Vaccine-Induced Immune Thrombotic Thrombocytopenia: Where does The PF4 Fit?
Vaccine-induced immune thrombotic thrombocytopenia (VITT) is a severe and usually fatal syndrome with an aggressive thrombotic process in unusual sites (notably, sinus veins of the brain and/or splanchnic veins) that accompanies profound thrombocytopenia. If not well managed, VITT may progress to a more severe systemic disease such as disseminated intravascular coagulation. VITT was associated with two adenoviral vector vaccines, mainly ChAdOx1 nCoV-19 (AstraZeneca) and Ad26.COV2.S (Janssen/ Johnson & Johnson) was first reported in late February 2021 and mid-April 2021, respectively. This thrombotic phenomenon closely resembled that of autoimmune heparin-induced thrombocytopenia. Yet, in 2021, the MHRA (Medicines & Healthcare products Regulatory Agency) and the European Medicines Agency (EMA) claim no association between thrombosis and vaccines. Not long after, three scientific societies from Norway, Germany, and the UK reported in the press and social media the detection of thrombocytopenia with cerebral venous sinus thrombosis (CVST) and anti-platelet factor 4 (anti-PF4) antibodies in patients succeeding adenovirus-based vaccination. The immune response against PF4; (also known as CXCL4) in VITT is probably triggered by the proinflammatory milieu; compounds such as human cell-line proteins, non-assembled adenoviral proteins, and potentially EDTA (edetic acid) could be contributing to the prothrombotic state. This review will address functional aspects of PF4 in the thromboinflammatory phenomena, its role in the current anti-SARS-CoV-2 adenoviral-based vaccines, and evidence for its role in triggering VITT. Physicians as well as the public need to be aware of this new disease to quickly provide accurate diagnosis and timely treatment.
Introduction
As of June 7, 2022, 11.66 billion doses of vaccines for SARS-CoV-2 have been administered in more than 180 countries [1]. The current pandemic scenario since 2020 has empowered many studies related to immunizations never seen before in vaccination history. The worldwide effort to mitigate the spread and infection of COVID-19 has been unprecedented in terms of the speed and the efficiency of the vaccines. View-hub, an interactive map-based platform for visualizing vaccine use and impact data, described 300 studies in 40 countries, 105 with an adenoviral-based platform [2]. Fears about thrombosis have generated confusion between the prothrombotic processes due to COVID-19 and ongoing vaccination in an emerging pandemic. COVID-19 has also been called the first thrombotic viral fever [3], in an opposite analogy to the so-called hemorrhagic viral fevers, such as Dengue and Ebola, among others [4]. Further expanding the terminology confusion, VITT has gained other synonyms such as “vaccine-induced prothrombotic immune thrombocytopenia”, or VIPIT, “Vaccine-associated thrombotic thrombocytopenia”, or VATT, or even “Thrombosis with Thrombocytopenia Syndrome”, or TTS [5]. To date, no other adenoviral SARS-CoV-2 vaccine, such as Ad5-based COVID-19 (CanSino Biologics) and Gam-COVID-Vac/Sputnik V (Gamaleya Institute), has been related to reported cases of VITT. Michalik S, et al. [6] noticed more significant amounts of impurities containing host cell proteins, adenoviral proteins and EDTA (Edetic Acid), among other contaminants, in ChAdOx1 nCoV-19 (AZ) than in Ad26.COV2.S (Jansen) adenoviral-based vaccines. Otherwise, according to these authors, the ChAdOx1 nCoV-19 showed a significant increase in capillary leakage and subsequently a greater incidence of VITT when compared with Ad26.COV2.S.
Furthermore, of these well-studied adenoviral-based vaccines, few reported cases of VITT are also related to mRNA platform vaccines [7, 8]. Covid19 is just over 2 years old, and its vaccines are much more recent. Therefore, the most solid studies are still insufficient, especially to identify the adverse effects of so many different vaccines, many not yet implemented. Perhaps we will have better evidence and more factual data to clarify many thrombosis phenomena, especially between other vaccine platforms. It is important to emphasize that thrombotic syndrome with thrombocytopenia occurs in other diseases such as thrombotic thrombocytopenic purpura, antiphospholipid syndrome or even heparin- induced autoimmune thrombocytopenia and that clinical, laboratory and epidemiological aspects are crucial to aid in the diagnosis of VITT. Besides, thrombocytopenia could be a pseudothrombocytopenia, thus the importance of looking at a blood smear under a microscope.
Epidemiology
Nina Schultz, et al. [9] reported 5 cases of VITT, 7 to 10 days after vaccination against COVID-19 in Norway. Three people had cerebral venous sinus thrombosis with intracranial haemorrhage, with a fatal outcome. Therefore, among 132,686 people vaccinated with AstraZeneca, there was an incidence of 1 case of VITT for every 26,000 vaccines. In the UK, the MHRA (Medicines & Healthcare products Regulatory Agency), the first regulatory body in Europe to authorize a SARS-CoV-2 vaccine, identified 367 cases of VITT after 24.7 million first doses of the vaccine. Further, 44 patients after the second dose, giving the ratio of one case of VITT to 67.302 and 518,181 vaccines, respectively [10]. In Germany, both vaccines produced an incidence of 0,56 suspected cases per 100,000 vaccine doses for Ad26.COV2.S and 1.49 suspected cases per 100,000 vaccine doses for ChAdOx1 nCoV-19 [11]. As clearly demonstrated by these numbers, VITT is a rare event and is less frequent than thrombosis resulting from severe COVID-19 despite thromboprophylaxis, especially considering intensive care patients [12]. In the USA, 12 cases of VITT were reported by See, et al. [13] after 7 million doses, leading to a rate of 1 VITT to every 583,000 vaccinations. Both adenoviral-based vaccines mentioned above are produced in human cell lines, embryonic kidney cells, a HEK293 derivate for AstraZeneca, and human embryonic retinal cells, PER.C6 TetR cells, for Jansen vaccine. When considering the incidence of thrombocytopenia and thrombosis as complications in the general population compared to VITT, the annual incidence of isolated thrombocytopenia, such as immune thrombocytopenia (ITP), or isolated cerebral venous sinus thrombosis (CVST) is higher than that reported for VITT. However, when adjusted for the two weeks characteristic of adenoviral-based vaccine-induced events, the corresponding rates are lower than those of VITT [14]. In summary, isolated CSVT is reported in 13 to 20 cases per million (0.5 -0.77 cases per million in any two weeks [15], and ITP in the general population occurs in 16 to 39 cases per million (0.61 -1.5 cases per million in any two weeks) [16].
The Weight of PF4 in Coagulation and Immunopathology
Platelet factor 4 (PF4), a small chemokine (CXCL4), is an abundant cationic, 7.8 kDa protein with 70-amino- acid secreted by activated platelets through it’s α-granules (composed of various chemokines, cytokines, membrane proteins, proteases, proinflammatory and anti-inflammatory mediators). PF4 interact with many biologic structures forming tetramers at ionic strength and making composite arrangements with anionic transporters. In addition, it has a solid affinity to endothelium which is plenty with glycosaminoglycan (GAG)-rich glycocalyx. This way, by binding to heparans, chondroitin sulfate/dermatan sulfate (CSGAGs), keratan sulfate, and hyaluronic acid, in a certain way, can inhibit antithrombin activity (the more potent thrombin inhibitor), thus stimulating coagulation as a procoagulant inducer. Damage to the endothelium enhances an imbalance in the blood coagulation system, primarily by increasing thrombin generation due to losses of natural anticoagulant forces and cytoprotective mechanisms from endothelial cells (Figure 1). The integrity of the endothelial GAGs is fundamental because it enhances biological anticoagulant activity like the antithrombin, tissue factor pathway inhibitor (TFPI) and thrombomodulin-thrombin complex that activates Protein C (APC), which can interact with its ligand and promote cell protection [3]. The PF4 released from the platelet α-granules interacts with thrombomodulin and the g-carboxyglutamic acid (Gla) domain, which anchor the natural anticoagulant protein C (and the vitamin K dependent factors) on membranes, enhancing its activation to an active structure called APC (activated protein C) together with endothelial protein C receptor (EPCR) [17]. Still, once generated, PF4 inhibited both (PS)-protein S-dependent factor Va by order of 5-fold, demonstrating that this chemokine impairs protein S (PS) action, which is a cofactor effect that booster the anticoagulant effect of APC [17, 18] (Figure 1).
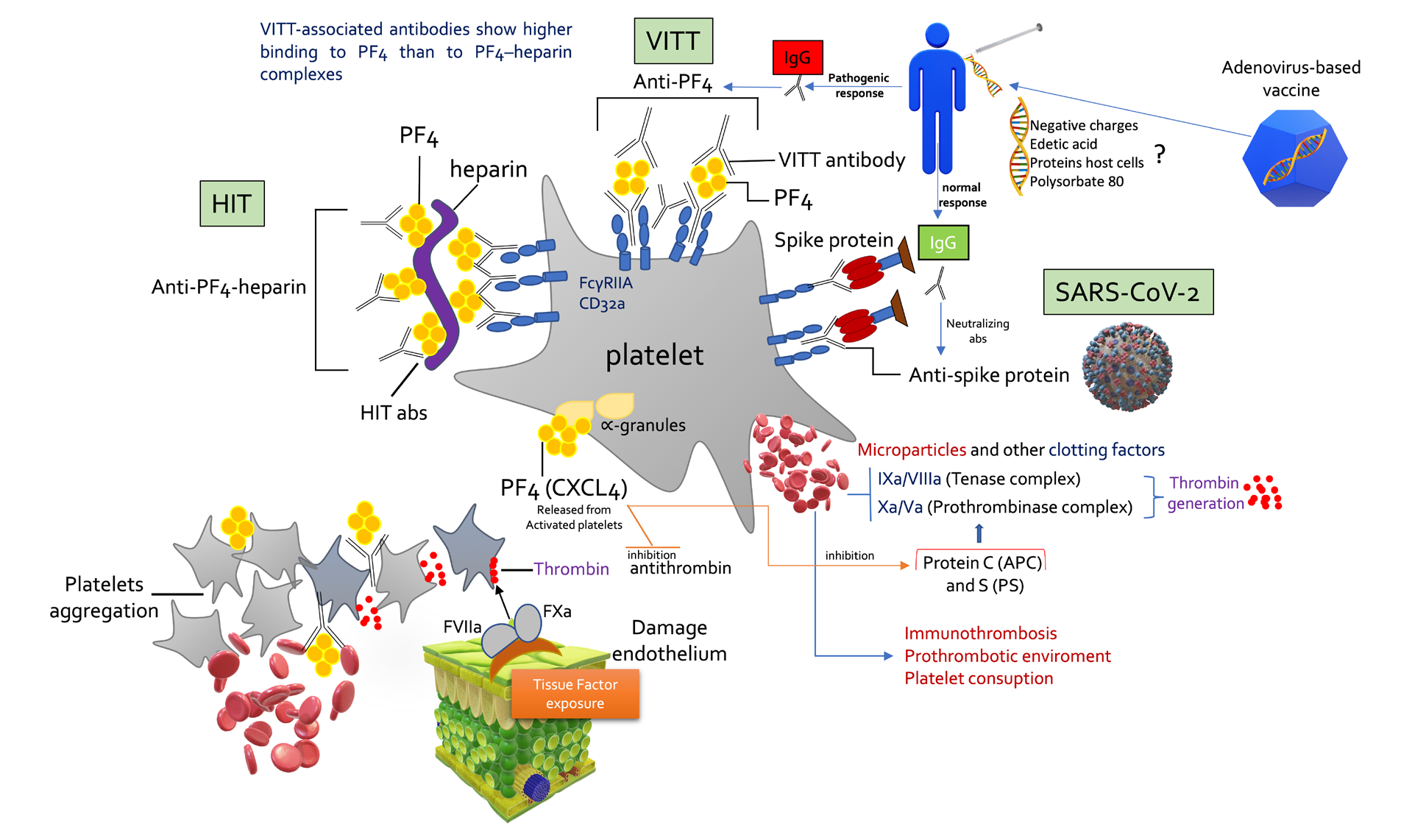
VITT antibodies have sufficient binding strength (avidity) to form immune complexes with PF4 and to crosslink FcγRIIa (CD32a) receptors on platelets and, consequently, activate platelets, producing microparticles rich in procoagulants such as tenase and prothrombinase complexes. These complexes located in the bulge of the micro particles activate thrombin generation in the order of magnitude 10 (6). The figure describes platelet activation promoted by HIT, VITT and COVID-19 in a didactic way. The anionic elements contained among vaccine impurities could activate the pathogenic response for the formation of anti-PF4 antibodies. This figure overwhelmed the presence of antigen-presenting cells that amplify thrombotic and inflammatory phenomena. VITT – Vaccine-induced immune thrombotic thrombocytopenia. HIT- Heparin-induced thrombocytopenia. APC- Activated Protein C.
Proteolysis of the cofactor (FVa) by APC is enhanced > 20-fold in the presence of PS, thus causing a blunting of the mechanisms of interruption of thrombin generation by the prothrombinase (Factor IXa/VIIIa) and tenase (Factor Xa/Va) complexes that might be dismantled by the action of APC and PS [3]. In other words, PF4 impairs APC anticoagulant function by interacting with its GLA domain [17]. Furthermore, PF4 does not directly alter the cytoprotecting effects of APC/EPCR, although it impairs the anticoagulant biological action of APC/PS. Additionally, by neutralizing the side chains of negatively charged GAGs on the surface of platelets and endothelial cells, PF4 facilitates platelet aggregation to form a thrombus, promoting more hypercoagulation, which is necessary to mitigate bleeding, i.e., such as trauma in which elevated levels of PF4 are also found [19]. Yet, PF4 has other functions beyond its foremost purpose, which would be to promote blood clotting. As chemokine, PF4 is also involved in innate and adaptive immunity when platelets are activated in response to infections, as it similarly functions as a chemoattractant for neutrophils, monocytes, and fibroblasts, which gives PF4 a paramount role in the thromboinflammation phenomena. The PF4 induces granulocyte recruitment and facilitates the release of myeloperoxidase and lysozyme, improving the phagocytosis of microorganisms through the opsonization of their surfaces and thus facilitating the binding of antibodies. Moreover, platelet activation induced P-selectin (CD62P) expression that, together with PF4, interacts with neutrophils and monocytes, leading to the formation of neutrophil extracellular traps (NETs). NETs, on the one hand, limit the infectious process by trapping pathogens, but on the other, it exacerbates the inflammatory process, further aggravating endothelial dysfunction [20]. Additionally, P-selectin activates monocytes to produce tissue factor (TF), which is the primary activator of the extrinsic pathway of coagulation. In COVID-19 patients, it has already been shown that there is a high platelet hyperreactivity linked to the increased expression of P-selectin [21].
Interestingly, PF4 can stimulate antigen-presenting cells (APCs) to induce the proliferation of lymphocytes and cytotoxic activity of NK cells and, in this way, increase the clearance of viruses in pulmonary infections [22]. Hottz, et al. [23] found that activated platelets in severe COVID-19 interacted with monocytes (platelet-monocytes) observed in plasma from healthy volunteers, which were strongly associated with tissue factor expression. This finding could be related to high mortality in patients with elevated thrombotic risk affected by severe COVID-19. Studies that reveal the presence of pathogenic PF4, involved in the thrombotic events of the different levels of manifestations of COVID-19, have not yet been demonstrated. In this scenario, thrombin also induces the release of P-selectin, which help bind to P-selectin glycoprotein ligand-1 (PSGL-1) on neutrophils, monocytes, and lymphocytes, mediating their tethering and rolling, giving more injury to the endothelium.
However, it is crucial to note that PF4 suffers structural conformational changes after binding to bacteria or any cell surface [24], thus transforming it into a pathogenic structure that induces antibody formation (Figure 1). This point is fundamental because anti-PF4 antibodies often bind to PF4 on platelets and endothelial cell surfaces, provoking platelet and leukocyte activation and chronic inflammation through FcγIIa/IIIa receptors responsible for endocytosis of immunoglobulin G (IgG) coated pathogens [25]. These interactions between PF4 and polyanions usually trigger the formation of antibodies of the IgG4 class to induce prothrombotic processes, such as the well- known heparin-induced thrombocytopenia (HIT) and, more recently, with adenoviral vectored vaccine-induced immune thrombocytopenia with a severe thrombotic syndrome (VITT). This new syndrome, resembling autoimmune heparin-induced thrombocytopenia, does not require the presence of heparins to follow. VITT, similar to HIT, also happen in unusual locations (brain, lung, hepatic vein, splanchnic vein) and is more frequent in females and young people [26]. Given these assumptions, the rationale for anticoagulation would be to mitigate the generation of thrombin, and high doses of immunoglobulins with steroid adjuvants in treating VITT would block FcγIIa/IIIa receptors mediated by cellular activation of cells involved in the immune system. This plethora of reactions ensues because the activated B cells, when secreting immunoglobulins specific to PF4, also stimulate a range of cells leading to NETosis and a massive explosion of thrombin increase, increasing the thromboinflammation [27]. Nonetheless, not all anti-PF4 building is pathogenic, i.e., individuals with chronic periodontal disease had elevated titers of anti-PF4 but no increased risk of thrombotic complications [28].
Platelets and Viral Infections
Platelets play an essential role in viral infections, especially through the presence of Toll receptors on their surface. TLRs activate innate immunity by binding pathogen-associated molecular patterns (PAPMs). Some (TLR3, TLR7/8 and TLR9) are located in endosomes and detect RNA or DNA-based molecular structures associated with pathogens [29]. Studies have shown that an intrinsic defence against influenza is mediated by interactions between platelets and neutrophils in a cross-communication that regulates host immunity and the complement system involving TLR7, among others [30]. Increased expression of TLRs and TLR-mediated platelet activation during COVID-19 may increase vascular thrombosis. The SARS-COV-2 viral ss- RNA is detected by the proteins TLR-3, TLR-7, and TLR-8. Other studies [31, 32] have found platelets’ internalization and clearance of ssRNA viruses such as influenza and HIV. For example, PF4 in a monomeric state bind to the HIV-1 envelope protein gp120 and inhibits the attachment of HIV-1 to the cell surface, thus reducing the entry of HIV-1 into CD4+ T cells [33]. Thus, HIV-1 infected with low serum monomeric PF4 levels tend to have worse clinical outcomes. As the concentration of PF4 increases, it tends to form tetramers, or higher-order forms, that worsen HIV-1 infection in vitro. Soluble glycosaminoglycans (GAGs) prevent the formation of tetrameric PF4. In other words, the more endothelial integrity, the less tetrameric state form of PF4.
After viral infection, platelets are activated through TLR7, which changes their phenotype, favouring the formation of platelet-neutrophil aggregates. After lysosomal degradation of the internalized viral particles, the ssRNA genome binds to endosomal TLR-7. The virus, via ssRNA, can mediate platelet activation through TLR-7, leading to degranulation and consequent secretion of PF4. This mechanism likely participates in antiviral immunity, as TLR- 7-depleted mice had increased mortality [34, 35]. Platelets always participate in thrombosis phenomena in different shades, so they are confused between those promoted by severe COVID-19 and those triggered by mass vaccination, especially vaccines based on the adenoviral platform (Figure 1). PF4 stands out as it is secreted by alpha granules after platelets are activated. However, in severe SARS-CoV-2, pathogenic PF4 antibodies’ contribution to the amplification and perpetuation of thrombotic phenomena has been poorly studied. Platelet overexpression of P-selectin on platelets in critically ill patients with SARS-CoV-2 suggests a central role of platelet activation as part of the pathogenic mechanism of COVID-19, which aids in the formation of micro thrombosis [21].
Lastly, the seminal study by Biembengut IV, et al. [36] investigated potential inhibitors of SARS-CoV-2 main protease (Mpro) using crystallographic structures and observed the potential for using antiviral drugs that act on this essential protease. These studies also identified anticoagulant compounds capable of inhibiting this protease, such as Edoxaban, Betrixaban and Apixaban, all Factor Xa inhibitors, with Apixaban showing high binding affinity with the viral Mpro [3, 36].
Occurrence of VITT Between Different Vaccines
Based on this actual physiopathology, although not fully understood, the current pillars of the way to approach VITT are centred on blocking the immunologic system and using non-heparin anticoagulation. The compounds embedded in viral platform vaccines, which carry a negative charge, such as EDTA (edetic acid), nucleic acids (DNA, RNA), polysorbate 80, proteins derived from host cells, glycosaminoglycans, sulfated compounds are important candidates that bind and transform this cationic chemokine CXCL4 (PF4). And among viral platform vaccines, VITT appears to be three times more frequent after the ChAdOx1 vaccine than Ad26.
COV2.S vaccine [37]. Analyses performed by Michalik et al. [6] showed significant differences between ChAdOx1 and Ad26.COV2.S vaccines regarding the proportion of host- cell proteins (54% in the ChAdOx1 nCoV19 vaccine). Only low levels of host-cell protein were found in Ad26.COV2.S (1.5%) indicating that different purification approaches and efficiencies methods are applied to their manufacture. These disparities between vaccine impurities may influence their ability to interact with PF4 to form large complexes, mainly after adding PF4 to the media containing virions of the ChAdOx1 nCoV19 vaccine. Again, data obtained by cryo- electron microscopy and in silico modelling of the ChAdOx1 nCoV-19 vaccine established a likely electrostatic interaction of cationic PF4 and anionic adenovirus hexon polypeptide [38].
Importance of Pathogenic Anti-PF4 Antibodies in Immune Syndromes
HIT [39] and VITT [40] are temporally prothrombotic immunological syndromes (Figure 1). Antibodies formation against pathogenic PF4 is enhanced by antigen creation in an inflammatory environment. Importantly, not all PF4 antibodies are pathogenic and lead to HIT or VITT, and current clinical tests are not practical to determine whether the PF4 abs is pathogenic or non-pathogenic. Zheng Cai et al. [19, 41] proposed that covalent bonding between pentasaccharide units in longer heparins (such as UFH) markedly increases the stability of the PF4 tetrameric complex. Consequently, making the molecule more “stable”, and thus, it becomes more antigenic, increasing the avidity for antibodies of the IgG class. The PF4 exists in a tenuous dynamic equilibrium between monomers and tetramers. By stabilizing a large number of PF4 protein monomers, there is a tendency to promote the dissociation of PF4 tetramers. This prevents the subsequent development of immunopathogenic PF4 complexes, as they rely on tetramers [19]. Plasma PF4 levels are related to the extent and duration of platelet activation and the turnover of this chemokine. All conditions that disturb endothelial health (diabetes, arteriosclerosis, sepsis, kidney disease, cardiovascular disease, and trauma) or even invasive procedures such as ECMO (extracorporeal membrane oxygenation) and CRRT (continuous renal replacement therapy) commonly used in severe COVID-19 would generate high levels of PF4 and consequently requires heparin administration. In classical HIT syndrome, cardiac surgery with cardiopulmonary bypass and large volumes of heparins (the highest negative charge density of all known natural molecules due to its heavily sulfated glycosaminoglycans) could cause a greater incidence of HIT than in clinical contexts with less activated platelets. Systemic vascular hyperpermeability might facilitate direct contact of the immune system with vaccine impurities due to its extravasation to the interstitial space and, consequently lymphatic system. In such a way, the endogenous natural IgG binds proteins of degrading cells to form immune complexes [42].
According to the study by Universitätsmedizin Greifswald, in 14 of the 15 patients with a follow-up of more than 12 weeks (93%; 95% CI, 70 to 100), the platelet- activation assay became negative within a median of 12 weeks (95% CI, 10 to 13). Also, full seroreversion to a negative ELISA result (i.e., an optical density of <0.5) was seen in only 3 patients, and 1 patient showed PF4-dependent platelet- activating antibodies and ELISA optical density values of greater than 3.0 for more than 12 weeks in association with sporadic thrombocytopenia [40]. Therefore, it is crucial to measure these antibodies by the ELISA method, given the risk of developing VITT in case of incomplete vaccination and the schedule for a second dose. Furthermore, the question remains whether patients using anticoagulants as part of the treatment of confirmed VITT should be monitored by the dosage of these anti-PF4 antibodies in an attempt to seek OD less than 0.5 by the ELISA method [43].
Are HIT and VITT Identical Syndromes?
VITT is similar to other PF4-mediated adverse drug effects, such as heparin-induced thrombocytopenia (HIT) and autoimmune HIT [44, 45]. In HIT, polyanions such as heparins form complexes with cationic chemokine CXCL4 (PF4), inducing a conformational change capable of producing HIT antibodies (Figure 1). This immune response in both syndromes is amplified by endhoteliitis and systemic inflammation. Studies have shown that one or more constituents of the ChAdOx1 nCoV-19 vaccine interact with PF4, making complexes containing PF4 and the adenovirus hexon protein, a negative charge molecule similar to heparins or GAGs which may induce thrombotic thrombocytopenic syndrome (HIT). Individual platelet phenotypic characteristics can affect assembly, size, and physicochemical characteristics, in addition to the properties of antigen-antibody complexes (anti-PF4–PF4). Then, their participation with Fc gamma receptors (FcγRIIa/IIIa) promotes a significant increase in the procoagulant state with more prominent thrombin generation and fibrin deposition by excess activation of the platelets, granulocytes, and endothelium. This process creates a highly self-perpetuating prothrombotic environment in the vessel lumen and cell surfaces. The origin and development of autoantibodies in VITT in the context of immunization remains an open question. However, much of what is known about the source of HIT antibodies may apply to VITT. Experimental studies have shown that anti-PF4 antibodies from patients with VITT – distinctive patients with HIT - without exposure to heparin, curiously are circumscribed to the same heparin- binding site [46].
Detection and Utility of VITT Antibodies
The typical laboratory feature described in the published VITT case series is composed of very high Dimers-D (4x the upper limit of normal), the presence of thrombocytopenia and low fibrinogen levels, which characterize a coagulopathy, especially in the most severe cases. These changes are also frequent in HIT. There are other syndromes with a similar prothrombotic profile, such as acquired thrombophilia such as antiphospholipid antibody syndrome, paroxysmal nocturnal hemoglobinuria, developed von Willebrand syndrome with ADAMTS-13 (A Disintegrin and Metalloproteinase with ThromboSpondin type 1 motif, number 13)deficiency, myeloproliferative neoplasms, thrombotic thrombocytopenic purpura and a distinct prothrombotic phenotype of disseminated intravascular coagulation [47], parameters like anti-PF4 antibodies with high levels of D-dimer (> 4 times ULN – Upper Limit Normal) usually are standard (except in DIC and TTP). Anti-PF4 antibodies are detected by the ELISA method in most patients with HIT and VITT. However, important underline that rapid tests such as immunoassays (PaGIA), lateral flow assay (LFA), and IgG-chemiluminescence immunoassay (CLIA) can yield false-positive results, contrary to those using enzyme-linked immunosorbent assays. Many reports have demonstrated a strong correlation between anti-PF4 seropositivity and functional platelet activation tests. However, recent case series have shown that individuals are vaccinated with Ad26. COV2.S had low positivity rates in these tests. For example, there was only one positive HIPA case out of 9 negative ones [13], making these tests to confirm VITT questionably.
Conclusion
VITT is a recent syndrome, a descendant of the COVID-19 syndemic [48]. A rare adverse effect of a necessary mass vaccination that fortunately has been mitigating millions of global deaths. However, thromboses have always concerned the survival of patients with comorbidities or severe infections. One in four people worldwide dies from diseases related to thrombosis, or more specifically, venous thromboembolism (VTE). Whether due to cardiovascular diseases, including stroke and venous thrombosis, such as those resulting from infectious processes, given that COVID-19 could be considered the first viral thrombotic fever [3]. VITT came as an undesirable disease arising from anti-SARS-CoV-2 vaccines, especially those that carry the viral Spike protein from SARS-CoV-2 through adenoviral vectors (Figure 1). The genesis of immunopathogenic PF4 has been studied for decades, identifying another very close syndrome related to VITT under immunological perspectives, called heparin-induced thrombocytopenia (HIT), and more recently discovered with autoimmune involvement, that is, without the presence of heparins [44]. The biggest challenge is to develop strategies to identify which patients are highest at risk for the development of vaccine-related thrombosis. In counterpart, discover better and more efficient techniques to perform vaccine purification, especially those from adenoviral vector platforms, particularly in removing these anionic molecules capable of triggering the formation of pathogenic PF4. This review intends to generate reflections and evidence about the problem of adverse events related to the COVID-19 vaccine’s severe effects. Thus, soon, we hope that the teams in the biomedical area will develop more studies and hypotheses that can mitigate this issue.
And finally, perhaps the development of laboratory techniques capable of identifying the oligomeric forms from the tetrameric PF4 states, which the pathogenic ones are. The basis that distinguishes pathogenic from non-pathogenic antibodies is still unknown, which leads to overdiagnosis and overtreatment.
Acknowledgements
The authors acknowledge Thiago Carmo Costa for his valuable help managing the English edition and correction. AMD-C and HCCF-N are fellows from CNPq and CNE/FAPERJ. HCCFN is a grantee from CNPq # 401700/2020-8.
Authors Contributions
Rubens C. Costa-Filho, Hugo Caire de Castro Faria-Neto and Alda Maria Da-Cruz participated in outlining the article. All Authors revised the manuscript critically for important intellectual content and construct. Marcela Azevedo conducted supervision and reviewed the literature as well as figure design.
References
-
COVID-19 Dashboard, Center for Systems Science and Engineering (CSSE), Johns Hopkins University (JHU).
-
COVID-19 Data, View-Hub 2022 [This section contains information on vaccine effectiveness studies that have been reported in preprint and published literature and reports.].
-
Costa-Filho RC, Castro-Faria HCN, Mengel J, Pelajo- Machado M, Martins MA, et al. (2021) Should COVID-19 be branded to viral thrombotic fever? Mem Inst Oswaldo Cruz 116: e200552.
-
Schnittler HJ, Feldmann H (2003) Viral hemorrhagic fever--a vascular disease? Thromb Haemost 89(6): 967- 972.
-
Warkentin TE, Pai M (2021) VIPIT versus VITT: what’s in a name? Thrombosis Research 207(11): 158-160.
-
Michalik S, Siegerist F, Palankar R, Franzke K, Schindler M, et al. (2022) Comparative analysis of ChAdOx1 nCoV-19 and Ad26.COV2.S SARS-CoV-2 vector vaccines. Haematologica 107(4): 947-957.
-
Sangli S, Virani A, Cheronis N, Vannatter B, Minich C, et al. (2021) Thrombosis With Thrombocytopenia After the Messenger RNA-1273 Vaccine. Ann Intern Med 174(10): 1480-1482.
-
Pishko AM, Cuker A (2021) Thrombosis after Vaccination with Messenger RNA-1273: Is This Vaccine-Induced Thrombosis and Thrombocytopenia or Thrombosis with Thrombocytopenia Syndrome? Ann Intern Med 174(10): 1468-1469.
-
Schultz NH, Sorvoll IH, Michelsen AE, Munthe LA, Johansen FL, et al. (2021) Thrombosis and Thrombocytopenia after ChAdOx1 nCoV-19 Vaccination. N Engl J Med 384(22): 2124-2130.
-
Klok FA, Pai M, Huisman MV, Makris M (2022) Vaccine- induced immune thrombotic thrombocytopenia. The Lancet Haematology 9(1): e73-e80.
-
Ehrlich PI (2021) Sicherheitsbericht. Federal Institute for Vaccines and Biomedicines.
-
Yamada S, Asakura H (2022) Coagulopathy and Fibrinolytic Pathophysiology in COVID-19 and SARS- CoV-2 Vaccination. Int J Mol Sci 23(6): 3338.
-
See I, Su JR, Lale A, Woo EJ, Guh AY, et al. (2021) US Case Reports of Cerebral Venous Sinus Thrombosis With Thrombocytopenia After Ad26.COV2.S Vaccination, March 2 to April 21, 2021. JAMA 325(24): 2448-2456.
-
Arepally GM, Ortel TL (2021) Vaccine-induced immune thrombotic thrombocytopenia: what we know and do not know. Blood 138(4): 293-298.
-
Otite FO, Patel S, Sharma R, Khandwala P, Desai D, et al. (2020) Trends in incidence and epidemiologic characteristics of cerebral venous thrombosis in the United States. Neurology 95(16): e2200-e2213.
-
Moulis G, Palmaro A, Montastruc JL, Godeau B, Mestre ML, et al. (2014) Epidemiology of incident immune thrombocytopenia: a nationwide population-based study in France. Blood 124(22): 3308-3315.
-
Preston RJS, Tran S, Johnson JA, Ni Ainle FN, Harmon S, et al. (2009) Platelet factor 4 impairs the anticoagulant activity of activated protein C. J Biol Chem 284(9): 5869- 5875.
-
Kohli S, Shahzad K, Jouppila A, Holthofer H, Isermann B, et al. (2022) Thrombosis and Inflammation-A Dynamic Interplay and the Role of Glycosaminoglycans and Activated Protein C. Front Cardiovasc Med 9: 866751.
-
Cai Z, Greene MI, Zhu Z, Zhang H (2020) Structural Features and PF4 Functions that Occur in Heparin- Induced Thrombocytopenia (HIT) Complicated by COVID-19. Antibodies (Basel) 9(4): 52.
-
Guo L, Rondina MT (2019) The Era of Thromboinflammation: Platelets Are Dynamic Sensors and Effector Cells During Infectious Diseases. Front Immunol 10: 2204.
-
Manne BK, Denorme F, Middleton EA, Portier I, Rowley JW, et al. (2020) Platelet gene expression and function in patients with COVID-19. Blood 136(11): 1317-1329.
-
Guo L, Feng K, Wang YC, Mei JJ, Ning RT, et al. (2017) Critical role of CXCL4 in the lung pathogenesis of influenza (H1N1) respiratory infection. Mucosal Immunol 10(6): 1529-1541.
-
Hottz ED, Quintanilha IGA, Palhinha L, Teixeira L, Barreto EA, et al. (2020) Platelet activation and platelet- monocyte aggregate formation trigger tissue factor expression in patients with severe COVID-19. Blood 136(11): 1330-1341.
-
Krauel K, Potschke C, Weber C, Kessler W, Furll B, et al. (2011) Platelet factor 4 binds to bacteria, [corrected] inducing antibodies cross-reacting with the major antigen in heparin-induced thrombocytopenia. Blood 117(4): 1370-1378.
-
Worth RG, Chien CD, Chien P, Reilly MP, McKenzie SE, et al. (2006) Platelet FcgammaRIIA binds and internalizes IgG-containing complexes. Exp Hematol 34(11): 1490- 1495.
-
Di Micco P, Camporese G, Cardillo G, Lodigiani C, Carannante N, et al. (2021) Pathophysiology of vaccine- induced prothrombotic immune thrombocytopenia (Vipit) and vaccine-induced thrombocytopenic thrombosis (vitt) and their diagnostic approach in emergency. Medicina (Kaunas) 57(10): 997.
-
Costa-Filho RC, Bozza FA (2017) Platelets: an outlook from biology through evidence-based achievements in critical care. Ann Transl Med 5(22): 449.
-
Greinacher A, Holtfreter B, Krauel K, Gatke D, Weber C, et al. (2011) Association of natural anti-platelet factor 4/heparin antibodies with periodontal disease. Blood 118(5): 1395-1401.
-
Girkin JLN, Maltby S, Bartlett NW (2022) Toll-like receptor-agonist-based therapies for respiratory viral diseases: thinking outside the cell. Eur Respir Rev 31(164): 210274.
-
Koupenova M, Corkrey HA, Vitseva O, Manni G, Pang CJ, et al. (2019) The role of platelets in mediating a response to human influenza infection. Nat Commun 10(1): 1780.
-
Koenen RR (2016) The prowess of platelets in immunity and inflammation. Thromb Haemost 116(4): 605-612.
-
Shiraki R, Inoue N, Kawasaki S, Takei A, Kadotani M, et al. (2004) Expression of Toll-like receptors on human platelets. Thromb Res 113(6): 379-385.
-
Auerbach DJ, Lin Y, Miao H, Cimbro R, Difiore MJ, et al. (2012) Identification of the platelet-derived chemokine CXCL4/PF-4 as a broad-spectrum HIV-1 inhibitor. Proc Natl Acad Sci USA 109(24): 9569-9574.
-
Fard MB, Fard SB, Ramazi S, Atashi A, Eslamifar Z (2021) Thrombosis in COVID-19 infection: Role of platelet activation-mediated immunity. Thromb J 19(1): 59.
-
Banerjee M, Huang Y, Joshi S, Popa GJ, Mendenhall MD, et al. (2020) Platelets Endocytose Viral Particles and Are Activated via TLR (Toll-Like Receptor) Signaling. Arterioscler Thromb Vasc Biol 40(7): 1635-1650.
-
Biembengut IV, Souza TDACBD (2020) Coagulation modifiers targeting SARS-CoV-2 main protease Mpro for COVID-19 treatment: an in silico approach. Mem Inst Oswaldo Cruz 115: e200179.
-
See I, Lale A, Marquez P, Streiff MB, Wheeler AP, et al. (2022) Case Series of Thrombosis With Thrombocytopenia Syndrome After COVID-19 Vaccination-United States, December 2020 to August 2021. Ann Intern Med 175(4): 513-522.
-
Baker AT, Boyd RJ, Sarkar D, Crespo AT, Chan CK, et al. (2021) ChAdOx1 interacts with CAR and PF4 with implications for thrombosis with thrombocytopenia syndrome. Sci Adv 7(49): eabl8213.
-
Marchetti M, Zermatten MG, Calderara DB, Aliotta A, Alberio L (2021) Heparin-Induced Thrombocytopenia: A Review of New Concepts in Pathogenesis, Diagnosis, and Management. J Clin Med 10(4): 683.
-
Schonborn L, Thiele T, Kaderali L, Greinacher A (2021) Decline in Pathogenic Antibodies over Time in VITT. N Engl J Med 385(19): 1815-1816.
-
Cai Z, Yarovoi SV, Zhu Z, Rauova L, Hayes V, et al. (2015) Atomic description of the immune complex involved in heparin-induced thrombocytopenia. Nat Commun 6: 8277.
-
Pfueller SL, Logan D, Tran TT, Bilston RA (1990) Naturally occurring human IgG antibodies to intracellular and cytoskeletal components of human platelets. Clin Exp Immunol 79(3): 367-373.
-
Costa-Filho RC, Monteiro de Souza ATA, Tantos Nunes G, Melo Amorim Filho L, Saddy F, et al. (2022) Vaccine- Induced Immune Thrombotic Thrombocytopenia (VITT) by A Third Dose of Chadox1 Ncov-19 (Astrazeneca) After BNT162b2 (Pfizer–BioNTech). Virology & Immunology Journal 6(2): 1-5.
-
Greinacher A, Selleng K, Warkentin TE (2017) Autoimmune heparin-induced thrombocytopenia. J Thromb Haemost 15(11): 2099-2114.
-
Greinacher A, Thiele T, Warkentin TE, Weisser K, Kyrle PA, et al. (2021) Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. N Engl J Med 384(22): 2092-2101.
-
Huynh A, Kelton JG, Arnold DM, Daka M, Nazy I (2021) Antibody epitopes in vaccine-induced immune thrombotic thrombocytopaenia. Nature 596(7873): 565-569.
-
Popescu NI, Lupu C, Lupu F (2022) Disseminated intravascular coagulation and its immune mechanisms. Blood 139(13): 1973-1986.
-
Horton R (2021) Offline: The case for No-COVID. Lancet 397(10272): 359.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania