Productivity and Quality of Horticultural Crop Capsicum Through Co-Inoculation of Novel Microbial Consortium Plant Growth Promoting Rhizobacteria, Glycoprotein Producing AM Fungi and Chemical Fertilizer under Low-Cost Protected Cultivation
Capsicum (Capsicum annum L.) is a common vegetable crop with a wide range of culinary utilization worldwide. In horticultural practices, inoculating the planting medium with a beneficial microbial consortium is a novel approach to cultivating highquality, healthy plants with abundant nutrition. In the following study, inoculation of a selected microbial consortium consisting of PGPR (Azotobacter, Pseudomonas, Fraturia, Azospirillum, and Bacillus spp.) and glycoprotein producing AM fungi (Aculospora logula-15%, Glomus fasciculatum-20%, Glomus intraradices-40%, Gigaspora margarita-15%, and Scutellospora heterogama-10%) was inoculated to the planting medium in beds to raise capsicum plants in a low cost protected cultivation. Plant height, stem girth, fruit weight, fruit diameter, number of fruits per plant, and weight of fruit per plant parameters to be considered in the study. Mycorrhizal root colonization, macro, and micronutrient uptake, and quality yield, were increased the most with multiple microbial inoculations. Treatments revealed that plants inoculated with the multiple microbial consortia increased substantially faster than plants treated with chemical fertilizer (100 % RDF) and control. Maximum yield (742.5 q ha-1) was recorded in treatment Absolute’s consortium PGPR+ AM Fungi along with maximum values of shelf life (7.10 DAS), TSS (Brix0 5.63), the number of fruit/plant (8.20), fruit length (8.5cm), fruit diameter (7. 81cm) and fruit weight (224g) as compared with control and other biological treatments. The best treatment with respect to projected yield was Absolute consortium PGPR + AM fungi followed by control. Based on the various growth and microbiological parameters studied, it was concluded that inoculation with the multiple microbial consortia (Absolute consortium PGPR+AM fungi) was beneficial for raising healthy, vigorously growing capsicum plants under low-cost protected conditions.
Introduction
Capsicum (Capsicum annuum L.), also known as a bell or sweet pepper, is one of the most widely grown vegetable crops in the world. Because of its richness in vitamins (A, C, E, and K1) and antioxidants, it is commonly regarded as a nutritious food for a healthy human (HH). Its color, scent, flavor, and crisp texture are important factors in its widespread use in a variety of cuisines around the world [1]. In India, it is one of the most significant cash crops. India is the world’s greatest producer of chilli, with an annual output of 1.1 million tons. Chilli accounts for roughly 17% of the global spice trade and about 33% in India due to its high economic value and high consumption rate [2]. Tamil Nadu, Karnataka, Uttar Pradesh, and the Deccan Plateau are additional commercially important capsicum-producing regions in India. However, in comparison to the United States, Holland, France, and other capsicum-producing countries throughout the world, there is insufficient supply resulting in lower production rates and limited areas under cultivation. Furthermore, Phytophthora and Colletotrichum spp. produce damping-off and anthracnose, respectively, which are severe devastative and destructive diseases that drastically lower prospective crop yields [3]. Damping-off is a Phytophthora-caused soil-borne fungal disease of seedlings that can be the misery of amateur seed growers that develops on the seeding table while young plants are just starting to grow. Seeds may rot before germinating or seedlings may perish before emergence during pre-emergence damping off. The immature plant seedlings develop rot at the crown if a post-emergence damping-off is present. Later, the tissue becomes soft, causing the plant to wilt and collapse over at the base [4]. Anthracnose, often known as ripe- fruit rot, is another frequent disease caused by the fungus Colletotrichum. Within just a few days, the disease reduces the output of ripened sweet pepper fruits and transforms them into rotten garbage [3].
Excessive use of chemical fertilizers (CF) and fungicides can induce various environmental risks as well as carcinogenic and mutagenesis effects in living beings when dealing with these soil-borne diseases and fungal pathogens. Plant protection is thus essential for maintaining long-term crop yields [5, 6, 7]. Alternatively, to address production losses caused by serious diseases, biological control measures must be considered (BCAs). Biofertilizers and biocontrol agents are a secure and reliable alternative to chemical fertilizers in the battle against plant pathogens [7, 8, 9]. Plant growth-promoting rhizobacteria (PGPR) such as Bacillus, and Pseudomonas are well-known for their biological control of soilborne plant pathogens. They colonize the roots of plants and have both direct and indirect impacts. Increased nitrogen uptake, indoleacetic acid (IAA) synthesis, phosphate solubilization, inhibition of soilborne diseases by creating siderophores, and other mechanisms of PGPR promote plant growth (PG) and yield [6, 7, 10, 11, 12]. PGPRs are rhizosphere inhabiting microorganisms that can improve phytoremediation effects. PGPR applied in contaminated soils, improves plant resistance to heavy metals, speeds up nutrient turnover, mitigates the negative effects of some metals, increases plant tolerance to pests and pathogens, and improves soil structure [7, 13, 14]. Exudates from plant roots provide a range of metabolites and nutrients which are necessary for rhizobacteria to thrive [15]. PG and yield can be increased using a variety of PGPR functions. They have the ability to fix nitrogen from the air, which is linked to the production of various growth-promoting plant hormones [7, 16]. Over 90% of the world’s plant species have symbiotic relationships with AM fungi [13].
The link is especially critical for agricultural plants, which require a lot of nutrients and water to develop and produce at their best. The AM fungi build a network of thin filaments that attach to plant roots and excrete strong nutrients that break down mineral nutrients, absorb water, inhibit soil pathogens, and bind soil particles together into a porous structure. Different chemical processes reduce the availability of phosphorus for plants, especially in arid and semi-arid soils [12]. AM fungal biofertilizers are a low- cost, long-term solution for improving plant nutrition, yield output, and reducing disease and pests [13, 17]. Although AM fungi are normally found in most soils, the number required for improved plant production and repair of the soil structure has declined drastically due to the overuse of CF’s insecticides, fungicides, and herbicides [18]. Farmers’ primary goals are to increase yield and improve the quality of the crop and soil health (SH). Bell pepper growers face two main issues: poor establishment and poorer production. AM fungal symbiosis can improve PG and nutrient uptake (NU), and there is a growing interest in its use. Understanding the role of mycorrhizae and how they interact with other bio inoculants would help to enhance inoculation techniques and planning so that the benefits of mycorrhizal association can be maximized. Furthermore, inoculating plants with PGPR and AM fungi promotes dry matter deposition and water moisture uptake, enhancing plant tolerance to stressors such as drought and salinity as shown in Figure 1. Utilizing them for plant growth in a variety of biological habitats can make a significant contribution to organic culturing for growth enhancement and yield improvement.
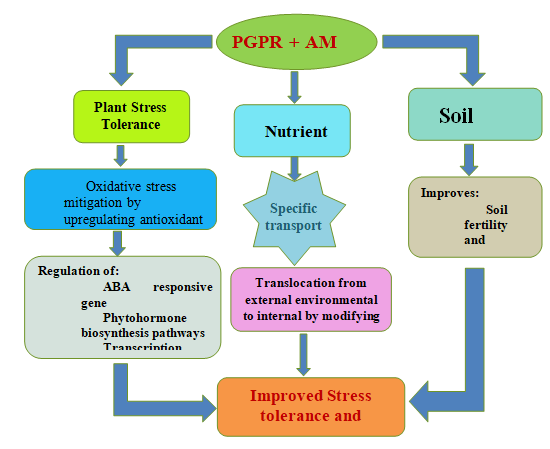
Figure1: A diagrammatic representation of PGPR and AM fungi to regulate various processes in the ecosystem and plant growth promotion under abiotic stress conditions.
Nutrient Acquisition by Way of PGPR and AM Fungi
Fertilizer requirements are often lower in soils with dynamic microbial ecologies and rich organic matter in traditionally treated soils [19]. Bulk microbial activity in soils is frequently considered while controlling the application of organic nutrient sources. Specific plant-microbe interactions that directly help in plant nutrition are beginning to emerge as a result of Phyto microbiome research [20]. Microbes that help plants acquire nutrients (biofertilizers) work in a variety of mechanisms, including increasing the surface area accessible to plant roots, nitrogen fixation, P solubilization, siderophore formation, and HCN production [21, 22]. As a result, regulating microbial activity promises a lot of potential for fulfilling crop nutritional requirements. The colonization of AM fungus is also believed to enhance nitrogen absorption in plants (figure 8). It is clear that inoculation with AM fungus can dramatically enhance the concentration of several macro and micronutrients, resulting in higher photosynthate synthesis and thus greater biomass accumulation [23, 24]. AM fungi have the potential to increase the absorption of inorganic nutrients, particularly phosphate, in practically the entire plants [7, 25]. Free-living N fixing bacteroid fixed atmospheric nitrogen and provider to plant for growth and development. After N, the next most limiting nutrient for crop plants is usually P, according to Liebig’s law of the minimum. While most agricultural soils get enough of P, the majority of it exists in insoluble forms. Furthermore, by releasing non- labile phosphorus from its refractory forms, phosphorus solubilizing microorganisms (PSMs) can assist plants in accessing the reservoir of non-labile phosphorus. Organic acids or H+ ions secreted by PSMs can solubilize inorganic P complexed with Ca, Fe, or Al. Many scientists have documented the involvement of AM fungi in the uptake of soil nutrients, particularly N and P, which can help host plants develop more successfully [7, 18, 26, 27, 28]. AM fungi have been proved to sustain P and N absorption, supporting plant development at higher and lower P levels under various irrigation regimes [29, 30]. AM fungi are also useful for assisting plants in absorbing nutrients from nutrient-deficient soils [31]. Apart from macronutrients, the interaction of AM fungus has been shown to boost the phyto-availability of micronutrients such as zinc and copper [7, 11, 12, 32]. The surface absorption capacity of host roots is improved by AM fungus [33]. Crop yields might be limited by other nutritional elements such as Fe and Zn. Fe and P can be plentiful in soils yet unavailable to plants. Many bacterial species produce organic acids or siderophores, which increase the availability of Fe [7, 12, 34, 35, 36, 37, 38, 39]. To evaluate the influence of multiple consortium bio inoculants PGPRs and most abundant AM fungi, on growth, quality parameters, yield, and nutrients uptake response of capsicum individually and along with a consortium of rhizobacteria and mycorrhizal inoculants in non-sterilized soils under low cost protected conditions of North India.
Materials and Methods
Location, Climate, and Soil of Experimental Site
The protected cultivation experiment was performed at Delhi NCR, Gurugram, and Haryana, India from July to November. The experimental site is positioned at 28.4211° north, 77.1109° East longitude, and its elevation from sea level is around 300 meters. The weather is sub-tropical with a mean maximum temperature ranging from 20.8 - 40.3°C in summer and a mean minimum between 5-17.7°C in winter. The mean annual rainfall is around 714 mm. The experimental site soil is well-drained sandy loam having good percolation and excessive fertility. The major portion of this location is under the cultivation of different agricultural, horticultural, medicinal, and aromatic crops.
Soil Sampling and Analysis
Soil samples (0-30cm) were collected randomly from each site using the standard conning and quartering method before (Initial time) and at harvest. Air-dried soil samples were used for different Physico-chemical analyses. pH was determined in 1.25 (w/v) solutions of dried soil samples in water and the same was used for the determination of electrical conductivity (EC). Air-dried soil samples were processed (addition of 40% NaOH and distillation) in a Kel Plus Nitrogen estimation system (Class DX, Pelican Equipment’s) followed by determination of available nitrogen by titration with 0.02N H2SO4 [40, 41]. The contents of soil organic carbon (SOC) were determined using Sims’ method [42]. 1.0 g of soil was combined with 10 ml of 1 N K2Cr2O7 and 20 ml of concentrated H2SO4 in this manner. This suspension was carefully mixed and diluted to 200 ml of distilled water, and then 10 ml of H3PO4 and sodium chloride were added. Estimates of total organic carbon (OC expressed as C) are used to assess the amount of organic matter in soils. Available phosphorus was determined by the Olsen method using samples with high pH sodium bi- carbonate as extracting agent [43, 44]. Available potassium was determined in a 1N ammonium extract using a flame photometer [44, 45].
Experimental Setup
An experiment was designed to investigate the effect of Absolute multiple consortium PGPR (Azotobacter, Pseudomonas, Fraturia, Azospirillum, and Bacillus)+consortium AM fungi (Aculospora logula-15%, Glomus fasciculatum-20%, Glomus intraradices- 40%, Gigaspora margarita-15%, and Scutellospora heterogama-10%), as well as an Absolute consortium of PGPR and consortium of AM fungi, Chemical fertilizer (CF), Di-ammonium phosphate (DAP), MoP, Urea, also used as control (100% RRF) and control without CF and bioinoculant on the growth parameter such as plant height, stem girth, quality yield, and nutrients uptake of capsicum (bell pepper). In the control (100% RRF) set, no inoculum was added. In each pit single seedling of capsicum was planted and placed in protected conditions. Plants were watered regularly as per needs.
Microorganisms Applications
Microbial Inoculum Preparation
Preparation of Absolute Consortium PGPR Inoculant: PGPR microbial inoculants (Azotobacter, Azospirillum, Pseudomonas, Fraturia, and Bacillus) have been proliferated in a nutrient broth medium. Then each PGPR develops eliminated on the top of a logarithmic growth phase and becomes aseptically transferred to plastic containers, which include triple sterile talc powder and then were mixed well. PGPR concentration was adjusted to 1×108 CFU/g in all inoculants. Consortium PGPR was prepared and contained Azotobacter, Azospirillum, Pseudomonas, Fraturia, and Bacillus in the same ratio. As per the treatment combination consortium, PGPR @ 1.0 g/plant becomes inoculated across the seedling during the time of transplanting. PGPR has been additionally applied to capsicum growth stages in four equal splits dose.
Preparation of Absolute Consortium of AM fungi Inoculant: The density of Absolute consortium AM fungi mixed with triple sterile talc powder, adjusted with 3000 infected propagules (IP) per gram of inoculant containing growing subtract, infected roots bits, and hyphal and mycelial biomass. AM fungi inoculum contained Aculospora logula-15%, Glomus fasciculatum-20%, Glomus intraradices-40%, Gigaspora margarita-15% and Scutellospora heterogama-10% infective propagules (IP). AM fungi @500 IP/ plant were applied as a root dipping method at the time of planting. After planting of capsicum, necessary irrigation was given to all treated and untreated plots.
Chemical Fertilizers (CF)
Various level of CF was applied in control treatment in the form of DAP 1.08 q ha-1, MoP 1.0 q ha-1, and Urea 2.275 q ha-1 as 100% recommended rate of fertilizers (RRF) as farmers practiced. Urea was also applied as a top dressing in equal splits as per recommendations.
Field Preparation, Nursery Raising, and Experimental Design
The experimental soil was opened with a power tiller and kept exposed to the sun prior to the next plowing. It was prepared afterward by plowing and cross plowing followed by laddering. Well decomposed organic manure at the rate of 20 kg m-1 was mixed with all soil. Raised beds were formed after bringing soil to a fine tilth. The bed size was 100 cm wide and 20 cm in height. Between the beds, 50 cm walking space was prepared. Capsicum (Capsicum annuum, family Solanaceae, Capsicum Hybrid Intruder) seedlings were prepared on an experimental field site. Capsicum seeds were treated with Trichoderma species before sowing in the seeding pro tray filled with fine sterilized cocopeat. After 30 days, the uniform size of capsicum seedlings was selected and treated with and without consortium PGPR/AM fungi and transplanted in experimental plots with a spacing of 60cm X 70cm. Planting was done in the raised bed in a pit size of 15cm x 20cm. AM fungi and PGPR treatment were considered as Absolute consortium AM fungi alone, Absolute consortium PGPR alone, and Absolute consortium PGPR + AM fungi and compared to control treatment without microbial inoculant (100% RRF) and control with microbial inoculant and RRF. The plantation was finished in the first week of July and the size of the experimental plot was 2.5X2.5 meters with four replicate in each treatment in a randomized complete block design (RCDB). 100% RRF was added to control treatments only. Recommended cultural operations (RCO) were carried out during the entire cropping period to ensure a healthy crop. The Physico-chemical properties of the soil were estimated at the Initial time and at harvest (Figures 2-7). Nutrient uptake in the shoot was determined after harvest (Figures 8-13).
Morphological and Yield Attributes of Trials
Data Collected and Measurements
The effects of different microbial inoculations observations, randomly 10 plants were selected after one meter of each plot in each replicates for all the characteristics such as the plant height (cm), Stem girth (cm), number of fruit per plant, fruit length (cm), fruit diameter (cm), and fruit weight (g) as well as grass yield (q ha-1).
Plant Height and Stem Girth (cm): Measured from the ground level to the top of a matured plant at 30, 60, 90,120, and 150 days after transplanting (DAT).
No. of Fruit/plant: The number of fruit per plant was counted in different intervals and the average was calculated.
Fruit Length (cm): The average length of the fruit of the plants measured at maturity and expressed in centimeters.
Fruit Diameter (cm): The average fruit diameter was measured using a veneer caliper and expressed in centimeters after harvest.
Fruit Weight (g): Fruit weight was measured using digital balance and recorded and expressed in gm per fruit.
Total Fruit Yield (q ha-1): The marketable and healthy capsicum fruit yields were expressed in q ha-1.
Analysis of TSS
Total Soluble Solids (TSS)
The TSS was determined from fifteen randomly selected bulbs using the procedures [46]. Aliquot juice was extracted using a juice extractor and 50 ml of the slurry was centrifuged for 15 minutes. The TSS was determined by hand refractometer (ATAGO TC-1E) with a range of 0 to 32° Brix and resolutions of 0.2° Brix by placing 1 to 2 drops of clear juice on the prism, washed with distilled water, and dried with tissue paper before use. The refractometer was standardized against distilled water (0% TSS). Amount of total soluble solids present in the bulb expressed in percentage.
Assessment of Shoot Macro and Micronutrient Uptake
Randomly four places were selected for plants samplings in all capsicum plots. For nutrient analysis of the shoot systems, the oven-dried samples were finely ground. Nitrogen (N) in the shoots was determined using an elemental analyzer (EA 3000, Eurovector, Italy). To estimate the phosphate (P) and potassium (K) level in the shoots, 1g of the finely ground sample was subjected to a wet oxidation treatment using tri-acid (HNO3:H2 SO4:HClO4; 10:1:4) digestion in a digestion block (KELPUS, KES121; Pelican Equipment, Chennai, India) at 200oC. Following acid digestion, the samples were diluted and filtered for further nutrient analysis. Shoots P was determined by the Vanado molybdophoshoric acid colorimetric method [47] using a spectrophotometer (Specord 200; Analytik Jena, Germany). K was measured by the ammonium acetate method of Hanway and Heidel [48] by using a flame photometer (Model FP114; Thermo Scientific, USA). To determine iron (Fe), copper (Cu), and zinc (Zn) content in shoots samples were digested in a microwave (Mars 5, CEM). Following the US EPA 3051A method (US EPA 2007), the metal concentration in the acid digestive was determined using atomic absorption spectrophotometer (AAS) (SOLAAR, TJA Solution, UK.
Assessment of Mycorrhizal Root Colonization (MRC) Percentage in Capsicum Root System
Assessment of mycorrhizal root colonization (MRC) percentage in the capsicum root system, approximately 1-2 g of freshly collected fine roots were used for staining and the assessment of MRC percentage. Roots were washed in freshwater, cleared with 10% KOH, acidified with 1N HCl, and stained with 0.05% Trypan blue [49]. Quantification of root colonization for AM fungi was conducted using the gridline intersection method [50] and 100 segments of each sample were observed under a compound microscope (Leica DM750). The presence or absence of AM fungal structure in the root system such as vesicles, arbuscules, and hyphae at particular fixed points was recorded, and the results were expressed as a percentage MRC of observations.
Statistical Analysis
All results were analyzed using Analysis of Variance (ANOVA), followed by a post hoc test through computer software SPSS 11.5 version. Means were then ranked at a P<0.0 level of significance using Duncan’s Multiple Range Test for comparison.
Results
Physico-chemical Properties of Soil (Initial time and at Harvest)
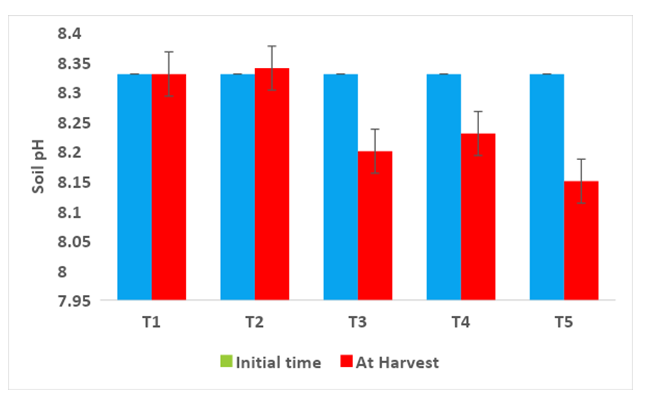
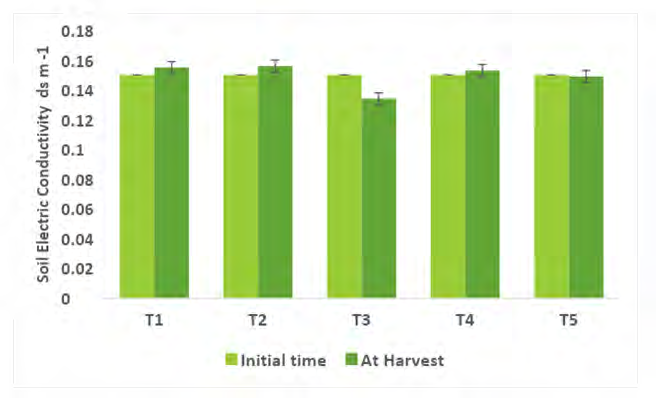
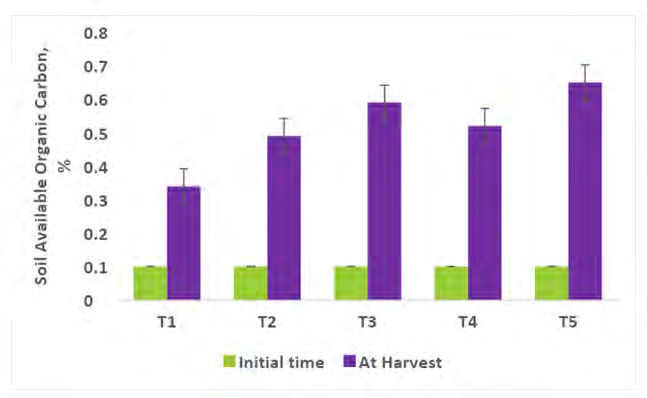
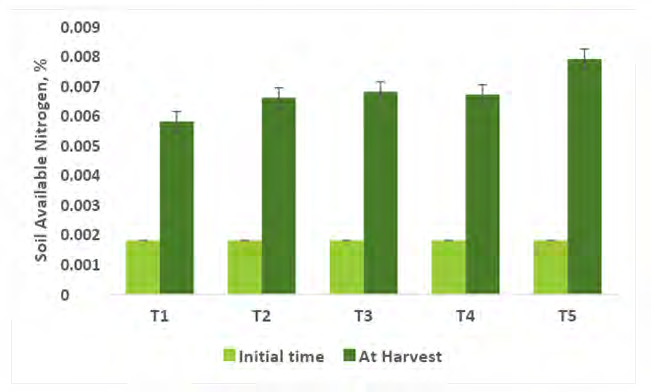
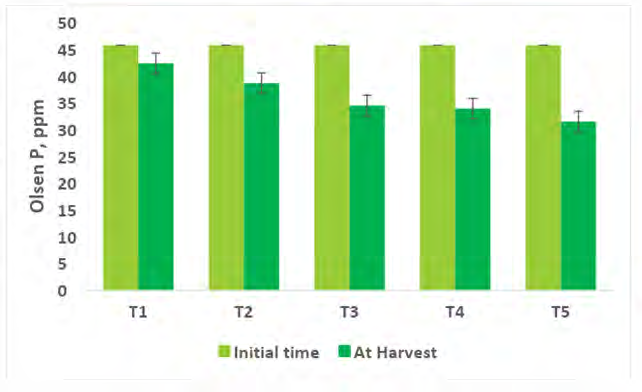
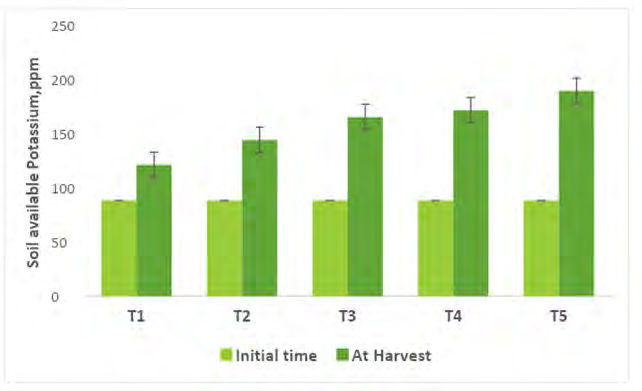
Comparing the Physico-chemical properties of the soil initial time and at harvest of experimental crops were presented in Figures 2-7. A slight alteration of pH was recorded in treatments (Figure 2). Microbial-mediated capsicum showed a significant (P<0.05) decrease in electrical conductivity (Figure 3). The factors are responsible for the change in pH and EC in the continuous variations of equilibrium between cations and anions present in the soil. Plant uptake of soluble salts by crops and or leaching of cations such as calcium, and magnesium can decrease the pH and at the same time chloride accumulation on the surface due to capillary action can be responsible for the decrease in EC [7, 11, 12, 51, 52]. Available N in the soil is directly associated with soil organic matter (SOM). The gradual increase in N is due to an increase in SOM and the microbial activities (MA) which make N available from organic matter (OM) to microbial inoculants treated plots. A maximum significant increase (P<05) in available N (0.0079%) was recorded from multiple Absolute consortium PGPR + AM fungi treated plots (Figures 4 & 5) and minimum in control (0.0058%). Available P content was noticed significantly lower in treatment T5 (31.45ppm) as compared to treatment T1 (43.24ppm) soil and it may be due to P mobilizing activity of mycorrhiza, added during plantation activities (Figure 6). A sharp decrease in K content in soil was noticed in all microbial treated capsicum compared to control (Figure 7). Maximum decrease of K has been recorded in Absolute consortium PGPR + AM fungi treatment T5 (189.67ppm) and minimum in control (121.25 ppm) as compared to initial time (88.25ppm). Various factors including weathering, upward translocation of soluble ions through capillary action, and involvement from the degradation of plants litters can be responsible for such variation of K content in different treatments of capsicum crop [11, 12, 14, 53].
Measurement of Plant Growth Parameter (MPGP)
The effect of the Absolute consortium PGPR, Absolute consortium AM fungi single and in combination, and CF on the growth and yield of capsicum differed significantly (Tables 1-4). Change in plant height was significant in all the treatments and maximum plant height was observed in the Absolute consortium PGPR + AM Fungi followed by Absolute consortium of AM Fungi and Absolute consortium of PGPR alone i.e., 35.76-116.70 cm, 31.03-111.97 cm, and 27.6o-108.60 cm respectively, while CF (100% RRF) had 22.36-103.37 cm height, which was much higher than the uninoculated plants (control) having 99.30 cm, after 30- 150 DAT from the DAT of capsicum (Table 1). A significant difference was observed between the treatments. Additionally, multiple Absolute consortium PGPR+ AM Fungi resulted from the maximum increase in capsicum plant stem girth followed by an Absolute single consortium of AM fungi and a single consortium of PGPR such as54.32 mm, 54.21 mm, and 54.15 mm respectively, while CF (100% RRF) having 53.29 mm, whereas uninoculated plant (control) having 52.29 mm stem girth, 150 DAT, which was very poor in comparison of the multiple consortia of PGPR + AM fungi and alone consortium of AM fungi or PGPR (Table 2). Statistically significant (P<0.05) differences were observed between the treatment. Similarly, the multiple Absolute consortium PGPR + AM Fungi in capsicum has the maximum weight per fruit followed by a single consortium of AM fungi and a single consortium of PGPR i.e., 224 gm, 216 gm, and 207.0 gm respectively, while CF (100% RRF) having the 196.0 gm, apart from that uninoculated plant (control) of capsicum having the 173.0 gm weight of fruit. While the maximum number of fruits/plant, fruit diameter, and fruit length were also found in the multiple Absolute consortium PGPR + AM Fungi, 8.20, 7.81 cm, and 8.5 cm respectively, followed by single consortium AM fungi (7.89 cm, 6.1 cm, and 8.03 cm) and single consortium PGPR was 6.80, 7.82 cm and 5.98 cm whereas CF (100% RRF) was 6.10cm, 5.98 cm, and 7.53 cm respectively, apart from that uninoculated plant (control) was a weight of fruits 173.0 gm, a number of fruits per plant 4.90, fruit diameter 4.63 cm and fruit length 6.47 cm which was very low in comparison to the multiple Absolute consortium PGPR + AM Fungi and single consortium AM fungi or consortium PGPR (Table 3).
| Treatment | Plant Height (cm) | ||||
|---|---|---|---|---|---|
| 30 DAT | 60 DAT | 90 DAT | 120 DAT | 150 DAT | |
| Control | 18.46 | 32.87 | 49.47 | 71.77 | 99.3 |
| Control (100% RRF) | 22.36 | 36.77 | 53.37 | 75.77 | 103.37 |
| Consortium AM Fungi | 31.03 | 45.37 | 61.97 | 84.37 | 111.97 |
| Consortium PGPR | 27.6 | 42 | 58.6 | 81 | 108.6 |
| Absolute Consortium PGPR+ AM Fungi | 35.76 | 50.1 | 66.73 | 89.1 | 116.7 |
| F Test | S | S | S | S | S |
| C.D.at 5% | 2.12 | 2.21 | 2.22 | 2.36 | 2.29 |
| Treatment | Stem Girth (mm) | ||||
| 30 DAT | 60 DAT | 90 DAT | 120 DAT | 150 DAT | |
| Control | 11.66 | 21.78 | 33.96 | 45.34 | 52.94 |
| Control (100% RRF) | 11.99 | 22.11 | 34.29 | 45.69 | 53.29 |
| Consortium AM Fungi | 13.01 | 23.13 | 35.31 | 46.71 | 54.21 |
| Consortium PGPR | 12.64 | 22.76 | 34.94 | 46.34 | 54.15 |
| Absolute Consortium PGPR+ AM Fungi | 13.37 | 23.49 | 35.7 | 47.07 | 54.32 |
| F Test | S | S | S | S | S |
| C.D.at 5% | 1.23 | 1.77 | 1.79 | 1.69 | 1.7 |
Table 1: Quantity parameters of capsicum influenced by different doses of biofertilizers under Low-Cost Protected Cultivation. DA
Table1: Plant height of Capsicum under Low-Cost Protected Cultivation (n=10). DAT-Days after transplanting
Table2: Morphological observation of capsicum stem girth (mm) under Low-Cost Protected Cultivation (n=10). DAT-Days after transplanting
| Treatment | Quantity Parameters | |||
|---|---|---|---|---|
| No. of fruit/plant | Fruit length (cm) | Fruit diameter (cm) | Weight Fruits (g) | |
| Control | 4.9 | 6.47 | 4.64 | 173 |
| Control (100% RDF) | 6.1 | 7.53 | 5.88 | 196 |
| Consortium AM Fungi | 7.89 | 8.03 | 6.1 | 216.99 |
| Consortium PGPR | 6.8 | 7.82 | 5.98 | 207.99 |
| Absolute Consortium PGPR+ AM Fungi | 8.2 | 8.5 | 7.81 | 224 |
| F Test | S | S | S | S |
| CD at 5% | 0.02 | 0.46 | 0.35 | 0.38 |
Table 2: Quantity parameters of capsicum influenced by different doses of biofertilizers under Low-Cost Protected Cultivation. DA
Table 3: Quantity parameters of capsicum influenced by different doses of biofertilizers under Low-Cost Protected Cultivation. DAT-Days after transplanting In a similar trend, the multiple consortium PGPR + AM Fungi showed the maximum yield and quality parameter (Fruit Yield (1.83 kg plant-1), total Fruit Yield (742.5 q ha-1), Shelf life7.10 days, and TSS (5.63, Brix0), followed by single consortium AM Fungi (Fruit Yield kg plant- (1.64 kg plant-1), total fruit yield (622.85 q ha-1), Shelf life 6.70 days and TSS (5.60 Brix0) and a consortium of PGPR (fruit yield (1.33kg plant-1, total fruit yield (537.3 q ha-1), Shelf life (6.5 days and TSS(5.55 Brix0). CF (100% RRF) (Fruit Yield,1.32 kg plant-1), total fruit yield 535.95 q ha-1, shelf life 6.2days, and TSS (5.52 Brix0), but uninoculated plants (control) showing fruit yield (0.84 kg plant-1), total fruit yield (341.55 q ha-1), shelf life 5.40 days and TSS (5.34 Brix0) which was very poor in compared with multiple consortium PGPR + AM Fungi and single consortium PGPR or AM fungi (Table 4). Significant differences were recorded between the treatments and the control. Prasad K [11] reported that when S. tuberosum cultivars (Kufri pukhraj, Kufri sindhuri, and Kufri laukar) and tomato were inoculated with consortium AM fungal inoculant (Aculospora logula, Glomus fasciculatum, Glomus intraradices, Gigaspora margarita, and Scutellospora heterogama), the percent of edible tubers and tomato increased due to improvement of root volume and macro and micronutrients absorption by plants. The yield of capsicum can be affected by the interaction of mycorrhizae, PGPR, and fertilizers (Table 4). Mean assessments indicated that in mycorrhiza and PGPR inoculated capsicum with increasing of K, P and other minerals rate than capsicum yield increased. The highest yield was noticed in Absolute consortium PGPR + AM fungi treated cucumber followed by consortium AM fungi, consortium PGPR, and the lowest in control (Table 4). It is widely assumed that the capsicum plant benefits positively from PGPR and AM fungi symbiosis [46, 54, 55], it makes little growth without mycorrhiza unless heavily fertilized [7, 11, 13, 56, 57]. The consortium PGPR and AM fungi alone and with combination-treated capsicum performed better than untreated control (100% RRF) and control. Significant differences were recorded between the treatments. Consortium PGPR and AM fungi-treated capsicum showed a significant increase in plant height, and the number of leaves/plant, compared to non-microbial control (100% RRF, Farmer’s practiced). Multiple consortium PGPR and AM fungi association has also positively correlated with plant growth and productivity. It is expected that the capsicum plant benefited positively from Pseudomonas, Azotobacter, Fraturia, Azospirillum, Bacillus PGPR microbes, and AM fungi symbiosis in an early application.
| Treatment | Yield Parameters | |||
|---|---|---|---|---|
| Fruit Yield kg plant- 1 | Fruit Yield q ha-1 | Shelf life (Days after harvest) | TSS (Brix0) | |
| Control | 0.84 | 341.55 | 5.4 | 5.34 |
| Control (100% RDF) | 1.32 | 535.95 | 6.2 | 5.52 |
| Consortium AM Fungi | 1.64 | 662.85 | 6.7 | 5.6 |
| Consortium PGPR | 1.33 | 537.3 | 6.5 | 5.55 |
| Absolute Consortium PGPR+ AM Fungi | 1.83 | 742.5 | 7.1 | 5.63 |
| F Test | S | S | S | S |
| CD at 5% | 0.015 | 1.16 | 0.55 | 0.17 |
Table 3: Yield and quality parameters of capsicum influenced by different doses of biofertilizers under Low-Cost Protected Cultiv
Macronutrients Translocation by Capsicum Shoot System
Nitrogen Translocation by Capsicum Shoot System
PGPR and AM fungi can improve plant growth by improving macro and micronutrient absorption. To determine the effect of multiple consortiums of PGPR + AM fungi as well as a single consortium of PGPR or AM fungi inoculants on the uptake of macronutrients state of pepper plants and its detoxifying role, N, P, K, were examined (Figures
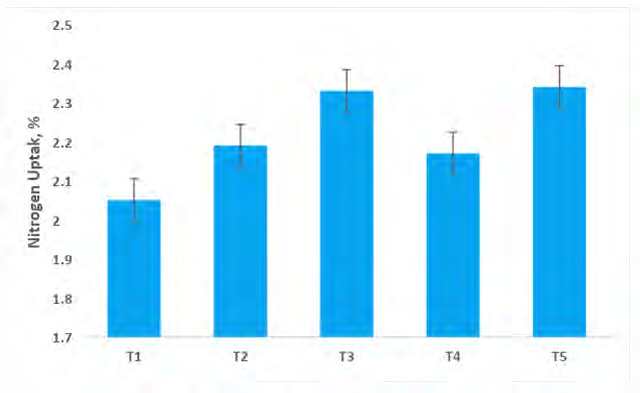
8-10). An assessment of data indicates that N translocation (Figure 8) through capsicum shoot systems shows that all the microbial treatments had a significant influence on N uptake as compared to NMC (100% RRF) and control treatment. The maximum N uptake (2.34% at harvest) was obtained under treatment T5 (Absolute consortium PGPR + AM fungi) where the six microbial inoculants were applied. However, the lowest value of N uptake (2.5% at harvest) by capsicum shoot was recorded under control treatment. The uptake of N by the capsicum plants went on increasing with the successive microbial application because the uptake is a resultant of strength and biological yield.
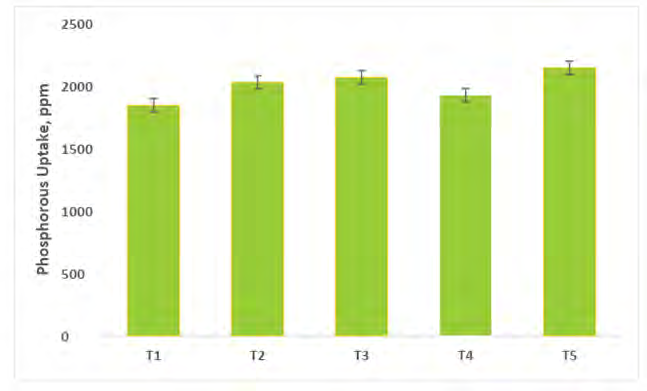
Phosphorus Translocation by Capsicum Shoot System
A glance at data in figure 9 shows the highest uptake of P (2145ppm) by the capsicum under treatment T5 (consortium PGPR + AM fungi). The minimum P uptake was recorded under T1 (1845.12 ppm at harvest). The effect of microbial consortium PGPR + AM fungi inoculation on P uptake was significant (Figure 9). P uptake increased may be due to improved absorption and utilization of available soil P at higher rates.
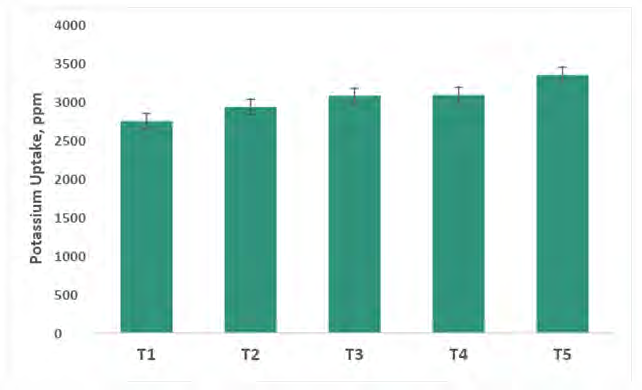
Potassium Translocation by Capsicum Shoot System
Potassium translocation in capsicum plants has been presented in figure 10. An inquisition of data indicates that maximum K uptake (3345 ppm) by capsicum shoot was recorded in consortium PGPR + AM fungi treatment, where the six consortium microbial stimulants were applied followed by T4 (consortium PGPR), T3 (consortium AM Fungi), T2 (control (100% RRF), and T1 (control). K uptake was increasing may be due to improved absorption and utilization of potassium at higher rates of available soil potassium.
Micronutrient Translocation by Capsicum Shoot System
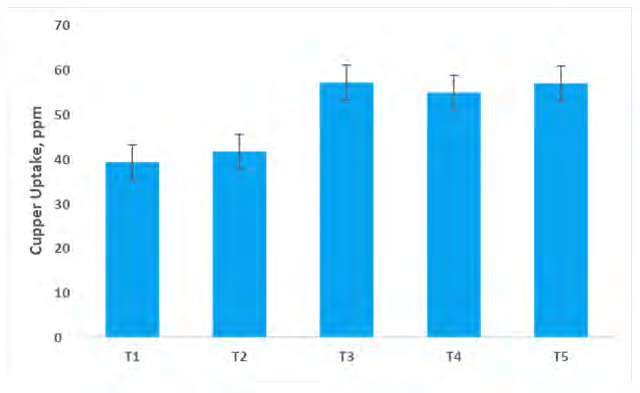
Copper Translocation by Capsicum Shoot System
The glimpse of data presented in Figure 11 indicates the highest uptake of copper (56.78 ppm) by the capsicum shoot system was recorded in the treatment consortium PGPR + AM fungi.The effect of six consortium bio inoculants treatments was noticed to exert a significant effect on the copper removal by capsicum plants. The minimum copper (39.1ppm) uptake was recorded under control without a treatment plant at harvest.
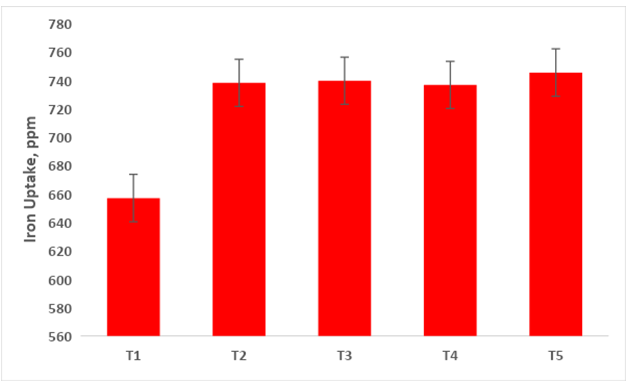
Iron Translocation by Capsicum Shoot System
The perusal of data presented in Figure 12 reveals the highest uptake of iron (745.34 ppm) by the capsicum plant was recorded in treatment T5 (Absolute consortium PGPR + AM fungi). The effect of multiple cultures of PGPR and AM fungi in treatments was noticed to exert a significant effect on the iron removal by capsicum shoot followed by control there no fertilizers and bioinoculant was applied (657ppm).
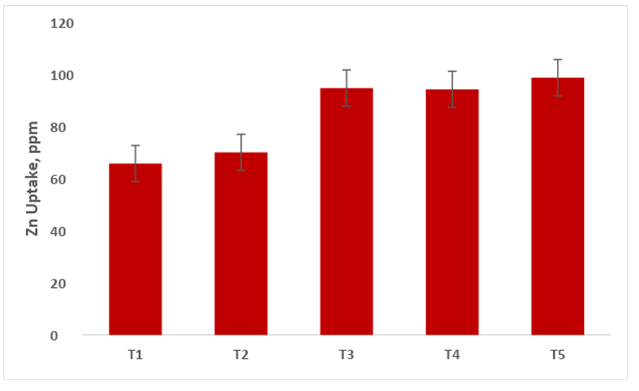
Zinc Translocation by Capsicum Shoot System
The data presented in Figure 13 reveals that maximum Zn translocation (98.78 ppm) by capsicum shoot system recorded under treatment T5 (Absolute consortium PGPR + AM fungi) followed by T3 (94.76ppm), T4 (91.31ppm), T2 (70.79ppm), T1 (65.78). The microbial inoculants such as PGPR and AM fungi facilitated capsicum plants were found to impose a significant effect on the Zn uptake.
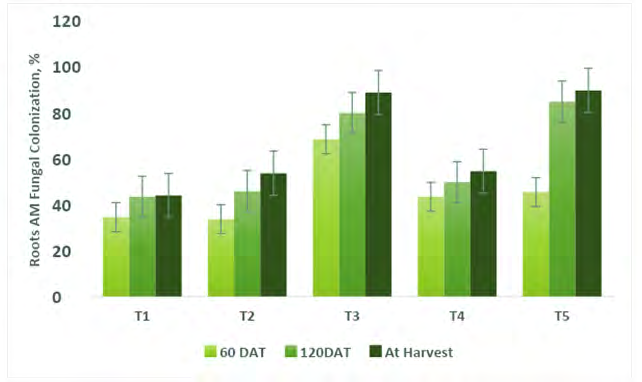
Mycorrhiza Root Colonization (MRC) Percentage In Capsicum
MRC percentage in the capsicum root system was observed in all treatments including control plants (Figure 14). Maximum 89.56% of MRC was observed in T5 (Absolute consortium PGPR + AM fungi) bioinoculant mediated capsicum crop followed by T3 (Absolute consortium AM fungi, 88.56%), T4 (Absolute consortium PGPR, 54.45%), T2 (control, 100% RRF, 53.56%) and T1 (control, 43.89%) plants at harvest (Figure 15). Statistically significant (P<0.05) differences were observed between the treatment. MRC was increased with additionally added PGPR with mycorrhizae. In general, AM fungal strains arbitrated plants have been encouraged by higher water and mineral nutrients uptake from the soils because they increased the total root surface [6, 11, 13, 18, 58, 59]. The colonization potential of AM fungi decreases in control treatment due to 100% RRF applied in capsicum whereas increases in PGPR application. MRC percentage was affected with an added in PGPR and values were statistically different compared to NMC (100% RRF) and control [58]. The results in this study revealed increased plant growth parameters, nutrients concentration in soil, nutrient translocation, and AM fungal root colonization percentage when consortium PGPR and AM fungi inoculated plants, and this was comparable to uninoculated plants treated with high dosages of fertilizer (100% RRF) and without fertilizer. The influence of consortium PGPR and AM fungi in decreasing fertilizer demand of major crop species was reported by [6, 11, 12, 56, 60]. It has been proven that PGPRs and AM fungi have the potential to reduce the high application rate of fertilizer needed to produce high capsicum yield [57]. Moreover, the capsicum plant benefits positively from AM fungal symbiosis. It creates small growth without mycorrhiza unless severely fertilized [55, 61]. Reduction in plant growth characteristics, shoot nutrient content, root colonization, and yield of control plant with improvement in fertilizer function. From the above results, it appears that capsicum should be integrated with a consortium of Azotobacter, Pseudomonas, Fraturia, Azospirillum, Bacillus in combination with a consortium of AM fungi for better growth, yield, and quality. For increasing capsicum shelf life, the combination of PGPR + AM fungi is effective. Though the Absolute consortium PGPR + AM fungi bioinoculant produced the best result compared to different combinations of biofertilizers and recommended rate of CFs. The next may be a particular level with certain considerations of sustainability in production and environmental protection.
Discussion
In general, capsicum responded better to different treatments for different characters as compared to the control. In this experiment, none of the single inoculation treatments had a significant effect on all of the experimental parameters or showed consistent plant growth-promoting effects as observed in multiple bio inoculation; however, the observed growth parameters were found to be highest in both the consortium PGPR + AM fungi and the alone consortium PGPR and AM fungi for inoculating capsicum plants. AM fungi are known to have the capacity to increase plant nutrient intake by creating an association with roots [62], as well as promote the growth of other rhizospheric microbes (PGPR), resulting in increased PG [63]. Another reason could be that once the AM fungus invades the host roots, they alter the root exudates produced and create the phosphatase enzyme in the rhizosphere. Extraradical hyphae of AM fungus produced phosphatases that could hydrolyze extracellular phosphate ester linkages, making P accessible to plants [13, 18, 64]. The root system was shown to be considerably more compatible with the consortium AM fungi + consortium PGPR in terms of absorbing and translocating nutrients through a large consortium. AM fungi aid in the uptake of diffusion-limited nutrients such as P, Zn, Cu, and others, provide drought resistance and pathogen protection, produce plant growth regulators, and interact synergistically with PGPR [7, 65].
Improved plant growth may lead to an increase in fresh shoot and root growth, as well as increased biomass build-up, which enhances P uptake. Because phosphorus is required for nitrogen fixation, consortium inoculation may have altered the plant’s P and N uptake in this experimentation as well. The P and N levels of the plants followed a similar pattern to that shown in previous studies on other crops [66]. Finally, the growth parameters (plant height, stem girth, yield, and quality) of the inoculated capsicum plant were considerably higher than those of the uninoculated (control) plant. This is owing to the presence of AM fungus, as well as the PGPR’s efficiency. Plant pathogens are protected by phytohormones produced by PGPR. In this study, it was found that inoculating the field with mixed consortium AM fungus + PGPR at the time of transplanting not only improved plant growth and production but also reduced CF application and improve SH as well as reduced pollution and save the environment.
Conclusion
Based on the findings; maximum growth and quality yield were noticed in Absolute consortium PGPR + AM fungi treatment followed by the single biological consortium PGPR, AM fungi NMC, and control treatments. On the other hand, these treatments showed had the maximum nutrients uptake by the shoot. A significant difference (P<0.05) was noticed in nutrient uptake by capsicum. It has been determined that the maximum utmost was within the consortium PGPR + AM fungi treatment and the minimum was discovered within the PGPR treatment that was higher than NMC (100% RRF) and control treatment. One of the physiological processes which can markedly alter or cut back the nutritionary quality of the various plant products consumed by humans is oxidative stress. The environmental factors that induce oxidative stress in plants include air pollution, herbicide/ pesticide utilization, heavy metal contamination, drought, salinity, injuries, UV light, unfavorable temperatures, and photoinhibition from excessive solar radiation [67, 68]. The utilization of the consortium of multiple PGPR and AM fungi could also be due to the ability to reduce the negative effects of environmental stress and improved capsicum quality. The production of plant growth regulators (PGR) by PGPR microorganisms is another vital mechanism usually related to growth stimulation [69]. AM fungi are identified to have an effect on PG and health by increasing resistance to the tolerance of biotic [70, 71] and abiotic stress [11, 72].
Based on the responses of different characters such as PG, nutrient uptake (P, N, K, Zn, Fe, Cu), and yield, it can be concluded that the multiple consortium PGPR + AM fungi are the best consortia of microorganisms for inoculating capsicum plants, followed by the single consortium AM fungi and the consortium PGPR. Inoculation with a microbial community like this could result in healthy, aggressively growing capsicum seedlings and plants. Because it is simple and environmentally safe any nurseryman or farmer can use it to inoculate capsicum seedlings in the nursery and in the field. As a result, this study suggests that farmers with similar soil and other environmental conditions add Absolute consortia PGPR + AM fungi to their farms and fields at the transplanting stage instead of other dangerous chemicals to improve crop establishment, production, and nutrients. This development combines long-term control with pathogen control and provides ISR with a defense mechanism.
Acknowledgment
The author is grateful to Absolute Biologicals, Life Science Plant Research, Division of Microbiology, Delhi NCR, Gurugram, Haryana, for providing laboratory facilities and financial assistance.
References
-
Kaushal M, Mandyal P, Kaushal R (2019) Field-Based Assessment of _Capsicum annuum_ Performance with inoculation of Rhizobacterial Consortia. Microorganisms 7(3): 89.
-
Datta M, Palit R, Sengupta C, Pandit KM, Banerjee S (2011) Plant growth-promoting rhizobacteria enhance the growth and yield of chilli (_Capsicum annuum_ L.) under field conditions. Australian Journal of Crop Science 5(5): 531-536.
-
Sahitya UL, Deepthi RS, Kasim DP, Suneetha P, Krishna MSR (2014) Anthracnose a prevalent disease in _Capsicum_. Research Journal of Pharmaceutical, Biological and Chemical Sciences 5: 1583-1604.
-
Than PP, Prihastuti H, Phoulivong S, Taylor PWJ, Hyde KD (2008) Chilli anthracnose disease is caused by _Colletotrichum_ species. J Zhejiang Uni Sci B 9(10): 764- 778.
-
Tariq M, Hameed S, Malik KA, Hafeez FY (2007) Plant root-associated bacteria for zinc mobilization in rice. Pak J Bot 39(1): 245-253.
-
Prasad K (2021a) Impact of Biological Fertilizer Arbuscular Mycorrhizal Fungi and Conventional Fertilizers Mobilization on Growth, Yield, Nutrient’s uptake, Quercetin and Allin Contents in _Allium_ Crops Cultivation under Field Conditions in Semi-Arid Region of India. South Asia Journal of Experimental Biology 11(1): 15-26.
-
Prasad K (2022) Influence of PGPR, AM Fungi and Conventional Chemical Fertilizers Armament on Growth, Yield Quality, Nutrient’s translocations and Quercetin Content in Onion Crop Cultivated in Semi-Arid Region of India. Journal of Microbiology and Biotechnology 7(1): 1-18.
-
Hassen W, Neifar M, Cherif H, Najjari A, Chouchane H, et al. (2018) _Pseudomonas rhizophila_ S211, a New Plant Growth Promoting Rhizobacterium with Potential in Pesticide-Bioremediation. Front Microbiol 9: 34.
-
Prasad K, Warke RV, Khadke K (2019) Management of soilborne pathogens to improve the production of pulses using organic Technologies for sustainable agriculture. International Journal of Research and Analytical Reviews 6(2): 82-101.
-
Sang MK, Kim KD (2012) Plant growth-promoting rhizobacteria suppressive to Phytophthora blight affect microbial activities and communities in the rhizosphere of pepper (_Capsicum annuum_ L.) in the field. Applied Soil Ecology 62: 88-97.
-
Prasad K (2021b) Influence of arbuscular mycorrhizal fungal biostimulants and conventional fertilizers on some solanaceous crops for growth, productivity and nutrient stoichiometry under field conditions in the semi-arid region of Maharashtra, India. Journal of Experimental Biology and Agricultural Sciences 9(1): 75-86.
-
Prasad K (2021c) Stimulation Impact of Rhizospheric Microbe’s Glomeromycota AM Fungi and Plant Growth Promoting Rhizobacteria on Growth, Productivity, Lycopene, Β-Carotene, Antioxidant Activity and Mineral Contents of Tomato beneath Field Condition Cultivated in the Western Ghats Covering Semi-Arid Region of Maharashtra, India. Journal of Bioscience & Biomedical Engineering 2(3): 1-14.
-
Prasad K (2017) Biology, Diversity and Promising Role of Mycorrhizal Entophytes for Green Technology. In: Maheshwari DK (Ed.), Endophytes: Biology and Biotechnology. Volume 1, Series Sustainable Development and Biodiversity 15, Springer International Publishing AG, Switzerland, pp: 267-301.
-
Prasad K (2021d) Diversification of Glomermycota forms Arbuscular Mycorrhizal Fungi Associated with Vegetable Crops Cultivated underneath Natural Ecosystems in Arid Region of Rajasthan, India. Current Investigations in Agriculture and Current Research 9(2): 1205-1212.
-
Han J, Sun L, Dong X, Cai Z, Sun X, et al. (2005) Characterization of a novel plant growth-promoting bacteria strain Delftia tsuruhatensis HR4 both as a diazotroph and a potential biocontrol agent against various plant pathogens. Syst Appl Microbiol 28(1): 66- 76.
-
Gupta S, Pandey S (2019) Unravelling the biochemistry and genetics of ACC deaminase- An enzyme alleviating the biotic and abiotic stress in plants. Plant Gene 18: 100175.
-
Prasad K (2015) Biofertilizers: A new dimension for agriculture and environmental development to improve production in a sustainable manner. Journal of Basic and Applied Mycology 11(1&II): 5-13.
-
Prasad K (2020) Positive Importance of Arbuscular Mycorrhizal Fungi for Global Sustainable Agriculture and Environment Management for Green Technology. Current Investigations in Agriculture and Current Research 9(2): 1182-1184.
-
Bender SF, Wagg C, Heijden MGVD (2016) An underground revolution: biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol Evol 31(6): 440-452.
-
Beattie GA (2015) Microbiomes: curating communities from plants. Nature 528(7582): 340-341.
-
Hafeez FY, Ullah MA, Hassan MN (2013) Plant growth- promoting rhizobacteria as zinc mobilizers: a promising approach for cereals biofortification. In: Maheshwari DK, et al. (Eds.), Bacteria in Agrobiology: Crop Productivity. Springer, Berlin, pp: 217-235.
-
Pii Y, Mimmo T, Tomasi N, Terzano R, Cesco S, et al. (2015) Microbial interactions in the rhizosphere: beneficial influences of plant growth-promoting rhizobacteria on the nutrient acquisition process. A review. Biology and Fertility of Soils 51: 403-415.
-
Chen S, Zhao H, Zou C, Li Y, Chen Y, et al. (2017) Combined Inoculation with multiple arbuscular mycorrhizal fungi improves growth, nutrient uptake and photosynthesis in cucumber seedlings. Frontiers in Microbiology 8: 2516.
-
Mitra D, Uniyal N, Panneerselvam P, Ansuman S, Ganeshamurthy AN, et al. (2019) Role of mycorrhiza and its associated bacteria on plant growth promotion and nutrient management in sustainable agriculture. International Journal of Life Sciences and Applied Science 1(1): 1-10.
-
Smith SE, Smith FA, Jakobsen I (2003) Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol 133(1): 16-20.
-
Smith SE, Jakobsen I, Grnlund M, Smith FA (2011) Roles of arbuscular mycorrhizas in plant phosphorus nutrition: interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol 156(3): 1050- 1057.
-
Prasad K (2021e) Effect of Dual Inoculation of Arbuscular Mycorrhiza Fungus and Cultivar Specific _Bradyrhizobium_ _Japonnicum_ on the Growth, Yield, Chlorophyll, Nitrogen and Phosphorus Contents of Soybean (_Glycine Max_ (L.) Merrill.) Grown on Alluvial Soil. Journal of Innovation in Applied Research 4(2): 1-10.
-
Prasad K (2021f) Potential Impact of Seed Coating with Beneficial Microorganisms to Meticulousness Sustainable Organic Agriculture for Quality Nutritive Food Production for Modern Lifestyle, Improve Global Soil and Environmental Health towards Green Technology. Aditum Journal of Clinical and Biomedical Research 2(4): 1-9.
-
Liu L, Gong Z, Zhang Y, Li P (2014) Growth, cadmium uptake and accumulation of maize (Zea mays L.) under the effects of arbuscular mycorrhizal fungi. Ecotoxicology 23(10): 1979-1986.
-
Liu C, Ravnskov S, Liu F, Rubaek GH, Andersen MN (2018) Arbuscular mycorrhizal fungi alleviate abiotic stresses in potato plants caused by low phosphorus and deficit irrigation/partial root-zone drying. Journal of Agriculture Science 156(1): 46-58.
-
Kayama M, Yamanaka T (2014) Growth characteristics of ectomycorrhizal seedlings of _Quercus glauca_, _Quercus_ _salicina_, and _Castanopsis cuspidata_ planted on acidic soil. Trees 28: 569-583.
-
Smith SE, Read DJ (1997) Mycorrhizal symbiosis. 3rd(Edn.), Academic Press, San Diego, pp: 607.
-
Bisleski RL (1973) Phosphate pools, phosphate transport, and phosphate availability. Annual review of plant physiology 24: 225-252.
-
Kloepper JW, Leong J, Teintze M, Schroth MN (1980) Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 286: 885- 886.
-
Shakeel M, Rais A, Hassan MN, Hafeez FY (2015) Root associated _Bacillus_ sp. improves growth, yield and zinc translocation for basmati rice (_Oryza sativa_) varieties. Front Microbiol 6: 1286
-
Neilands JB (1995) Siderophores: structure and function of microbial iron transport compounds. J Biol Chem 270(45): 26723-26726.
-
Ramesh A, Sharma SK, Sharma MP, Yadav N, Joshi OP (2014) Inoculation of zinc solubilizing _Bacillus_ _aryabhattai_ strains for improved growth, mobilization and biofortification of zinc in soybean and wheat cultivated in Vertisols of central India. Applied Soil Ecology 73: 87-96.
-
Saha M, Sarkar S, Sarkar B, Sharma BK, Bhattacharjee S, et al. (2016) Microbial siderophores and their potential applications: a review. Environ Sci Pollut Res 23(5): 3984-3999.
-
Ahmed E, Holmstrom SJM (2014) Siderophores in environmental research: roles and applications. Microb Biotechnol 7(3): 196-208.
-
Subbiah BV, Asija GL (1956) A rapid procedure for the determination of available nitrogen in soils. Current Science 25: 259-260.
-
Brady NC, Weil RR (1999) The nature and properties of soils. 12th(Edn.), Prentice-Hall, Upper Saddle River, New Jersey, USA.
-
Sharma P, Khurana AS (1997) Effect of single and multi- strain Rhizobium inoculants on biological nitrogen fixation in summer Mungbean, _Vigna radiata_ (L.) Wilczek. Research Development Report 14(2): 8-11.
-
Olsen SR, Cole CV, Wantabe FS, Dean LA (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Circular (United States Department of Agriculture), pp: 93914.
-
Singh D, Chhonkar PK, Dwivedi BS (2005) Soil analysis. In: Manual on soil, plant and water analysis. Westville Publishing House, Delhi, India, pp: 200.
-
Piper CS (1966) Soil and plant analysis: A laboratory manual of methods for the examination of soils and the determination of the inorganic constituents of plants. Soil and Plant Analysis. Asia Edition, Hans Publishers, Bombay, India, 20(3).
-
Ahmed MJ, Iya IR, Dogara MF (2020) Proximate, Mineral and Vitamin Content of Flesh, Blanched and Dried Tomatoes (_Lycopersicum esculentum_). Asian Food Science Journal 18: 11-18.
-
Tandon HLS (2005) Methods of analysis of soil, plants, waters and fertilizers & organic manures. FDCO, New Delhi, India.
-
Nelson LB, Heidel H (1952) Soil analysis methods as used in Iowa state college soil testing laboratory. Iowa Agriculture 57: 1-31.
-
Phillips JM, Hayman DS (1970) Improved procedure for clearing roots and staining parasitic and vesicular- arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society 55(1): 158-161.
-
Giovanetti M, Mosse B (1980) An evaluation of techniques for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytology 84(3): 489-500.
-
Brinkman R (1980) Saline and sodic soils. In: Land reclamation and water management. International Institute for Land Reclamation and Improvement (ILRI), Wageningen, Netherlands, pp: 62-68.
-
Carrow RN, Dunvan RR (2020) Soil salinity monitoring: present and future
-
Ashley MK, Grant M, Grabov A (2006) Plant responses to potassium deficiencies: a role for potassium transport proteins. J Exp Bot 57(2): 425-436.
-
Melo PED (2003) The root system of onion and _Allium fistulosumin_ in the context of organic farming a breeding approach. Ph. D thesis. Wageningen University, Netherlands, pp: 136.
-
Steinmetz KA, Potter JD (1996) Vegetables, fruits and cancer prevention: a review. J Am Diet Assoc 96(10): 1027-1039.
-
Mosse B (1973) Advances in the study of Vesicular arbuscular mycorrhiza. Annual review of phytopathology 11: 171-196.
-
Prasad K (2021g) Glycoprotein Producing AM Fungi lifecycle and Potential Role in Agricultural Plant Lifespan and Global Environmental Changes for Sustainable Green Technology. Journal of Ecology & Natural Resources 5(3): 1-18.
-
Prasad K (2021h) Advantages and Nutritional Importance of Organic Agriculture Produce Food on Human, Soil and Environmental Health in Modern Lifestyle for Sustainable Development. Aditum Journal of Clinical and Biomedical Research 1(2): 1-7.
-
Lidermann RG, Davis EA (2004) Evaluation of commercial inorganic and organic fertilizer effects on arbuscular mycorrhizae formed by Glomus intraradices. Horttechnology 14(2): 196-202.
-
Gerdemann JW (1968) Vesicular-Arbuscular mycorrhiza and plant growth. Annual review of Phytopathology 6: 397-418.
-
Schreiner RP, Mihara KL, McDaniel H, Bethlenfavay GJ (1997) Mycorrhizal fungi improve plant and soil functions and interaction. Plant and Soil 188: 199-207.
-
Johnson JF, Paul LR, Finley RD (2004) Microbial interaction in the mycorrhizaosphere and their significance for sustainable agriculture. FEMS Microbiology Ecology 48(1): 1-13.
-
Joner EJ, Briones R, Leyval C (2000) The metal-binding capacity of arbuscular mycorrhizal mycelium. Plant and Soil 226: 227-234.
-
Bagyaraj DJ (2014) Mycorrhizal fungi. Proc Indian National Science Academy 80(2): 415-428.
-
Akhtar MS, Siddiqui ZA (2010) Effect of AM fungi on the plant growth and root-rot diseases of Chickpea. American-Eurasian Journal of Agriculture and Environmental Science 8(5): 544-549.
-
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress and signal transduction. Annu Rev Plant Biol 55: 373-399.
-
Buchanan BB, Gruissem W, Jones RL (2000) Biochemistry and Molecular Biology of Plants. American Society of Plants Physiologist, Rockville, MD, pp: 1367.
-
Vessey KJ (2003) Plant growth-promoting rhizobacteria as biofertilizers. Plant and Soil 255: 571-586.
-
Trotta A, Varese GC, Gnavi E, Fusconi E, Sampo S, et al. (1996) Interaction between the soil-borne pathogen _Phytophthora_ _parasitica_ _var._ _parasitica_ and the arbuscular mycorrhizal fungus Glomus mosseae in tomato plants. Plant and Soil 185: 199-209.
-
Cordier C, Gianinazzi S, Pearson VG, Trouvelot A (1996) Arbuscular mycorrhiza Technology applied to micro propagated Prunus avium and to protection against Phytophthora cinnamomi. Agronomies 16(10): 679-688.
-
Siddiqui S, Alrumman SA, Meghavanshi MK, Chaudhary KK, Prasad K, et al. (2015) Role of Soil Amendment with Micronutrients in Suppression of Certain Soil-Borne Plant Fungal Diseases: A Review. In: Meghavanshi MK, et al. (Eds.), Organic Amendments and Soil Suppressiveness in Plant Disease management, Springer Soil Biology 46. Springer International Publishing, Switzerland, pp: 363- 380.
-
Turnau K, Haselwandter K (2002) Arbuscular mycorrhizal fungi, an essential component of soil microflora in ecosystem restoration. In: Gianinazzi S, et al. (Eds.), Mycorrhizal Technology in Agriculture. Birkhauser, Basel, pp: 137-149.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania