Antimicrobial Activity of Lactic Acid Bacteria (Lab) Isolated from Yoghurt and Fish against Pathogenic Bacteria Isolated from Fish in Ethiopia
Pathogenic bacteria are causative agent of infectious diseases in fish. Antimicrobials are widely applied in fish products for prevention of fish-borne diseases. The global concern of antimicrobial resistance of bacteria raises attentiveness of the different ways of using antimicrobial peptides (AMPs). The aim of the present study was to detect the antimicrobial activity of the AMPs from Lactic acid bacteria (LAB) against the growth of pathogenic bacteria isolated from fish in Ethiopia. Cell Free Supernatants (CFSs) from 4 strains of LAB consisting of lactobacillus paracasei sub.paracasei, lactobacillus casei LC2W, Pediococcus acidilactici and lactobacillus plantarum were tested against pathogenic bacteria of Aeromonas hydrophila, Aeromonas veronii, Edwardsiella tarda, Escherichia coli, Proteus mirabilis, Pseudomonas fluorescens and Shigella flexneri via using agar well diffusion assay. All tested CFSs revealed antimicrobial activity against the tested fish pathogenic isolates at different extents. CFSs of lactobacillus plantarum, Lb. casei LC2W and lactobacillus paracasei sub.paracasei showed strong antibacterial activity against Edwardsiella tarda (19 mm), Aeromonas hydrophila (18 mm) and Pseudomonas fluorescens (18 mm) respectively, with the well diffusion assay. The study shows the antimicrobial activity of the 4 strains of LAB CFSs against the bacterial fish pathogens and indicates the application opportunities as fish product bio preservative.
Introduction
Pathogenic bacteria such as Aeromonas, Edwardsiella, and Pseudomonas are an important causative agent for infectious diseases in fish and cause significant loss in fish products [1]. Most of the commercially available food preservatives are synthesized chemically, causing a lot of toxic effect on human health. An ever growing demand for food products with minimal chemical additives has generated a necessity for exploring new alternatives for food preservation. Lactic acid bacteria (LAB) can produce a wide range of inhibitory compounds to reduce pathogens invasion. These include bacteriocins, organic acids, ethanol, diacetyl, carbon dioxide, and hydrogen peroxide [2]. Bacteriocins are ribosomally synthesized Antimicrobial Peptides (AMPs) produced by both Gram negative and Gram positive bacteria. Bacteriocins produced by LAB referred here as LAB bacteriocins are most often devoid of cytotoxic traits [3], and endowed with antagonistic functions as well as additional beneficial attributes [4]. LAB bacteriocins are emerging as a novel wave of antibiotics with potent in vitro and in vivo activities [5].
In contrast to traditional antibiotics, LAB bacteriocins target specific species and do not affect other population within the same ecosystem. LAB bacteriocins are known to exert either bacteriostatic or bactericidal activity toward sensitive organisms [3]. Their modes of action have been widely but not thoroughly investigated. Recent insights on modes of action are reviewed elsewhere [6]. Combinations of LAB bacteriocins and antibiotics are emerging as novel therapeutic options for food producing animals [7]. Different reports have established the main advantages and synergistic actions of LAB bacteriocins with other biomolecules. These are the case of enterocin AS-48 and ethambutol against Mycobacterium tuberculosis [6], nisin and citric acid against Staphylococcus aureus and Listeria monocytogenes, nisin and beta-lactams against Salmonella enterica serovar Typhimurium, and Garvicin KA-farnesol against a set of Gram positive and Gram negative bacteria [8].
Orally administration of these substances is a challenge because of their enzymatic degradation. This case was reported in vivo for lacticin 3147 and nisin [9]. Organic acids, including short chain fatty acids, lactic and formic acids, were shown to inhibit potentially pathogenic bacteria of importance for livestock animals. LAB is producing lactic acid as the main product of sugar metabolism [10]. However, LAB metabolically known as heterofermentative species can concomitantly produce other end products such as acetic acid. Organic acids are known to act by reducing the intracellular pH and inhibiting the active transport of excess internal protons which requires cellular adenosine triphosphate (ATP) consumption leading to cellular energy depletion [11]. The main targets of organic acids are the bacterial cell wall, cytoplasmic membrane, and specific metabolic functions like replication and protein synthesis of pathogenic microorganisms leading to their disturbance and death [12].
LAB isolates inhibit pathogenic bacteria by various mechanisms, including production of antimicrobial metabolites. Cell free supernatants (CFSs), a peptide produced by LAB widely used as a natural and safe preservative in food products. CFS from the LAB isolates was strongly inhibited the growth of food-borne bacterial pathogens. Inhibitory activity was dependent primarily on active substance(s) secreted into the supernatant [13]. The inhibitory substance(s) synthesized by LAB are proteinaceous in nature and act differently against common pathogenic and spoilage bacteria. Thus, the antimicrobial activity of the CFS of LAB could serve as potential bio preservative to control food borne pathogens and spoilage bacteria [14, 15].
Lactic acid produced by LAB induces an unfavorable local microenvironment for pathogenic bacteria [16]. However, the diversity of fish microbiota and the different antimicrobial effects of LAB CFSs tested indicate the need for further research into these antimicrobial activities. Therefore, the aim of the present study was to investigate the antimicrobial activities of CFSs of LAB isolated from yoghurt and fish intestine on the major bacterial pathogens isolated from fish in Ethiopia.
Materials and Methods
Study Area and Design
Laboratory based experimental study was conducted to identify the antimicrobial activity of LAB isolated from yoghurt and fish intestine at Addis Ababa University, College of Natural and Computational Science, Institute of Biotechnology (health Biotechnology Laboratory).
Sample Collection
Naturally infected and apparently healthy fish were collected from selected Ethiopian rift valley lakes (Hawassa, Langanoo and Ziway) as a source for isolation of bacterial pathogens, Aeromonas hydrophila, Aeromonas veronii, Edwardsiella tarda, Escherichia coli, Proteus mirabilis, Pseudomonas fluorescens and Shigella flexneri.
Fish Samples
The fish samples were dissected using a sterile dissecting scissor by following the Leal, et al. [17] procedures, and tissue samples of kidney, intestine and liver were taken aseptically using sterile scalpel blade (forceps) and kept in sterile universal bottles of 100 ml capacities separately and homogenized in physiological saline solution.
Isolation and Identification of Pathogenic Bacteria
Inoculums were taken from each sample of physiological saline solution homogenized intestine, liver and kidney using sterile inoculating loops and inoculated on Nutrient Agar (HIMEDIA, India) and incubated at 37°C for 24 hours under aerobic condition. Each type culture colony was picked up and sub cultured on selective and differential media EMB agar medium and XLD agar media (HIMEDIA, India) and incubated at 37°C for 24 hours. Suspected bacterial colonies were picked up and inoculated into Tryptone Soy Broth (TSB) (HIMEDIA, India) and incubated at 37°C for another 24 hours.
Morphological Observations of Isolates
For the identification of selected isolates, colony morphology such as colony form, elevation, margin, surface and pigmentation were studied. Colony color was noted by visual inspection and bacterial cell suspension using fresh culture was used for microscopic examination of isolates with simple staining and differential staining (Gram- staining) following Society of American Bacteriologists (SAB) procedure [18]. The morphologically identified isolates were stored at -20°C in 50% glycerol (Fine Chemical, Ethiopia) using cryovial tubes of 1.8ml (IMEC, China) for further biochemical identification.
Biochemical Studies of the Isolates
All isolates were identified biochemically by streaking bacterial colonies over TSB and incubated at 37°C for 24 hours and, then identified by using a range of biochemical tests performed according to respective manufacturer’s instructions. Following Society of American Bacteriologists (SAB) [18] and Bergey’s manual of determinative bacteriology [19] procedure, the biochemical tests of the isolated bacteria were carried out on February 2022.
DNA Extraction
In order to extract the DNA, bacterial isolates were grown overnight in 10 ml of TSB broth at 37°C for 24 hours. Extraction of DNA from each bacterial isolate was done by using Qiagen Flexi Gene DNA kit and it was stored at -20 °C till use [20].
PCR Amplification
Polymerase Chain Reaction (PCR) amplification was performed in a DNA thermal cycler (Eppendorf, Hamburg, Germany). PCR reactions were performed in a final volume of 25μl containing 3μl of template DNA, 2.5μl buffer solution,
14μl double distilled water (ddH2o), 1μl of each primer rD1(5’-CCCGGGATCCAAGCTTAAGGAGGTGATCCAGCC-3’) and fD1 (5’CGAATTCGTCGACAACAGAGTTTGATCCTGGCTCAG-3’), 2μl of MgCl2 solution, 0.5μl Taq DNA polymerase and 1μl deoxynucleotide triphosphate mix (dNTPs). PCR conditions were as follows: 95 °C for 15 min, 30 cycles at 95 °C for 45 s, 55 °C for 45 s, 72 °C for 1 min, followed by a final elongation at 72 °C for 7 min. The PCR products (8μl) were analyzed by electrophoresis on 1.5% agarose gels stained with ethidium bromide using 1 μM Tris-Acetate-EDTA buffer at 100v for 1 hour and were visualized by UV transillumination [21].
DNA Sequencing
The PCR products were sequenced with 16S universal bacterial primers mentioned above (rD1 and fD1) by Sanger sequencing techniques at BaseClear DNA research laboratory, Leiden, The Netherlands). For sequencing of amplified 16S rRNA directly, four identical 100µl amplification reactions were performed on each sample, with the resultant material being pooled and purified. A 500ng amount of template (16S rRNA) was combined with 10 ng of primer, 2µl of Sequence buffer, and water to 10µl. This sample was held at 98°C for 7 min and cooled to room temperature for 1 min, and then the labeling reaction was performed at 37°C for 5 min. Chain elongation was terminated with sample loading buffer, and sequencing was performed on buffer gradient gels [22].
NCBI Accession Number
The 16S rRNA gene sequences of Aeromonas hydrophila, Aeromonas veronii, Edwardsiella tarda, Escherichia coli, Proteus mirabilis, Pseudomonas fluorescens and Shigella flexneri have been submitted to NCBI- GenBank under accession number of KC906257, LC420134, DQ884466, KJ643928, MK802107, MF419185 and MT604864 respectively.
Lactic Acid Bacteria Strain
The common four strains used in this study were lactobacillus paracasei sub.paracasei, lactobacillus casei LC2W, Pediococcus acidilactici and lactobacillus plantarum. The three strains (lactobacillus paracasei sub.paracasei, lactobacillus casei LC2W and Pediococcus acidilactici) were obtained from the collection center of Ethiopian National Agricultural Biotechnology Research Center, Microbial Biotechnology Laboratory of Holetta branch, previously isolated and identified from yoghurt and one strain (lactobacillus plantarum) was isolated from apparently healthy fish intestine and identified following growth on MRS Agar followed by phenotypic (morphological, microscopic and biochemical tests) characterization.
Data Analysis
All measurements of antagonistic activity were performed in triplicate. Data were analyzed within one-way ANOVA and statistical analysis was performed using SPSS 26 (IBM Statistics, Armonk, NY) statistical software. P ≤ 0.05 was considered as the statistical significance level.
Preparation of Cell Free Supernatants (CFSs)
Strains to be tested for antimicrobial activity were incubated in MRS broth for 48 h at 37°C. Bacterial cells were removed by centrifugation of the culture at 5000 × g for 20 min at 4°C. After centrifugation, the supernatants were filtered using syringe filters Luer Lock having pored size 0.22 μM (MF-Millipore, USA). The pH values of supernatants were adjusted to pH 7.0 by the addition of 1N NaOH and stored at 4°C until use [23].
Cell Free Supernatants Characterization
Effect of pH: The effect of pH on the CFSs antimicrobial activity were determined by adjusting the pH of CFS using 1N HCl, 1N NaOH, and pH meter following that pH 3, pH 5, pH 7, pH 9, and pH 11 were used. After 3 hours of incubation at room temperature, the antimicrobial activity of treated CFSs against E. coli was assayed using the agar diffusion assay [24]. Effect of Temperature: The effect of heat treatment at 40 to 121°C was carried out in test tubes overlaid with paraffin oil and covered with aluminum foil to prevent evaporation during heating for 10 min. The antimicrobial activity of heated CFSs against E. coli was carried out using the agar diffusion assay [24]. Effect of Amylase and Proteolytic Enzymes: The effect of amylase and proteolytic enzymes on the activity of CFSs was conducted according to the Zhou F, et al. [25] method. The crude CFS was treated with 5mg/ml of amylase (BIO BASIC Canada INC), trypsin (BIO BASIC Canada INC) and pepsin (BIO BASIC Canada INC). Incubation was held at 37°C for 1 h. After incubation, the enzymes were denatured by heating the samples at 80°C for 10 min, and the residual CFS antimicrobial activity against E. coli was carried out using the agar diffusion assay [24].
Antimicrobial Assay
The antimicrobial activity of LAB was determined by a well-diffusion assay as described by Kojic, et al. [24]. The antimicrobial susceptibility was initially assayed by the agar well-diffusion assay on the Mueller Hinton agar (MHA). 100μl of each CFS was prepared. The fish pathogenic bacteria cell suspensions were adjusted to 0.5 McFarland turbidity standards to prepare 1x108 bacteria per ml inoculums. Each bacterial suspension was inoculated on MHA plates using sterilized cotton swab, and the plates were then allowed to dry for 5 minutes. Wells were punctured in the medium using sterilized 6mm sized micropipette tips then sealed with sterile molten agar. Cell-free supernatant (CFS) (100μl) was added to the respective wells. MRS broth was used as a negative control. CFSs were neutralized to pH 7 to maintain that the inhibition was not due to lactic or other organic acids but by antimicrobial substances. After 48 h incubation at 37°C aerobically, the diameter of zones of inhibition (mm) was measured.
Result
Morphological Identification
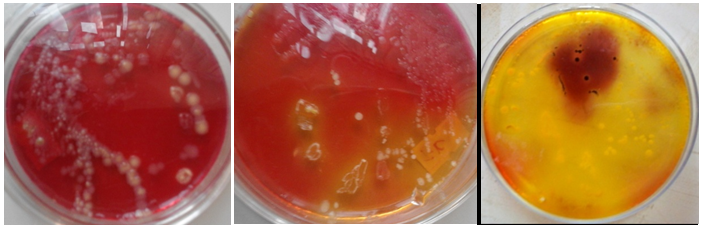
The morphological characteristics of the bacterial pathogens strains were determined including Gram staining, bacterial colony’s and bacterial cell’s morphology. On the basis of colony morphology, all strains were short rod and non- spore former. Colonies of the isolated isolates were found to be different in their form, elevation, margin, surface, colour and optical characteristics (Figures 1 & 2).
Biochemical Characterization
To identify the isolated bacteria, a range of biochemical tests (Indole, methyl red, Voges-Proskauer (VP), Citrate, H2S gas, Urease, sugar fermentation and catalase test) were carried out. The results showed that different groups of bacterial pathogens were identified. These were Aeromonas hydrophila, Aeromonas veronii, Edwardsiella tarda, Escherichia coli, Proteus mirabilis, Pseudomonas fluorescens and Shigella flexneri.
Analysis of 16S rRNA Gene
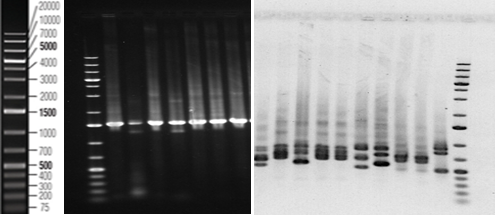
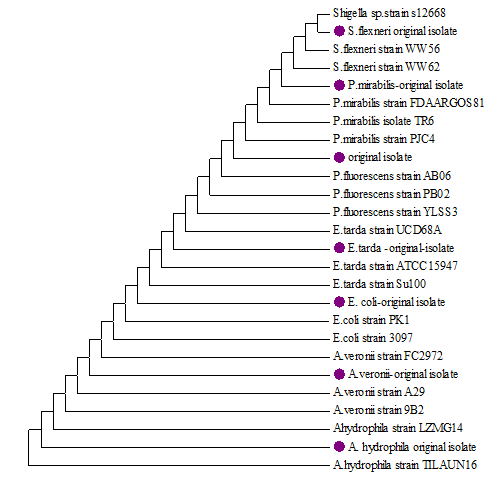
The seven identified isolates by morphological characteristics and biochemical tests were subjected to further identify by 16S rRNA (1-7) and sequenced by Sanger sequencing (BaseClear DNA research laboratory, Leiden, The Netherlands) and were compared to those available in the GenBank database (Figure 3). The universal primers (rD1 and fD1) were used to amplify of 16S rRNA of bacterial isolates, with amplicon size ∼1.4kbp (Figure 3). Compared with GenBank database, the nucleotide sequences of 16S rRNA gene could detect the isolates of bacteria in the level species, according to levels of homology of nucleotide ranging from 97.65-100% (Figure 4).
Antibacterial Potential of LAB CFS on Pathogenic Organism
The antibacterial activity of the 4 LAB strains CFS against the tested fish pathogenic bacteria were confirmed by the presence or absence and magnitude of inhibition zones around well on the agar plates. As presented in Table 1, CFS of some isolates exhibited strong inhibitory effects against all the tested bacterial fish pathogens. In this assay, most isolates exerted consistent antibacterial effects against each pathogen with a zone of inhibition diameters ranging from 9 to 19 mm. MRS medium, used as a negative control, had no inhibitory effect (Table 1). Among the test pathogens, the most sensitive were Edwardsiella tarda for Lactobacillus plantarum and Lactobacillus casei LC2W, while Aeromonas hydrophila and Pseudomonas fluorescens were more sensitive to Lactobacillus casei LC2W and Lactobacillus paracasei sub. paracasei CFSs respectively. Proteus mirabilis was the least sensitive in all cases.
| Inhibition zone (mm) | |||||||
|---|---|---|---|---|---|---|---|
| Isolates | Aeromonas hydrophila | Aeromonas veronii | Edwardsiella tarda | Escherichia coli | Proteus mirabilis | Pseudomonas fluorescens | Shigella flexneri |
| Lactobacillus paracasei sub. paracasei | 14±0.53a | 12±0.44a | 15 ±0.31a | 17±0.57a | 9±0.39b | 18±0.63b | 10±0.52b |
| Lactobacillus casei LC2W | 18±0.74a | 13±0.76a | 16±0.79a | 11±0.61b | 12±0.67a | 14±0.51a | 17±0.63a |
| Pediococcus acidilactici | 13 ±0.50a | 11 ±0.51a | 12±0.72b | 15±0.42a | 14±0.54a | 12±0.77a | 13±0.54a |
| Lactobacillus plantarum | 16 ±0.64a | 14 ±0.24a | 19 ±0.62a | 17±0.51a | 15±0.50a | 17±0.42b | 16±0.36a |
| NC (MRS) broth | − | − | − | − | − | − | − |
Table 1: Inhibition of various bacterial pathogens by CFS produced by the selected lactic acid bacteria (LAB).
NC: Negative control; −: no inhibition zone. All values are means of triplicates SD. Levels not connected by same letter under the same column are significantly different (p≤0.05). Table 1: Inhibition of various bacterial pathogens by CFS produced by the selected lactic acid bacteria (LAB).
Inhibitory Activity of CFS following Heat Treatments
The CFS produced by the four LAB isolates were heat treated at 40, 60, 100 and 121◦C for 10 minutes. The antimicrobial compound in CFSs was thermally stable Inhibition zone (mm) at different temperatures Isolates 40◦C, 10 min 60◦C, 10 min 100◦C, 10 min 121◦C,10 min Lactobacillus paracasei sub. paracasei 13 ±0.62a 8 ±0.72a 2 ±1.00a − Lactobacillus casei LC2W 10 ±0.51a 9 ±0.79a 3 ±0.48a 1 mm Pediococcus acidilactici 11±0.42a 7 ±0.52a 1 ±0.52a − Lactobacillus plantarum 12 ±0.50a 8 ±0.65a 2 ±0.71a − Note: no inhibition zone. All values are means of triplicates SD. Levels not connected by same letter under the same column are significantly different (p≤0.05). Table 2: Sensitivity of antimicrobial activity of CFS produced by the LAB at various heat treatments (Escherichia coli was used as an indicator pathogen).
Inhibitory Activity of CFS following Enzyme Treatments
The sensitivity of the CFS to trypsin, pepsin, and Amylase was determined in controlled and reproducible conditions under 40, 60 and 100◦C heat conditions as their inhibitory effects against Escherichia coli were observed (Table 2). Nevertheless, CFS lost activity after temperature treatment at 121°C for 10 min displaying no inhibition zones compared to the control.
as shown in Table 3. The inhibitory substance was fully inactivated by proteolytic (trypsin and pepsin) enzymes, while the amylase enzyme had no effect on the antagonistic activity.
| Inhibition zone (mm) | |||
|---|---|---|---|
| Isolates | Amylase | Trypsin | Pepsin |
| Lactobacillus paracasei sub. paracasei | 11 ±0.50a | − | − |
| Lactobacillus casei LC2W | 7 ±0.63a | − | − |
| Pediococcus acidilactici | 10 ±0.51a | − | − |
| Lactobacillus plantarum | 9 ±0.72a | − | − |
Table 2: The inhibitory substance was fully inactivated by proteolytic (trypsin and pepsin) enzymes, while the amylase enzyme had
Note: no inhibition zone. All values are means of triplicates SD. Levels not connected by same letter under the same column are significantly different (p≤0.05). Table 3: Inhibitory activity of CFS produced by the LAB treated with various enzymes (Escherichia coli was used as an indicator pathogen).
Inhibitory Activity of CFS at Different pH Condition
The exposure of CFS to different pH values showed Inhibition zone (mm) at different pHs Isolates pH 3 pH 5 pH 7 pH 9 pH 11 Lactobacillus paracasei sub. paracasei 18 ±0.56a 15 ±0.50a 16 ±0.60a 13 ±0.55a 5 ±0.60a that all antimicrobial substances remained fully active in the pH range from 3-11. Reduced activities of antimicrobial substances from CFS of all isolates were found after treatment at pH 11.0 (Table 4).
Lactobacillus casei LC2W 12 ±0.78b 17 ±0.79a 10 ±0.42b 8 ±0.63a 2 ±0.57a
Pediococcus acidilactici 16 ±0.53a 14 ±0.63a 15 ±0.53a 11 ±0.13a 3 ±0.74a
Lactobacillus plantarum 18 ±0.61a 16 ±0.74a 15 ±0.61a 12 ±0.54a 4 ±0.24a
- All values are means of triplicates SD. Levels not connected by same letter under the same column are significantly different
- (p≤0.05).
Table 3: Inhibitory activity of CFS produced by the LAB at various pHs (Escherichia coli was used as an indicator pathogen).
Discussion
The antimicrobial activities of the Cell free supernatant against Aeromonas hydrophila, Aeromonas veronii, Edwardsiella tarda, Escherichia coli, Proteus mirabilis, Pseudomonas fluorescens and Shigella flexneri (Table 1) revealed that the four LAB strains produce Cell free supernatant able to inhibit these pathogens growth with different inhibition zones and suggesting that this antimicrobial activity is strain dependent. Similarly, Putra TF, et al. [26] and Dejene F, et al. [27] suggested that lactic acid bacteria Cell free supernatant isolated from various fermented foods and fish can suppress a number of pathogenic and food borne pathogenic organisms growth. Khalil RA, et al. [28] also suggests that food derived LAB strains exhibited an antagonistic effect against different pathogenic bacteria and spoilage fungi and could be selected for biotechnological application to control pathogenic bacteria and spoilage fungi. Numerous strains of LAB isolated from the gastrointestinal (GI) tract of finfish are able to produce antibacterial substances toward different potential fish pathogenic bacteria as well as human pathogens and also become the most promising bacterial genera as probiotic in aquaculture [29]. The greatest antimicrobial activity against each pathogen was found in Lactobacillus plantarum, which showed strong antimicrobial activity against Edwardsiella tarda, but it showed relatively low antimicrobial activity against Aeromonas veronii. This shows agreement with the finding of Kaewchomphunuch T, et al. [15] which indicated that the CFS of Lactobacillus plantarum isolated from pigs showed strongest antibacterial activity against food-borne pathogenic E. coli. When the inhibition zones produced by each isolate of LAB strain were compared, the cell free supernatant of those isolates exhibited a significant (p<0.05) antibacterial effect against the majority of tested fish pathogenic bacteria between groups as demonstrated by one-way ANOVA (Table 1). But the result showed that there was no statistically significant difference between the groups that is treated against Aeromonas hydrophila and Aeromonas veronii pathogens. A complex and dynamic community of microorganisms play important roles within the fish gastrointestinal (GI) tract of which LAB are generally considered as favorable microorganism due to their abilities to stimulating host GI development, digestive function, and mucosal tolerance, stimulating immune response, and improved disease resistance [29].
The antimicrobial components of the four LAB strains, Pediococcus acidilactici, Lb. casei LC2W, Lactobacillus paracasei sub.paracasei and Lactobacillus plantarum are most likely proteinous CFS. This is characterized by no loss in their inhibitory activities even though the pH of the supernatant was adjusted to 11. The CFS of Lactobacillus casei LC2W is a protein or peptide resistant to high temperature (Table 2). The antimicrobial potential of heat killed supernatant supports the thermostable nature of the antimicrobial compounds [30, 4]. CFS from the four LABs strains is shown to induce antimicrobial activity resistant to various heating temperatures and pH. The CFS antagonistic activity of isolated bacteria can be influenced by a few proteolytic enzymes driving to loss in their antagonistic action. In this study the sensitivity of the produced CFS was further tested using various enzymes with Escherichia coli as indicator pathogen and the results are presented in Table 3. These results indicated that treatment with trypsin and pepsin resulted in the loss of antimicrobial activity of CFS against Escherichia coli and suggested that the component of the CFS responsible for the antimicrobial effect is not starch but protein. This statement was supported by the loss of antimicrobial activity of CFS treated with proteolytic enzymes such as trypsin and proteinase K. Thus, the current results showed that the CFS produced by LAB is a highly thermostable proteinaceous compound with antimicrobial activity even after autoclaving at 121°C [24]. Most of the LAB bacteriocins are peptide compounds, heat resistant, cationic, and amphiphilic [31].
The antagonistic activity of CFS illustrated by the four isolates of LAB was also pH dependent. The foremost antagonistic activity was displayed in the acidic pH range of 3 to 7, whereas a decrease of antagonistic was observed in alkaline pH condition (pH > 12). The same result was reported by Dejene F, et al. [27] in which LAB strains of Enterococcus NCFS, Pediococcus NCFS, Leuconostoc NCFS and Weissella NCFS isolated from fermented beverage and finfish, appeared to have the most elevated antagonistic activity and stability at pH 2 and 5.
Conclusion
CFS produced by the four isolates of LAB showed antimicrobial activity against a wide range of bacterial fish pathogens, indicating its potential application as a fish products bio preservative. These antibacterial LAB strains products can be used in the food industry instead of chemical preservatives to produce organic foods. Furthermore, LAB may preserve the nutritional value of fish products and delay spoilage. From a health perspective, the conventional antimicrobial agents usually provide effective antibiotic therapy for bacterial infections. Therefore, using antimicrobial compounds from LAB may inhibit bacteria having clinical value in treating certain fish-borne microbial infection. Further studies will help make these findings applicable to food safety.
Funding
The Office of the Director for Research, Addis Ababa University, financially supported this study.
Author Contributions
GD conceived and designed the experiments, collected samples, performed the laboratory works, analyzed the data and drafted the manuscript. BL and HM conceived and designed the experiments, critically comment and revised the manuscript.
Competing Interests
The authors declare that there is no competing interest.
Data Availability and Materials
All data and materials are within the manuscript.
Ethics Approval
The study has got ethical approval by the College of Natural and Computational Sciences Institutional Review Board (IRB), Addis Ababa University.
Consent for Publication
This is not applicable.
Acknowledgement
The authors are grateful to the fishermen in the three study Lakes for their technical assistance during sample collection.
References
-
Walczak N, Puk K, Guz L (2017) Bacterial flora associated with diseased freshwater ornamental fish. J Vet Res 61(4): 445-449.
-
Liao SF, Nyachoti M (2017) Using probiotics to improve swine gut health and nutrient utilization. Anim Nutr 3(4): 331-343.
-
Drider D, Bendali F, Naghmouchi K, Chikindas ML (2016) Bacteriocins: not only antibacterial agents. Probiotics Antimicrob Proteins 8(4): 177-182.
-
Chikindas ML, Weeks R, Drider D, Chistyakov VA, Dicks LM (2018) Functions and emerging applications of bacteriocins. Curr Opin Biotechnol 49: 23-28.
-
Seddik HA, Bendali F, Gancel F, Fliss I, Spano G, et al. (2017) Lactobacillus plantarum and its probiotic and food potentialities. Probiotics Antimicrob Proteins 9(2): 111-122.
-
Pérez C, Gracia B, Rodrigues L, Vitoria A, Cebrián R, et al. (2018) Synergy between circular bacteriocin AS-48 and ethambutol against Mycobacterium tuberculosis. Antimicrob Agents Chemother 62(9): e00359-e00418.
-
Al Atya AK, Belguesmia Y, Chataigne G, Ravallec R, Vachée A, et al. (2016) Anti-MRSA activities of enterocins DD28 and DD93 and evidences on their role in the inhibition of biofilm formation. Front Microbiol 7: 817.
-
Chi H, Holo H (2018) Synergistic antimicrobial activity between the broad spectrum bacteriocin garvicin KS and nisin, farnesol and polymyxin B against Gram-positive and Gram-negative bacteria. Curr Microbiol 75(3): 272- 277.
-
Gough R, Rubio RC, Connor PMO, Crispie F, Brodkorb A, et al. (2018) Oral delivery of nisin in resistant starch based matrices alters the gut microbiota in mice. Front Microbiol 9: 1186.
-
Russo P, Arena MP, Fiocco D, Capozzi V, Drider D, et al. (2017) Lactobacillus plantarum with broad antifungal activity: a promising approach to increase safety and shelf-life of cereal-based products. Int J Food Microbiol 247: 48-54.
-
Ricke SC (2003) Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult Sci 82(4): 632-639.
-
Nair MS, Amalaradjou MA, Venkitanarayanan K (2017) Antivirulence Properties of Probiotics in Combating Microbial Pathogenesis. Adv Appl Microbiol 98: 1-29.
-
Mariam HS, Zegeye N, Tariku T, Andargie E, Endalafer N, et al. (2014) Potential of cell-free supernatants from cultures of selected lactic acid bacteria and yeast obtained from local fermented foods as inhibitors of Listeria monocytogenes, Salmonella spp. and Staphylococcus aureus. BMC Res Notes 7: 606.
-
John GE, Lennox JA (2019) Antibacterial Activity of Cell Free Supernatant of Lactic Acid Bacteria Isolated From Food Samples against Food Borne Pathogens. International Journal of Scientific and Engineering Research 10(6): 1037-1054.
-
Kaewchomphunuch T, Charoenpichitnunt T, Thongbaiyai V, Ngamwongsatit N, Kaeoket K (2022) Cell-free culture supernatants of Lactobacillus spp. and Pediococcus spp. inhibit growth of pathogenic Escherichia coli isolated from pigs in Thailand. BMC Vet Res 18(1): 60.
-
Dittoe DK, Ricke SC, Kiess AS (2018) Organic acids and potential for modifying the avian gastrointestinal tract and reducing pathogens and disease. Front Vet Sci 5: 216.
-
Leal MC, Cardoso ER, Nóbrega RH, Batlouni SR, Bogerd J, et al. (2009) Histological and stereological evaluation of zebra fish (Danio rerio) spermatogenesis with an emphasis on spermatogonial generations. Biol Reprod 81(1): 177-187.
-
Society of American Bacteriologists (SAB) (1957) Manual of microbiological methods. McGraw Hill, New York, pp: 315.
-
Holt JG, Bergey DH (1994) Bergey’s manual of determinative bacteriology. 9th(Edn.), William & Wilkins, London.
-
Flexi Gene DNA Handbook (2014) QIAGEN Sample and Assay Technologies for purification of DNA from human whole blood, Buffy coat and cultured cells.
-
Weisburg GW, Barns MS, Pelletier AD, Lane JD (1991) 16S ribosomal DNA Amplification for Phylogenetic Study. J Bacteriol 173(2): 697-703.
-
Biggin MD, Gibson TJ, Hong GF (1983) Buffer gradient gels and 35S label as an aid to rapid DNA sequence determination. Proc Natl Acad Sci U S A 80(13): 3963- 3965.
-
Arena MP, Silvain A, Normanno G, Grieco F, Drider D, et al. (2016) Use of Lactobacillus plantarum strains as a bio-control strategy against food-borne pathogenic microorganisms. Front Microbiol 7: 464.
-
Kojic M, Svircevic J, Banina A, Topisirovic L (1991) Bacteriocin- Producing Strain of Lactococcus lactis subsp. diacitilactis S50. Appl Environ Microbiol 57(6): 1835-1837.
-
Zhou F, Zhao H, Bai F, Piotr D, Liu Y, et al. (2014) Purification and characterisation of the bacteriocin produced by Lactobacillus plantarum, isolated from Chinese pickle. Czech Journal of Food Sciences 32(5): 430-436.
-
Putra TF, Suprapto H, Tjahjaningsih W, Pramono H (2018) The antagonistic activity of lactic acid bacteria isolated from peda, an Indonesian traditional fermented fish. IOP Conference Serious. Earth Environ Sci 137: 012060.
-
Dejene F, Dadi BR, Taddesse D (2021) In Vitro Antagonistic Effect of Lactic Acid Bacteria Isolated from Fermented Beverage and Finfish on Pathogenic and Food borne Pathogenic Microorganism in Ethiopia. International Journal of Microbiology 210: 1-10.
-
Khandakar RA, Kabir Y, Islam R, Hossain S, Paul S, et al. (2014) Antagonism of Lactic Acid Bacteria against Selected Pathogenic Bacteria and Spoilage Fungi. American-Eurasian Journal of Agriculture and Environmental Science 14(3): 193-198.
-
Ringø E, Hoseinifar SH, Ghosh K, Doan HV, Beck BR, et al. (2018) Lactic Acid Bacteria in Finfish-An Update. Front Microbiol 9: 1818.
-
Perez RH, Zendo T, Sonomoto K (2014) Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microbial Cell Fact 13(Suppl 1): S3.
-
Zacharof MP, Lovitt RW (2012) Bacteriocins produced by lactic acid bacteria. A review article. APCBEE Procedia 2: 50-56.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania