheng J, Wang L, Zeng P, He Y, Zhou R, et al. (2016) Identification of genes involved in rice seed priming in the early imbibition stage. Plant Biology 19(1): 61- 69.
-
Wang D, Pang Y-X, Wang W-Q, Wan C-Y, Hou J-L, et al. (2013) Effect of molybdenum on secondary metabolic process of glycyrrhizic acid in Glycyrrhiza uralensis Fisch. Biochemical Systematics and Ecology 50: 93- 100.
-
Dalme M (2014) Glycyrrhiza glabra (Liquorice)-a potent medical herb. International Journal of Herbal Medicine 2(2): 132-136.
-
Wang L, Yang R, Yuan B, Liu Y, Liu C (2015) The antiviral and antimicrobial activities of licorice, a widely-used Chinese herb. Acta Pharmaceutica Sinica B 5(4): 310-315.
-
Badkhane Y, Yadav AS, Bajaj A, Sharma AK, Raghuwanshi DK (2014) Glycyrrhiza glabra L. a miracle medicinal herb. Indo American Journal of Pharm Research 4(12): 5808-5816.
-
Ablakulova NA, Kushiev Kh (2015) Influence of rowth regulators on contamination of winter wheat. The American Journal of Biological and Chemical Research 4: 91-99.
-
Chandra JH, Gunasekaran H (2017) Screening of phytochemical, antimicrobial and antioxidant activity of Glycyrrhiz glabra root extract. Journal of Environmental Biology 138: 161-165.
-
Ablakulova NA, Ismailova KM, Kushiev Kh (2015) Regulation of growth of fungal diseases of winter wheat with use of physiologically active compounds. Bulletin of the Agrarian Science of Uzbekistan 4: 62.
-
Ismailova K, Ablakulova N, Kushiev Kh (2016) Fungitoxic and growth-promoting propoties of the complexs copper component and glycyrrhizin acid. European Science Review (5-6): 3-6.
-
Shende S, Walunj S (2013) Molecular characterization of wheat (Triticum species) using random amplified polymorphic DNA marker. Recent Research in Science and Technology 5(2): 29-32.
-
Khan S, Mirza KJ, Tayaab Md, Abdin MZ (2009) RAPD profile for authentication of medicinal plant Glycyrrhiza glabra Linn. Medicinal and Aromatic Plant Science and Biotechnology 3(1): 48-51.
-
Cifci AE, Yagdi K (2012) Study of genetic diversity in wheat (Triticum aestivum) varieties using random amplified polymorphic DNA (RAPD) analysis. Turkish Journal of Field Crops 17(1): 91-95.
-
Ebadi M, Eghbali M (2017) The comparison of ISSR and RAPD markers with different species Triticum. Bioscience Biotechnology Research Communications 1: 288-292.
-
Sadigova S, Sadigov H, Eshghi R, Salayeva S, Ojaghi J (2014) Application of RAPD and ISSR markers to analyses molecular relationships in Azerbaijan wheat accessions (Triticum aestivum L.). Bulgarian Journal of Agricultural Science 20(1): 87-95.
-
Bhutta WM, Akhtar J, Ibrahim M, Shahzad A (2006) Genetic variation between Pakistani wheat (Triticum aestivum L.) genotypes as revealed by Random Amplified Polymorphic DNA (RAPD) markers. South African Journal of Botany 72(2): 280-283.
-
Deshmukh R, Tomar NS, Tripathi N, Tiwari S (2012) Identification of RAPD and ISSR markers for drought tolerance in wheat (Triticum aestivum L.). Physiology and Molecular Biology of Plants 18(1): 101-104.
-
Dalimov DM, Isaev YT, Askarov IR (1999) Synthesis of some biogenic metal diglycyrrhizinate derivatives and their complex compounds with ammonia and ethylendiamine. Uzbekskii khimicheskii zhurnal, pp: 24-25.
-
Isaev YI, dalimov DN, Askrov IR (1999) Synthesis of diglycyrrhizinates of some microelements. Special edition, Chem prirod sed, pp: 71-72.
-
Murashige T, Skoog F (1962) A revised medium for rapid growth and bio assays with tabacco tissue cultures. Physiologia Plantarum 15(3): 473-497.
-
Saghai-Maroof MA, Soliman KM, Jorgensen RA, Allard RW (1984) Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location and population dynamics. Proceedings of the National Academic of Sciences 81(24): 8014-8018.
-
Baldi A, Srivastava AK, Bisaria VS (2009) Fungal elicitors for enhanced production of secondary metabolites in plant cell suspension cultures. Symbiotic Fungi 18: 373-380.
-
Govindaraju S, Indra Arulselvi P (2016) Effect of cytokinin combined elicitors (l-phenylalanine, salicylic acid and chitosan) on in vitro propagation, secondary metabolites and molecular characterization of medicinal herb–Coleus aromaticus Benth (L). Journal of the Saudi Society of Agricultural Sciences.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note