Growth of the Broad Bean (Vicia faba) Under the Edaphoclimatic Conditions of Florianópolis, Santa Catarina, Brazil
The cultivation of the Vicia faba, commonly referred to as broad bean, is still not widely spread in Brazil, mainly due to the lack of scientific information on the growth and development of the species. In this context, quantitative research on growth is necessary to better understand the various aspects related to development rates and carbohydrate accumulation in plant organs. Thus, the objective of this work was to analyze the growth of the Italian broad bean during the winter-spring period in Florianópolis, Santa Catarina. In the experiment, a randomized block design with plots subdivided over time was adopted, evaluating leaf area (LA), total dry mass (TDM), absolute growth rate (AGR), relative growth rate (RGR), and net assimilation rate (NAR) at 21, 35, 49, 63, 77, 91, 105, 119, 133, and 147 days after emergence (DAE). The distribution of assimilates in the different plant organs showed sigmoid patterns. A greater accumulation of LA and TDM was observed at 99 and 133 DAE, respectively. Regarding AGR, the maximum values were recorded at 119 and 133 DAE. The rates of AGR, NAR, and RGR varied throughout the cycle, without showing a defined quantitative performance pattern, possibly due to climatic fluctuations and the intrinsic growth rates of the species.
Introduction
The cultivation of broad beans (Vicia faba), also known as fava beans or Italian broad beans in Brazil, is still in its nascent phase, largely confined to small rural estates without commercial objectives. However, on a global scale, the cultivation of the broad bean is significant, covering approximately 24 million hectares and yielding about 446 million tons of grains [1, 2]. In the United States, Canada, and various European countries, this species has become increasingly popular due to its notable nutritional characteristics, which are superior to those of other legumes. These include a high content of proteins, carbohydrates, B-complex vitamins, and essential minerals such as iron, zinc, magnesium, potassium, and phosphorus [3]. Furthermore, the broad bean is recognized for its medicinal uses, notably producing significant amounts of L-Dopa in its parts, a key precursor to dopamine that is used in treating Parkinson’s disease and hormonal imbalances [3].
The species shows good development in mild climates, being frost tolerant and exhibiting optimal growth under conditions of medium to high water availability [4]. Southern Brazil emerges as a conducive environment for its cultivation, although high temperatures and low humidity, especially during the flowering phase, can harm production, leading to floral abortion and reduction in pod production. The cultivation of the broad bean, whether in predominantly cereal areas or in consortium with vegetables, can improve the chemical and microbiological characteristics of the soil and favor the integrated management of pests and diseases [5]. Moreover, the preceding cultivation of the broad bean can enrich the soil with 30 to 120 kg of nitrogen per hectare, with the potential for free atmospheric nitrogen fixation of up to 300 kg ha-1 [4, 6].
However, scientific literature is still scarce regarding the growth and development of the broad bean, highlighting the need for studies that elucidate aspects related to its growth. Quantitative growth research is fundamental for an in-depth understanding of the development rates and carbohydrate accumulation in the plant organs [7].
Principles and practices of plant growth analysis aim to describe and interpret the quantitative performance of species under natural or controlled conditions. This holistic and integrative approach facilitates the interpretation of the form and functionality of the plant, having been developed in the early 20th century by British researchers and established as the standard method for estimating the biological productivity of plant communities [8, 9]. Monitoring plant growth dynamics through mathematical formulas allows for the quantification of plant production and the evaluation of the contribution of different organs to the final growth of the plants, without the need for laboratories or sophisticated equipment, using only the dry matter mass of the plant and the dimension of the photosynthetic apparatus [10, 11]. Thus, quantitative growth analysis proves to be a precise and accessible tool for evaluating plant development and the influence of different physiological processes under various agrotechnological conditions [9].
Plant growth indices quantify the plants’ ability to synthesize and allocate carbohydrates in their organs, essential for the performance of net assimilation over time [12]. Therefore, studies on the quantitative growth of the broad bean under specific conditions can provide valuable insights into the species, contributing to effective management aimed at maximizing productivity according to its edaphoclimatic requirements.
In this context, the present work aims to analyze the growth of the broad bean (Vicia faba) cultivated during the winter-spring in Florianópolis-SC, seeking to contribute to the knowledge gap regarding its development under specific edaphoclimatic conditions.
Materials and Methods
The study was conducted at the Ressacada Experimental Farm, belonging to the Federal University of Santa Catarina (UFSC), located in Florianópolis, SC, Brazil, at coordinates 27º41’06.28” S and 48º32’38.81” W. The soil in this area is classified as a Hydromorphic Quartzipsamment [13]. Based on Köppen’s climate classification, the region features a constantly humid subtropical climate, with an average annual temperature of 20ºC, absence of a dry season, and hot summers. Annual rainfall ranges from 1270 to 1600 mm, with an average relative humidity around 82% and total annual sunshine between 2021 and 2166 hours [14]. To prepare the field for experimentation, plowing was followed by the use of a rototiller to form beds of 1.20 m width and 0.30 m height. Soil amendments and fertilization were carried out as per the recommendations derived from soil analysis, tailored to broad bean cultivation needs [15]. Following liming and fertilization, the soil was reworked, and beds were mulched with a 3 cm layer of branch chips to conserve moisture and suppress weeds.
Seeds of the experimental genotype UFSC-VF-01, selected from UFSC’s vegetable germplasm bank for its unique characteristics, and were sown on May 26, 2022. The layout was designed with a spacing of 0.45 m between rows and 0.20 m between seeds within rows. A randomized block design with subdivided plots was utilized, comprising four replications fixed over time and treatments allocated at different time points. Each plot measured 5.0 m by 1.20 m.
Irrigation was managed through a micro-drip system, calibrated to meet the water requirements of the crop. No chemical treatments were applied for pest and disease management due to the absence of significant economic threats. Weed control was achieved through manual removal.
Temperature data, including daily minimum (Tmin) and maximum (Tmax), were recorded by the meteorological station at UFSC’s agricultural climatology laboratory, located on the experimental farm. These data were used to calculate the daily average air temperature (Tavg).
Primary growth data collection commenced on June 22, 2022 (21 days after emergence [DAE]), with subsequent samplings every fourteen days up to 147 DAE. Leaf area was determined using a transparent clipboard marked with 0.25 cm² squares, selecting three leaves from each of four randomly chosen plants per bed. The dry mass of roots, stems, leaves, and fruits was obtained after drying the material in an oven at 65ºC to a constant weight and weighed on a precision scale accurate to 0.001 g. Total dry mass (TDM) was calculated by summing the dry weights of the plant parts.
Growth rates including absolute growth rate (AGR), relative growth rate (RGR), and net assimilation rate (NAR) were calculated following methodologies outlined in prior research [12, 16]. Data underwent normality and homogeneity of variance tests (Lilliefors and Bartlett, respectively) before being analyzed through ANOVA. Significant results led to polynomial regression analysis, performed using ASSISTAT version 7.7.
Results and Discussions
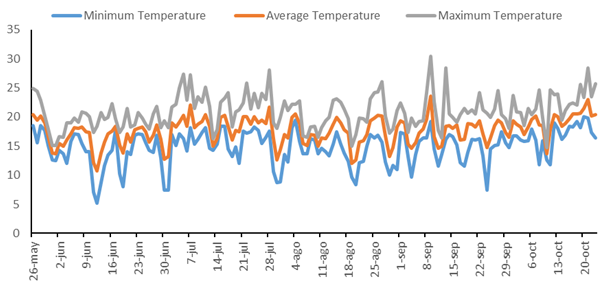
Throughout the experimental period, the minimum temperature was consistently observed to range from 5.0°C (June 8, 2022) to 19.0°C (October 22, 2022), and the maximum from 17.0°C (June 1, 2022) to 32.0°C (September 10, 2022) (Figure 1). Atypical temperatures for this period, such as lows below 10.0°C in September and highs exceeding 28.0°C in July, were recorded, showing significant variations that notably impacted the broad bean’s development. These thermal amplitudes, with at least 10°C variations between the monthly minimum and maximum temperatures, underscore the climate›s influence on plant growth from emergence.
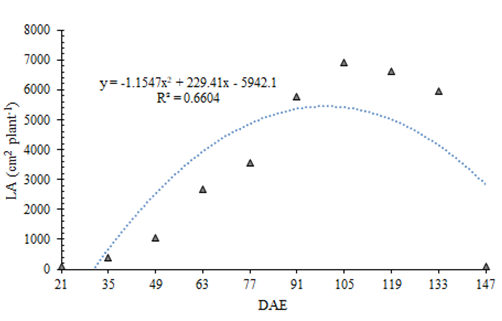
There were significant differences between collection dates for all studied parameters (p < 0.05). The leaf area (LA) of the broad bean exhibited a sigmoid behavior, with a significant increase between 77 and 105 DAE, reaching the maximum estimated LA value at 99 DAE, according to the quadratic adjustment of the regression. Subsequently, the trend was stabilization until 133 DAE, followed by a sharp decline due to senescence, which halted the emission of new leaves (Figure 2). This pattern was also observed in other species, such as cowpea (Vigna unguiculata) Bastos EA, et al. [17], Linhares CM de S, et al. [18] and common bean (Phaseolus vulgaris) [19].
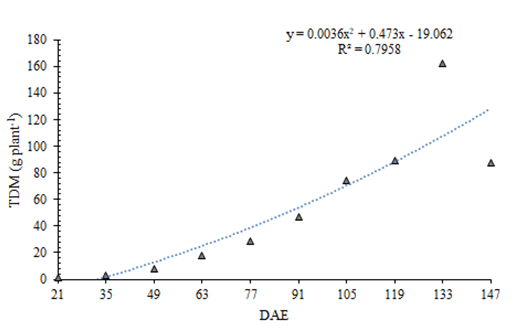
Similar to LA, the TDM of the broad bean exhibited sigmoid behavior. The greatest TDM accumulation occurred between 119 (89.54 g per plant) and 133 (161.89 g per plant) DAE, followed by a decrease from 133 (161.89 g per plant) to 147 (88.00 g per plant) DAE (Figure 3). This sigmoid pattern for TDM has been similarly observed in cowpea (Vigna unguiculata) Moraes J, et al. [20] and jack bean (Canavalia ensiformis) Bortoluzzi MP, et al. [21].
The production of dry mass in the broad bean was slow until 49 DAE, characterized by a high number of expanded leaves and short internodes (Figure 3). From that point, the plant began to branch profusely, and the stems started to contribute more significantly to dry mass production [22]. The maximum accumulation of TDM was reached at 133 DAE (Figure 3), followed by a loss of dry mass due to the senescence of plant organs.
The allocation of assimilates among the different organs of the broad bean followed a clear trend throughout the cycle. With the onset of the reproductive phase, especially marked by the start of fruiting at 91 DAE, there was a more intense direction of photoassimilates to the fruits, which persisted until the end of the cycle. From germination until 49 DAE, the roots exhibited steady growth. However, after this period, a decrease in the dry mass of the roots was noted, coinciding with a significant increase in the dry mass of the stems, which The fruiting phase began at 91 DAE, and the dry mass of the fruits significantly increased until 133 DAE, remaining stable until 147 DAE. Concurrently, a reduction in the dry mass of other plant parts was observed (Figure 4). From the beginning of the formation of the first fruits, there was a more pronounced direction of assimilates from leaves, stems, and roots to the fruits, as these organs began to act as preferential sinks [12, 23].
continued until 91 DAE.
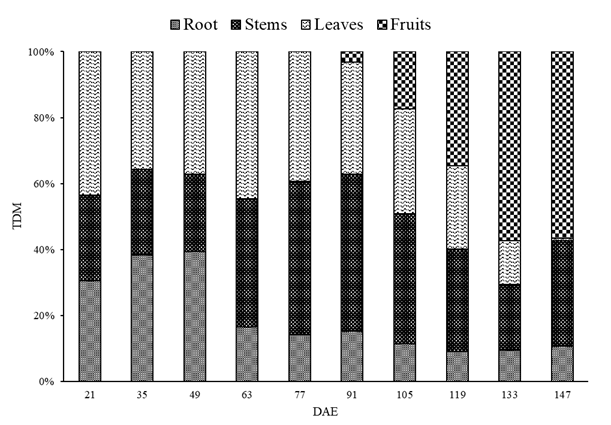
While leaves provide carbon from the photosynthetic process, roots supply water and minerals to the other organs. At the same time, as dry mass accumulates in the aerial part, new roots are also emitted that absorb essential elements promoting growth, flowering, and fruiting [24, 25]. Thus, while the root system is an important source, it requires the direction of assimilates from the leaves to enable the emission and growth of roots.
Leaves acted as source-sink, being the primary producers of photoassimilates, and also accumulated the majority of TDM, which was then redistributed to organs unable to ‘self-sustain,’ such as fruits. Conversely, from the beginning until 91 DAE, stems acted as sinks, showing continuous accumulation of dry mass, followed by a slight drop (105, 119, and 133 DAE), then increasing again at the last data collection (Figure 4). The effects of the source- sink relationship on photosynthesis are regulated by various mechanisms, which directly impact productivity and are influenced by the morphological and physiological characteristics of the source (photosynthesizing organs) and sink (photosynthesized organs, mainly carbohydrates) [26].
The highest accumulation of leaf dry mass occurred at 63 DAE (Figure 4), which continued to have an increase in LA, then becoming a source for other plant organs (with the maximum point estimated at 99 DAE) (Figure 2). The distribution of dry matter among the different plant organs is the final result of a set of metabolic and transport processes, which are the flow of assimilates through the source-sink mechanism [27]. Alongside these processes, plants receive external stimuli, mainly coordinated by thermal energy, directly influencing the sum of degree days that govern growth, development, and phenological stage changes [28]. As phenological stages change, meristematic zones, previously functioning for biomass production, become responsible for the reproductive phase, i.e., fruit development utilizes reserves accumulated mainly in the leaves [29]. This process began in the broad bean plants from 91 DAE (Figure 4), with leaves then experiencing the greatest decrease in dry mass, followed by stems and roots.
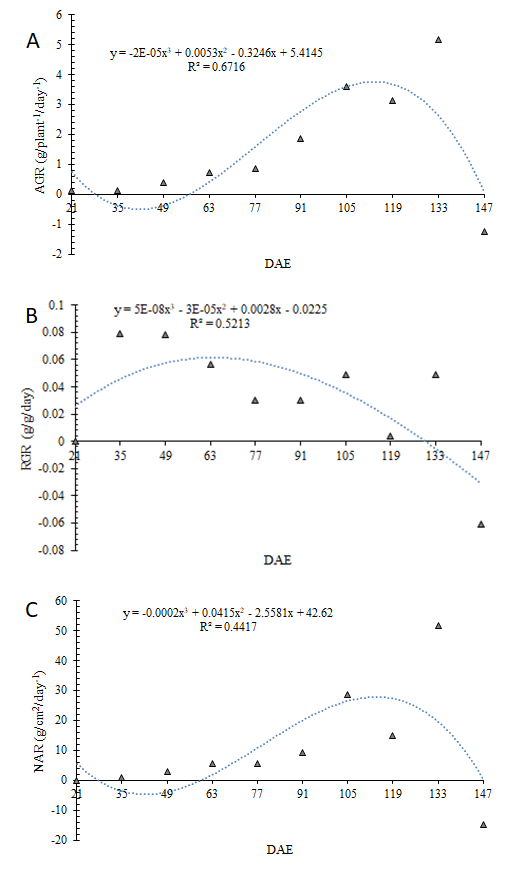
Regarding AGR, NAR, and RGR, variations occurred throughout the cycle without a pattern of quantitative performance, with adjustments to the cubic regression model (Figure 5). For AGR, the maximum rate was between 119 (3.14 g/plant/day) and 133 DAE (5.16 g/plant/day) (Figure 5A), with a declining trend thereafter, including a negative value at 147 DAE (-1.23 g/plant/day), possibly due to the translocation of photoassimilates. For RGR, the maximum value occurred at 35 DAE (0.07 g/g/day), which remained until 49 DAE, presenting variations during growth and a negative value at the end of the cycle (-0.06 g/ plant/day) (Figure 5B). Finally, for NAR, broad bean plants showed an increasing trend in assimilation until reaching the maximum at 133 DAE (51.61 g/cm2/day) and the lowest value observed at 147 DAE (-14.67 g/plant/day) (Figure 5C).
Variations in AGR, NAR, and RGR throughout the cycle (Figure 5) are possibly due to climatic fluctuations (Figure 1) and the inherent growth rates of the species itself. The behavior of NAR and RGR in this study resembles the findings of research conducted on cowpea by Linhares CM de S, et al. [18], showing a tendency to decline during the crop cycle. With the increase in dry mass accumulated by the plants, there is an increased need for photoassimilates to maintain the already formed structures Zeist AR, et al. [12]. Thus, the amount of photoassimilates available for growth tends to be smaller, and consequently, RGR decreases over time [30].
Constant growth in NAR was observed during the vegetative period of the crop. The evolution of this physiological parameter throughout the phenological stages suggests a progressive decrease, followed by a relative constancy of net assimilation in the reproductive phase and a decrease at the end of the cycle [31]. The observation of negative NAR values at the end of the reproductive period results from the reduction in biomass accumulation by the plant [31]. In general, the overall growth of the plant, in terms of volume, weight, linear dimensions, and structural units, is due to what is stored and produced in terms of structural material, presenting variations throughout the cycle due to the phenological characteristics of the species.
Conclusions
In this work, we analyzed the development of the broad bean (Vicia faba) under the edaphoclimatic conditions of Florianopolis, Santa Catarina, throughout the winter-spring period. The investigation fills a gap in the literature, given the limited knowledge on the cultivation of this legume in the mentioned region. A sigmoid growth pattern was observed, with moments of significant biomass accumulation beginning approximately 91 days after plant emergence. This study is distinguished by the observation of peaks in leaf area and total dry mass, recorded at 105 and 133 days, respectively.
The obtained results emphasize the adaptability of the broad bean to local conditions, pointing to the viability of its sustainable cultivation in Florianopolis and areas with similar climatic conditions. The variability in growth rates, influenced by climatic fluctuations and intrinsic species characteristics, reinforces the need for an understanding of resource allocation by the plant, particularly during the reproductive period, for the optimization of species management practices.
This study contributes to the knowledge of Vicia faba growth under specific conditions, providing valuable information for the improvement of agronomic practices and direction of future research. Moreover, it opens perspectives for future investigations on varietal selection, cultivation methods, and integration of the broad bean into crop rotation systems, aiming at sustainability and agricultural productivity.
It is hoped that the findings of this study will motivate subsequent research that contributes to the development of Vicia faba cultivation.
Conflicts of Interest
The authors declare no financial or other conflicts of interest.
Acknowledgements
Federal University of Santa Catarina
References
-
Maalouf F, Hu J, O’Sullivan DM, Zong X, Hamwieh A, et al. (2019) Breeding and genomics status in faba bean (Vicia faba). Plant Breeding 138(4): 465-473.
-
FAOSTAT (2024) Food and Agricultural Organization of the United Nations.
-
Etemadi F, Hashemi M, Barker AV, Zandvakili OR, Liu X (2019) Agronomia, Valor Nutricional e Aplicação Medicinal do Feijão Faba (Vicia faba L.). Horticultural Plant Journal 5(4): 170-182.
-
Singh A, Bharati R, Manibhushan N, Pedapati A (2013) An assessment of faba bean (Vicia faba L.) current status and future prospect. African journal of agricultural research 8(50): 6634-6641.
-
Köpke U, Nemecek T (2010) Ecological services of faba bean. Field Crops Research 115(3): 217-233.
-
Jensen ES, Peoples MB, Hauggaard-Nielsen H (2010) Faba bean in cropping systems. Field Crops Research 115(3): 203-216.
-
Barbero LM, Prado TF, Basso KC, Lima LA, Motta KM, et al. (2013) Análise de Crescimento em Plantas Forrageiras Aplicada ao Manejo de Pastagens. Veterinária Notícias 19(2).
-
Blackman VH (1919) The Compound Interest Law and Plant Growth. Annals of Botany 3(3): 353-360.
-
Peixoto C, Cruz T, Peixoto M (2011) Análise Quantitativa do Crescimento de Plantas: Conceitos e Prática. Enciclopédia Biosfera 7(13).
-
Júnior JPF, Silva L, Santos S, Costa R, Silva J (2015) Crescimento de Bauhinia variegata Linn. SOB Intervalo de Irrigação e Condicionador de Solo, Fortaleza, Ceará, Brazil, pp: 2640.
-
De Souza GS, Lima JC, Dos Santos AR, De Oliveira UC, Bezerra MN (2017) Produção de Fitomassa de Sálvia Officinalis L. Cultivada Sob Malhas Coloridas e Doses de Esterco Avícola. Revista Verde de Agroecologia e Desenvolvimento Sustentável 12(2):182-186.
-
Zeist AR, Faria MV, Resende JTV, Gabriel A, Nonato JJ, et al. (2019) Biomass Association in Specimens and Interspecific Hybrids of Tomatoes. Acta Sci Agron 42: e42806.
-
Dos Santos HG (2018) Sistema Brasileiro de Classificação de Solos. In: 5th (Edn.), DF: Embrapa, pp: 356.
-
Aparecido LE de O, Rolim G de S, Richetti J, Souza PS de, Johann JA (2016) Köppen, Thornthwaite and Camargo climate classifications for climatic zoning in the State of Paraná, Brazil. Ciênc agrotec 40(4): 405-417.
-
Trani P, Passos F, Pereira J, Semis J (2015) Calagem e Adubação do Feijão-Vagem, Feijão-Fava (ou Fava- Italiana), Feijão-de-Lima e Ervilha Torta (ou Ervilha-de- Vagem).
-
Ghamari H, Ahmadvand G (2013) Growth Analysis of Dry Bean (Phaseolus vulgaris L.) in Different Weed Interference Situations. Not Sci Biol 5(3): 394-399.
-
Bastos EA, Ramos HMM, Junior ASA, Nascimiento FN, Cardoso MJ (2012) Parâmetros fisiológicos e produtividade de grãos verdes do feijão-caupi sob déficit hídrico. Water Resources and Irrigation Management 1(1): 31-37.
-
Linhares CM de S, Freitas FCL de, Silva K de S, Lima MFP de, Dombroski JLD (2014) Cowpean Growth under the Effect of the Herbicides Fomesafen and Bentazon+Imazamox. Revista Caatinga 27(1): 41-49.
-
Santana AV de, Heinemann A, Stone LF, Nascente AS (2016) Reflectance index in estimating leaf area and leaf biomass in common bean. Colloquium Agrariae 12(1): 7-19.
-
Moraes J, Souza P, Faria R, Martorano L, Lima M (2016) Análise de Sensibilidade e Calibração do Modelo Cropgro- Cowpea para Simulação do Crescimento do Feijão Caupi (Vigna unguiculata L.) Walp.) No Nordeste Paraense. In Florianópolis-SC Anais.
-
Bortoluzzi MP, Heldwein AB, Leonardi M, Silva JR da, Hinnah FD, et al. (2019) Crescimento, Acúmulo de Fitomassa e Produtividade do “Feijão-de-Porco” em Diferentes Datas de Semeadura. Amazonian Journal of Agricultural and Environmental Sciences.
-
Woo HR, Masclaux-Daubresse C, Lim PO (2018) Plant Senescence: How Plants Know When and How to Die. Journal of Experimental Botany 69(4): 715-718.
-
Lopes W de A, Negreiros MZ de, Dombroski JLD, Rodrigues GS de O, Soares AM, et al. (2011) Análise do Crescimento de Tomate “SM-16” Cultivado Sob Diferentes Coberturas de Solo. Hortic Bras 29: 554-561.
-
Xu X, Yang X, Yang Y (2011) Effect of HCO3- on root growth and nutrient absorption of different rice genotypes. Ying Yong Sheng Tai Xue Bao 12(4): 557-560.
-
Martinazzo EG, Perboni AT, Posso DA, Aumonde TZ, Bacarin MA (2015) Growth analysis partitioning of assimilate in tomato plants cv. Micro-Tom submitted to nitrogen and pyraclostrobin. Sem Ci Agr 36(5): 3001- 3012.
-
Lins HA, Queiroga RCF de, Pereira A de M, Silva GD da, Albuquerque JRT de (2013) Produtividade e Qualidade de Frutos de Melancia em Função de Alterações na Relação Fonte-Dreno. Revista Verde de Agroecologia e Desenvolvimento Sustentável 8(3): 143-149.
-
Silva ABD (2015) Phytomass, yield and quality of melon fruits subjected to changes in the source: drain relationship. Course Completion Work (Bacharelado em Agronomia)-Centro de Ciências e Tecnologia Agroalimentar, Universidade Federal de Campina Grande, Pombal, Paraíba, Brazil.
-
Bardhan K (2016) Crop Regulation: Concepts and Its Commercial Uses. In: Patel NL, Chawla SL, et al. (Eds.), Commercial Horticulture, New India Publishing House, New Delhi, India, pp: 257-275.
-
Zeist AR, Resende JT de, Lima RB, Gabriel A, Henschel JM, et al. (2019) Phenology and Agronomic Components of First and Second-Cycle Strawberry. Hortic Bras 37(1): 29-34.
-
Benincasa MMP (2003) Análise de Crescimento de Plantas (noções básicas). FUNEP, Jaboticabal, pp: 42.
-
Urchei MA, Rodrigues JD, Stone LF (2000) Análise de Crescimento de Duas Cultivares de Feijoeiro Sob Irrigação, em Plantio Direto e Preparo Convencional. Pesq agropec bras 35(3): 497-506.
-
Hunt R, Causton DR, Shipley B, Askew AP (2002) A Modern Tool for Classical Plant Growth Analysis. Ann Bot 90(4): 485-488.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note