Production of Single Cell Oils from Bacterial Sources
Single Cell Oils (SCO) is of profound interest for a variety of purposes ranging from biofuels to nutritional adjuvants, pharmaceutical applications and biotransformation for valuable products. A number of microorganisms have been shown to produce and accumulate SCO. In the present study a methodical attempt was made to isolate potential SCO producers from Indian water sources. Saltwater samples from the Arabian Sea and freshwater samples from an Indian cold-water river (Pindhari River, Uttarakhand) were collected and studied for the occurrence of lipid producing microbes. Of the several isolates shortlisted as lipid producers three isolates from this study were identified as potential SCO producers based on their lipid producing abilities. The types of fatty acids comprising the SCO from selected isolates were studied by Gas Chromatography (GC) and confirmed by Gas Chromatography/Mass Spectrometry (GC/MS). Lipid profiles from the GC analysis showed that the isolates in this study produced economically and nutritionally valuable Monounsaturated fatty acids (MUFA) like Palmitoleic acid and Oleic acid. Also, two isolates from the Arabian Sea were seen to produce a valuable omega-3 Polyunsaturated Fatty Acid (PUFA) like Eicosapentanoic acid. While a freshwater isolate produced Linoleic acid an omega-6 PUFA. Selected isolates were characterized for their biochemical characteristics and identified molecularly by 16S rRNA sequencing. Ornithinibacillus sp. Marseille-P3601 strain isolated in our study from the cold-water River Pindhari, Uttarakhand is found capable of producing PUFA.
Introduction
All living organisms produce lipids and oil for various structural and functional roles. Some classes of microorganisms are known for producing microbial oil or single cell oil (SCO), which are storage lipids accumulated, composed majorly of triacylglycerols (TAGs) [1, 2]. SCO formation has been known for over 100 years, it is only within the last 20 years or so that attempts have been made to understand the process of lipid accumulation in the oleaginous organisms [2]. SCOs, in last 2 decades have gained commercial importance for various biotechnological products ranging from food supplements, biodiesel to oleo chemicals and high value products like PUFAs [3].
Microorganisms having high lipid accumulating capability i.e., between 20%-80% of their biomass can be considered as oleaginous microorganisms [1, 4]. Lipid accumulation occurs in the stationary phase, and is associated with excess of carbon coupled with limitation of nitrogen, phosphorous etc [3]. Some of the common oleaginous organisms include genus Mortierella, Cryptococcus etc [5, 6].
Polyunsaturated Fatty Acids (PUFA), also known as Omega-3 fatty acids, are a family of unsaturated fatty acids that have in common a final carbon-carbon double bond in the n-3 position likeα-Linolenic Acid (ALA) 18:2n-3, Eicosapentanoic Acid (EPA) 20:5n-3, Arachidonic Acid (AA) 20:2n-6, Docosahexanoic acid (DHA) 22:6n3 etc. They are known for significant they play in general well-being of an individual [7]. Being essential fatty acids PUFA are preventive and co-therapeutic for several major diseases such as coronary heart disease, atherosclerosis, cancer and Type II diabetes and disorders like arthritis, mental decline and auto immune conditions. They also reduce the clotting ability of platelets, thus potentially decreasing the incidence of heart attacks and strokes [8, 9, 10]. Oily fishes (like mackerel, salmon, striped bass, rainbow trout, halibut, tuna and sardines), vegetable sources and seed oils (like olive oil, rapeseed, chia, perilla, pursalane, lingoberry flax seed oil, linseed and hemp seed oils), are some of the rich sources of PUFAs [8, 11, 12].
Since, humans cannot synthesize the essential fatty acids, they need to be taken as supplements [13]. Though rich source of fats, fish oils are less preferred because of unpleasant odour, seasonal variations, contamination with heavy metal, toxins and pollutants. Moreover, worldwide fish stocks are under threat, therefore, newer microbial sources are being sought for production of SCO [6, 14]. Recently, certain algal, fungal, bacterial sources including marine sources, also have been proven as rich sources of PUFA [3, 15]. From a total of 27 isolates from both marine and fresh water sources that were seen to be strongly positive for lipid accumulation, we have presented results for 3 PUFA producers in this paper.
Materials and Methods
All microbiological media were obtained from Himedia®, India and the chemicals were procured from Merck® Specialty Chemicals Limited, Germany.
Sampling
Water samples were collected from 2 sources namely- Arabian Sea water from a coastal village named Pallande situated in western Maharashtra, and fresh water from a cold river Pindhari situated in Uttarakhand, India. The samples were collected in sterilized airtight 100ml plastic screw capped bottles and stored at 0ᵒC till further analysis.
Isolation of Microbes
Similar to the protocol followed by Patnayak and Sree, 2005 [16] for marine samples, sea water sample collected was serially diluted in saline followed by pour plating in Salt Water Complete Medium (Composition w/v: 0.5g Peptone, 0.3g Yeast Extract, 0.3 ml Glycerol, 3g Agar in 75% Sea Water) The plates were incubated at 25oC for 120h. Similarly, M9 medium and Alkaline M9 (pH adjusted to 9) was used as plating media for freshwater samples [17]. Based on morphological colony characteristics, colonies were shortlisted and transferred to master plate. The samples from Pindhari river were incubated at 20oC, owing to the year-round cooler temperatures of the river.
Preliminary Screening
For Lipid Accumulation: The isolates obtained from water samples needed to be screened for the lipid accumulation ability. Three parameters were selected for the preliminary screening-i) intensity of Sudan Black B staining, ii) biomass yield on nitrogen limited semi-synthetic medium and iii) percentage of lipid quantity against biomass [18]. *All experiments were carried out in triplicates). Sudan Black B staining method was adopted to visually see the lipid accumulation in bacterial cells [19].
Induction of Lipid Accumulation : The isolates obtained upon 24hrs of incubation on complete media [16, 18] were transferred to solid Semi synthetic minimal medium; to induce lipid accumulation under nitrogen limitation and incubated at 25oC for 120h.
Intensity of Lipid Accumulated: Three isolates with most intense Sudan Black B stain under 1000X magnification, were shortlisted. The selected cultures were cultured thrice on Semi synthetic minimal agar medium, stained and observed to confirm the reproducibility of lipid accumulation [20]. Amount of Biomass on Semi-Synthetic Medium [21]: The 3 selected isolates were cultured in 250ml Erlynmeyer flasks containing 20ml liquid Semi-synthetic minimal medium at a temperature of 20oC for 120hr at 180rpm on a temperature controlled orbital shaker having 25mm stroke. Biomass was harvested by centrifugation at 8,000 g for 10 min in a cooling centrifuge. Wet weights of the cultures under study were recovered after decanting the broth.
Lipid Extraction
Accumulated lipid was extracted from the cultures under study by the modified Bligh & Dyer Method, 1959. Lipids were extracted from each culture using 20 times weight of Methanol: Chloroform (2:1 v/v) against weight of collected biomass, and by shaking in sealed conical flasks overnight at 60rpm in a shaker at 25°C [22]. Broth was then filtered through Whatmann No.1. The total extracted lipid was weighed after separating and evaporating the chloroform layer containing the lipid. The percentage of lipid accumulated was calculated against the biomass i.e. mg total extracted lipid/mg wet weights of cells [23].
Gas Chromatography
Only 3 strong lipid accumulators capable of giving reproducible amount of SCO accumulation were considered for GC analysis. Fatty acid methyl esters (FAMEs) were prepared from the obtained lipid by transmethylation with methanolic HCl in a water bath at 80oC as described by Ranjekar PK, et al. [24]. The esters were extracted in hexane and dried under nitrogen. FAME were analyzed by Shimadzu 2014 Gas Chromatograph (Shimadzu Corporation, Kyoto, Japan) equipped with Flame Ionization Detector and Rt®- 2330 (Restek Corporation, USA) capillary column (30m × 0.32mm ID × 0.2µm df). Nitrogen was used as carrier gas. Initial column temperature was set at 140oC which was later raised to 230°C at 4°C min-1 and final hold of 5min. The injector and detector were kept at 240°C with an injection volume of 0.2 µl. Chromatographic comparison with authentic FAMEs standards (Supelco™, Sigma-Aldrich®) were used for identifying the FAME from cultures. The quantities of individual fatty acids were estimated from the peak areas on the chromatogram.
GC/MS
GC-MS analysis was performed as confirmatory technique for the three selected SCO producers. FAME were analyzed on Shimadzu GCMS QP 2010 ULTRA equipped a Quadrupole Mass Spectrophotometer (Shimadzu Corporation, Kyoto, Japan). Rt®-2330 (Restek Corporation, USA) capillary column (30m × 0.32mm ID × 0.2µm df) was used for analysis. The oven temperature program was same as on GC analysis (mentioned in section 2.5). Helium was used as the carrier gas at a linear velocity of 44.4 cm/sec. The ion source temperature was maintained at 200oC, and analysis performed in the EI (Electron Impact Ionization) Scan mode at 70eV. Scans from 40 to 600 m/z were run with an event time of 0.30 seconds and scan speed of 2000 u/sec [18].
Microbial Identification
Morphological and Biochemical Characterization: The two selected isolated were studied for their morphologies, Gram characters, motility as described in the manual Microbiological Methods by Collins [25]. About 46 biochemical characteristics like sugar fermentation, MR-VP test, nitrate reduction, indole production, citrate utilization, TSI, ammonia production, amino acid metabolism and enzyme tests were studied using the VITEK®2 Microbial Identification System from bioMérieux, France [26].
Molecular Identification: The molecular technique of 16S r-RNA sequencing employing universal PCR primers, was used for molecular identification of the isolate. An isolated colony was used for DNA extraction using Phenol-Chloroform method for PCR Template preparation. Unincorporated PCR primers and dNTPs were removed from PCR products by using Montage PCR Clean up kit (Merck Millipore, USA). Further, the purified PCR product was sequenced by Cycle sequencing using Universal primers 27F AGAGTTTGATCMTGGCTCAG and 1492RTACGGYTACCTTGTTACGACTT. Sequencing reactions were performed using the ABI PRISM® BigDyeTM Terminator Cycle Sequencing Kit with AmpliTaq® DNA polymerase (FS enzyme) from Applied Biosystems Inc., USA (Applied Biosystems Inc. © 1998) [27]. To identify the closest similarity hit to our isolates the sequenced 16S rRNA regions from the isolates ranging around 800bp were analysed using the NCBI nBLAST (nr/nt) Database [28].
Results and Discussion
A total of 27 lipid producers were isolated from the fresh water (Pindhari river, Uttarakhand) and sea water (warm Arabian Sea). Screening for PUFA producers form the marine waters from Arabian Sea and fresh water from Indian rivers and lakes were encouraging. Bacterial PUFA producers are of great interest as they are cheap and reliable sources and find applications in industrial and health segments. Thus, the current study was undertaken to isolate that may prove to be superior to the existing algal and fungal PUFA producing microbial cultures. Furthermore, a bacterial fermentation- based process could provide a reliable economically attractive PUFA source.
Preliminary Screening
Sudan Black B Staining: Sudan Black B is a slightly basic dye that interacts with the acidic groups in the lipid compounds, thereby staining the lipoproteins, phospholipids, triglycerides etc., bluish black. The counter stain saffranine stains the rest of the cell pink. Completely pink cells were considered negative and discarded. Amongst the lipid accumulators, 3 isolates (namely PM2, PM5 and AlkM3 were selected for further studies, for they demonstrated intense Sudan black B-stained cells, indicative of maximum lipid build-up (Figure 1).

Percentage of Lipid Content w.r.t. Biomass: In a preliminary attempt to quantitate the lipid production the cultures were cultured in nitrogen limited liquid medium as described in the section 2.3. The percentage of lipid they could accumulate was measured against their wet biomass (Table 1). When the nitrogen gets exhausted, biomass production reduces and lipid accumulation starts [29].
Method of lipid extraction is crucial and depends on the application of the lipids, especially in foods sector. The solvents used in lipid extraction should be removable / recoverable and non-toxic. They also need to be protected from degradation by oxidation or enzymatic activities. Storage lipids (neutral fatty acids and triglycerols) are extracted with non-polar solvents like hexane and chloroform, as polarity is the key to efficient extractions. On the contrary polar solvents are used for extraction of more polar lipids like the phospholipids and glycolipids [30]. Owing to wide variations in the organisms, pre-treatment and extraction method have been adapted individually, thus several methods have been developed.
| Criteria | PM2 | PM5 | AlkM3 |
|---|---|---|---|
| Wet Biomass (mg) | 771 | 896 | 490 |
| Total Lipid (mg) | 212 | 329 | 110 |
| Percentage Lipid vs Wet Biomass | 27.5 | 36.7 | 22.5 |
| Physical Nature of Lipid | White Solid | White Solid | Brown Solid |
Table 1: Biomass and Lipid, Percentage Lipid produced by cultures under study.
Cultures namely AlkM3, PM2 and PM5 were selected as they showed a comparably higher biomass and lipid percentage as against the other cultures isolated in this study. Among these three, highest percentage i.e. 27.5% of lipid was produced by the Isolate No. PM2 obtained from the Arabian Sea. Isolate no. PM2 can be considered as oleaginous microorganisms as it can accumulate more than 20% of its biomass as lipid [4]. Around 14.4% lipid content was noted for the Isolate No. PM5 and about 3.06% lipid content for isolate No. AlkM3 isolated from the cold River Pindhari in northern India. Our results are in agreement with study by Delong and Yanos, 1986, which states that PUFA are not only found in deep sea barophilic bacteria but also in Vibrio spp isolated from relatively shallow depths. We believe PUFA production by these strains most likely represents an adaptation of low temperature rather than the high pressure. Only a few studies reported so far, have lipid producers isolated form fresh water sources. Thus, our study supports the hypothesis that fresh water samples also should be equally explored for isolation of lipid procedures. These fresh water mesophiles can be attractive PUFA producers as they can be cultures at ambient temperature without sea salts thus making the fermentation process more economical.
Gas Chromatography
SCO from these 3 shortlisted isolates was analyzed for identifying their fatty acids by lipid extraction and derivatization followed by Gas Chromatography Analysis. Identification of fatty acid methyl esters (FAMEs) derived from the hexane extracts of bacteria was based on matching GC retention times of analyses with those of FAMEs standards. Gas chromatograms of the FAMEs obtained from the 3 cultures under study can be seen in Figure 2. Also, the noteworthy fatty acids identified by GC are presented in Table 2.
| Isolate | PM2 | PM5 | AlkM3 | |||
|---|---|---|---|---|---|---|
| Fatty Acids | F.A. | % | F.A. | % | F.A. | % |
| Saturated Fatty Acids | Myristic acid (14C) | 54 | Myristic acid (14C) | 12 | Myristic acid (14C) | 30 |
| Palmitic acid (16C) | 13 | Palmitic acid (16C) | 23.8 | |||
| Stearic acid | 1 | Stearic acid (18C) | 2.6 | |||
| Monounsaturated Fatty Acids (MUFA) | (18C) | Behenic acid (20C) | 1.6 | |||
| Myristoelic acid (14C:1) | 10.7 | Palmitoleic acid (16C:1) | 15 | Palmitoleic acid (16C:1) | 3.6 | |
| Palmitoleic acid (16C:1) | 16.2 | Oleic acid (18C:1) | 7.8 | |||
| Poly-unsaturated Fatty acids (PUFA) | Oleic acid (18C:1) | 2 | ||||
| Eicosapentaenoic acid (20C:5) | 0.3 | Eicosapentaenoic acid (20C:5) | 0.2 | Linoleic acid (18C:2) | 1.7 |
Table 2: Noteworthy fatty acids identified by Gas Chromatography (GC). Key: F.A. = Fatty Acid, % = relative peak area percentage
Table 2: Noteworthy fatty acids identified by Gas Chromatography (GC). Key: F.A. = Fatty Acid, % = relative peak area percentage in GC chromatogram (%) As can be seen from Table 2 the bacteria under consideration produced 14-20 carbon saturated fatty acids (SFA). Monounsaturated fatty acids (MUFA) were also produced of which Palmitoleic acid (16:1) was seen in all of the three isolates. Another commercially and nutritionally important MUFA Oleic acid (18:1) was seen in considerable amounts in both Isolate No. PM2 (7.0%) and Isolate No.
AlkM3 (7.8%). Linoleic acid (18:2 n-6), an omega-6 PUFA, was produced by Isolate No. AlkM3. A valuable long chain polyunsaturated fatty acid 5,8,11,14,17-Eicosapentaenoic acid (20:5n-3) (EPA) was detected the two isolates (Isolate No. PM2 and PM5) from the Arabian Sea sampled from Pallande offshore, Maharashtra India.
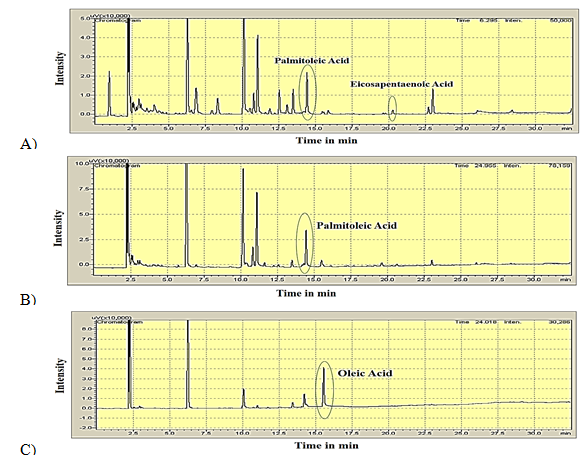
GC/MS: The results obtained from GC analysis were confirmed by GC/MS analysis of the total lipids from the isolates. Mass spectra acquired were matched with the National Institute of Standards and Technology Mass Spectral Reference Library (NIST08.LIB). The GC-MS spectrum showing the presence of Palmitoleic acid in the lipid extracted from isolate No. PM2 is seen in Figure 3.
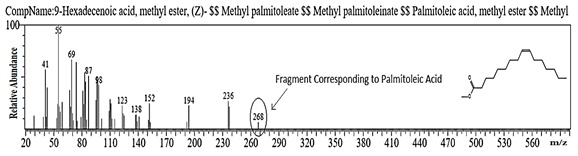
The GC/MS spectrum depicts the typical fragmentation obtained for Oleic Acid and Linoleic Acid from the extracted lipid extracted from Isolate No. AlkM3 is seen below in Figure 4. Fragments 296 and 294 corresponding to oleic acid and linoleic acid, respectively were observed in Isolate No. AlkM3.
Phenotypic Characteristics: (Biochemical and Physiological): A summary of the biochemical and molecular identification results of the cultures has been tabulated below: (Table 3).
| Isolate | Morphology | Biochemical Identification | Molecular Identification by Sequencing & NCBI BLAST Search | Percent Identity by NCBI BLAST Search | 16S rRNA Seq bp Length | NCBI Reference Sequence Accession. No. |
|---|---|---|---|---|---|---|
| Isolate No. PM2 | Gram Negative thick coccobacilli | Brevibacillus laterosporus | Achromobacter xylosoxidans strain DN002 | 98.81% | 838 | CP045222.1 |
| Isolate No. PM5 | Gram Positive filamentous bacilli | Unidentified Organism | Cobetia pacifica strain GPM2 chromosome | 99.88% | 859 | CP047970.1 |
| Isolate No. AlkM3 | Gram Positive short curved rods | Sphingomonas paucimobilis | Ornithinibacillus sp. Strain XIXJ148 16S rRNA gene partial sequence | 100% | 801 | MH801076.1 |
Table 3: Biochemical and Molecular Identification results for 3 cultures under study.
One of the theories put forward by DeLong and Yayanos, 1986, suggests that the deep sea bacterial isolates produce polyenoic fatty acids that help them adapt to the high pressure, low temperature conditions existing in the oceans. Bacterial isolates from Antarctic regions and similar low- temperature marine ecosystems demonstrated higher tendencies of lipid accumulation [31, 32, 33, 34].
Conclusion
Nowadays the main focus of single cell oil technology development is on production of long chain Polyunsaturated Fatty Acids (PUFA) with applications in human health, as nutraceuticals, pharmaceuticals and pharmaceutical precursors [3]. Bacterial PUFA producers offer a lucrative option for economical production. Also, PUFA are highly priced when obtained from natural sources and hence using bacterial systems for their commercial production may prove to be of great economic advantage. In the present study newer bacteria capable of producing PUFA were isolated from salt and fresh water sources from India. Three isolates were confirmed as potential SCO producers by GC/ MS. These screened isolates were identified by Biochemical and Molecular Characterization Techniques. It is an exciting finding that a Ornithinibacillus sp. Marseille-P3601 strain isolated in our study from the cold-water River Pindhari, Uttarakhand is found capable of producing PUFA. We are most likely the first one to show that Ornithinibacillus sp. isolated from an Indian freshwater river is capable of producing PUFA. The biochemical studies of the organisms revealed valuable information about their substrate utilization and metabolic activities. Complete identification of isolates along with their strain numbers was accomplished by 16S rRNA sequencing, a modern confirmatory identification tool. It is evident that, while the strains analyzed do not contain high levels of SCO found in some algae and fish oil, they do show valuable PUFA like EPA in their lipid, hence hold promise for future process development.
References
-
Ratledge C, Cohen Z (2008) Microbial and algal oils: do they have a future for biodiesel or as commodity oils? Lipid Technology 20(7): 155-160.
-
Meng X, Yang J, Xu X, Zhang L, Nie Q, et al. (2009) Biodiesel production from oleaginous microorganisms. Renewable energy 34(1): 1-5.
-
Ochsenreither K, Glück C, Stressler T, Fischer L, Syldatk C (2016) Production strategies and applications of microbial single cell oils. Front Microbiol 7: 1539.
-
Patel A, Karageorgou D, Rova E, Katapodis P, Rova U, et al. (2020) An overview of potential oleaginous microorganisms and their role in biodiesel and omega-3 fatty acid-based industries. Microorganisms 8(3): 434.
-
Asadi SZ, Darani KK, Nikoopour H, Bakhoda H (2015) Evaluation of the effect of process variables on the fatty acid profile of single cell oil produced by Mortierella using solid-state fermentation. Crit Rev Biotechnol 35(1): 94-102.
-
Béligon V, Christophe G, Fontanille P, Larroche C (2016) Microbial lipids as potential source to food supplements. Current Opinion in Food Science 7: 35-42.
-
Bellou S, Triantaphyllidou IE, Aggeli D, Elazzazy AM, Baeshen MN, et al. (2016) Microbial oils as food additives: recent approaches for improving microbial oil production and its polyunsaturated fatty acid content. Curr Opin Biotechnol 37: 24-35.
-
Ward OP, Singh A (2005) Omega-3/6 fatty acids: alternative sources of production. Process biochemistry 40(12): 3627-3652.
-
Shahidi F, Ambigaipalan P (2018) Omega-3 polyunsaturated fatty acids and their health benefits. Annual Rev Food Sci Technol 9: 345-381.
-
Kapoor B, Kapoor D, Gautam S, Singh R, Bhardwaj S (2021) Dietary polyunsaturated fatty acids (PUFAs): Uses and potential health benefits. Curr Nutr Rep 10(3): 232-242.
-
Hossain MA (2011) Fish as source of n-3 polyunsaturated fatty acids (PUFAs), which one is better-farmed or wild. Advance Journal of food science and technology 3(6): 455-466.
-
Jurić S, Jurić M, Siddique MAB, Fathi M (2020) Vegetable oils rich in polyunsaturated fatty acids: Nanoencapsulation methods and stability enhancement. Food Reviews International 38(1): 32-69.
-
Siriwardhana N, Kalupahana NS, Moussa NM (2012) Health benefits of n-3 polyunsaturated fatty acids: eicosapentaenoic acid and docosahexaenoic acid. Adv Food Nutr Res 65: 211-222.
-
Ratledge C (2013) Microbial oils: an introductory overview of current status and future prospects. Ocl 20(6): D602.
-
Jadhav VV, Jamle MM, Pawar PD, Devare MN, Bhadekar RK (2010) Fatty acid profiles of PUFA producing Antarctic bacteria: correlation with RAPD analysis. Annals of microbiology 60(4): 693-699.
-
Patnayak S, Sree A (2005) Screening of bacterial associates of marine sponges for single cell oil and PUFA. Lett Appl Microbiol 40(5): 358-363.
-
Guihéneuf F, Stengel DB (2013) LC-PUFA-enriched oil production by microalgae: accumulation of lipid and triacylglycerols containing n-3 LC-PUFA is triggered by nitrogen limitation and inorganic carbon availability in the marine haptophyte Pavlova lutheri. Mar Drugs 11(11): 4246-4266.
-
Masurkar AA, Datar AG, Vakil BV (2014) Screening and identification of PUFA producers from various fish of indian coastal and fresh water habitats. International Journal of Life Sciences Biotechnology and Pharma Research 3(3): 181.
-
Hartman TL (1940) The use of Sudan Black B as a bacterial fat stain. Stain technology 15(1): 23-28.
-
Masurkar AAK, Vakil BV (2015) Optimization of process variables for bacterial PUFA production by OFAT method. Int J Curr Microbiol App Sci 4(7): 160-181.
-
Masurkar AAK, Datar AG, Vakil BV (2015) New microbial sources of PUFA from the Arabian Sea and fresh water habitats of Indian Territory. International Journal of Pharmaceutical Sciences and Research 6(1): 349-360.
-
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37(8): 911-917.
-
Nichols DS, McMeekin TA (2002) Biomarker techniques to screen for bacteria that produce polyunsaturated fatty acids. J Microbiol Methods 48(2-3): 161-170.
-
Ranjekar PK, Hinge A, Hegde MV, Ghate M, Kale A, et al. (2003) Decreased antioxidant enzymes and membrane essential polyunsaturated fatty acids in schizophrenic and bipolar mood disorder patients. Psychiatry Res 121(2): 109-122.
-
Collins CH, Lyne PM, Grange JM, Falkinham JO (1984) Microbiological methods. 8th(Edn.), pp: 1-465.
-
Pincus DH (2006) Microbial identification using the bioMérieux Vitek® 2 system. Encyclopedia of Rapid Microbiological Methods, pp: 1-32.
-
Prism ABI (1998) BigDye™ Terminator Cycle Sequencing Ready Reaction Kit. Appl Biosystems, pp: 1-4.
-
Johnson M, Zaretskaya I, Raytselis Y, Merezhuk Y, McGinnis S, et al. (2008) NCBI BLAST: a better web interface. Nucleic Acids Res 36(2): W5-W9.
-
Wynn JP, Ratledge C (2005) Oils from microorganisms. Bailey’s industrial oil and fat products.
-
Byreddy AR, Gupta A, Barrow CJ, Puri M (2015) Comparison of cell disruption methods for improving lipid extraction from thraustochytrid strains. Mar Drugs 13(8): 5111-5127.
-
Allen EE, Facciotti D, Bartlett DH (1999) Monounsaturated but not polyunsaturated fatty acids are required for growth of the deep-sea bacterium Photobacterium profundum SS9 at high pressure and low temperature. Appl Environ Microbiol 65(4): 1710-1720.
-
Russell NJ, Nichols DS (1999) Polyunsaturated fatty acids in marine bacteria—a dogma rewritten. Microbiology 145(4): 767-779.
-
Elrazak AA, Ward AC, Glassey J (2013) Polyunsaturated fatty acid production by marine bacteria. Bioprocess Biosyst Eng 36(11): 1641-1652.
-
Moi IM, Leow ATC, Ali MSM, Rahman RNZRA, Salleh AB, et al. (2018) Polyunsaturated fatty acids in marine bacteria and strategies to enhance their production. Appl Microbiol Biotechnol 102(14): 5811-5826.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania