The Efficacy of Various Plant Extracts on the Eradication of Microbial Biofilm
The biofilm and its effects on humans and animals are critical topics to investigate. The first step in treating biofilm with neutral or medicinal plant extracts is to understand the mechanisms of action and how these extracts target the biofilm bacterial species. According to the most up-to-date research, medicinal plant extracts offer a wide range of biological potential, involving antioxidant, antibacterial and anti-inflammatory effects. Biofilm formation occurs in several stages involving attaches to the surface, develops an irreversible matrix, begins to form and releases the balance, and then initiates a new cycle. Quorum sensing (QS) is a mechanism through which bacteria communicate among each other through using tiny molecules. The main mechanisms by which natural products inhibit the development quorum sensing networks and biofilms are by suppressing cell adhesion, stopping the production of extracellular matrix, and reducing synthesis of variance factors. The ability of plant extracts to act as resistance modifying agents is a promising field in preventing the spread of bacterial resistance. The information in this review article was gathered from scientific articles indexed in databases such as PubMed, Scopus, ISI and Google Scholar from 2001 to 2022.
Introduction
A growing number of researchers are considering the use of novel plant extracts to target and eradicate biofilm formation. As a result of the rising incidence of antibiotic resistance against the majority of bacterial species isolated from various sources, there is a concern in clinical settings about randomly prescribing antibiotics to patients. Administering the wrong antibiotic to a patient who is suffering from a disease such as a urinary tract infection or other bacterial infections makes the bacteria more resistant and causes a mutation in the bacterial DNA. The current review focused on studies that used different plant extracts as new antibacterial agents and how these extracts targeted biofilm as well as the processes involved in biofilm formation. In recent years, more attention has been given to plant- derived products which are gaining popularity in preventing oral illness, due to adaptation of antibiotic resistance in oral microbial pathogens against synthetic antibiotics [1, 2, 3, 4]. According to the World Health Organization (WHO), up to 80% of the world’s population relies on traditional medicine for primary health care [5]. A great amount of traditional knowledge utilising the local biodiversity is held by the community or the natural ecosystem’s primary inhabitants [6]. Therapeutic approaches have used herbal plant extracts as antibacterials, and they have generated a lot of interest in the prevention of many diseases [7, 8, 9, 10].
Biofilm Formation Process
There are different stages involving in biofilm development including: (i) motility, approaching the surface, (ii) initial adhesion, (iii) irreversible maturation, (iv) dispersion and (v) propagation as illustrated in (Figure 1).
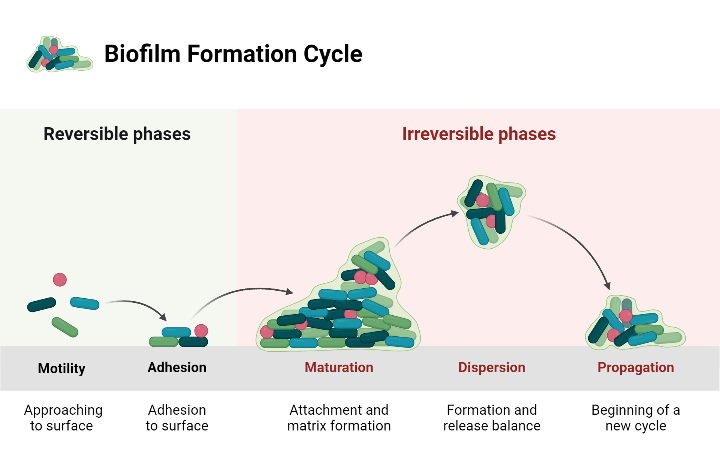
Figure 1: A diagram showing the biofilm development stages. It is clear that the bacterial biofilm moves to approach the surface in the first stage, attaches to the surface in the second stage, forms an irreversible matrix in the third stage, starts to form and releases the balance in the fourth stage, and begins a new cycle in the final phase. The diagram was generated using the BioRender.com.
Attachment
Bacterial adherence develops in aquatic environments [11]. Adherence-receptor interactions mediate bacterial adhesion on the surface of the solid-liquid [12]. Physical variables that contribute to bacterial adhesion involve covalent bonding, electrostatic interactions, and acid- base interactions [13]. In light of this, attachment seems constantly shifting and reversible, enabling certain microbes to split apart and return to their planktonic state in the event that external forces disrupt them [14].
Maturation
Bacteria frequently utilise extracellular appendages, such as flagella and type IV pili, to aid in their initial attachment [15]. The adhesive process is combined with bacterial activity along with disruptive activities during the maturation phase that follows attachment to a surface to produce intricately structured, three-dimensional water channels [16]. Depending on this intricate biofilm formation, two characteristics of bacterium biofilm have been strongly linked: elevated extracellular polymeric substance synthesis and the rise of antibiotic resistance [16]. The bacteria can now develop a microenvironment that protects them from broad spectrum antibiotics and medical plant extracts, also their antibiotic resistance can be up to 1000 times higher than that of bacteria in a planktonic condition [17]. The extracellular polymeric substance matrix is made up of a variety of components, including proteins, nucleic acids, lipids, and phospholipids, but its main component is a polysaccharide biopolymer. The outside of cells polymeric substance matrix, that also forms the bacterial diversity’s intercellular space, gives the biofilm its structure [18]. The bacterial species, environment, and conditions under which biofilms form all have a significant impact on the formation of the biofilm matrix [19].
Small molecules are used by bacteria to interact with each other as part of a process named quorum sensing [20]. The process of quorum sensing has been closely linked with the development of biofilms due to the release of small molecules called autoinducers, also referred to as pheromones, into the atmosphere around them. When there are few bacteria present, the autoinducers do not accumulate to a level that will cause gene expression. It is necessary for the number of bacteria secretes autoinducers to achieve a crucial level for either or suppression of genes that depend on quorum sensing [21]. Although the elemental composition of autoinducers varies between different microorganism species, There appear to be three fundamental groups that can be subdivided into three distinct typical classes: (i) autoinducing oligopeptides that gram-positive bacteria can recognise; (ii) acyl-homoserine lactones identified by gram- negative bacteria; and (iii) a novel signal molecule known as autoinducer-2 that acts as universal signals for both gram- positive and negative species [22, 23].
Dispersion
After sessile cells multiply to form microcolonies, planktonic cells possess a strong inclination to invade the whole surface, producing flat, smooth, uniform biofilms [24]. Physical, environmental, and extracellular secretion factors are among those responsible for the separation of biofilm. The two main categories of biofilm detachment methods are active and passive. Unlike passive detachment, which is mediated by external forces, active cell dispersion is initiated by the microorganisms themselves [24]. Both of the distinct types of biofilm dispersal mechanisms use the three stages of erosion, sloughing, and seeding in the fresh surroundings to separate the bacteria from their biofilms. Erosion, which is caused by some type of nutritional limitation, is the constant release of individual bacteria or small bacterial clusters from the top layers of a biofilm. The sudden and extensive elimination of a biofilm is known as sloughing, and it usually happens in the final phases of the creation of a biofilm. Sloughing can occur simultaneously with erosion [25]. The swarming or “seeding,” is the term used to describe the quick dispersal of significant single or clusters of bacteria within a biofilm colony. It is believed that exopolymer breakdown plays a major role in the sloughing and seeding processes in biofilms [26].
The Relationship Between Quorum Sensing and Formation of Biofilm
One of the main factors contributing to bacteria acquiring resistance to multiple drugs is the creation of biofilm. It has been noticed that QS, a cell-to-cell communication mechanism, is essential for the formation of biofilms in species that are either a gram-negative as well as positive. Many studies have been conducted on the process by which QS plays a part in the formation of biofilms. QS Enables the bacteria to sense and measure the accumulation of specific created signalling molecules produced by the community, allowing the bacteria to determine the density of the population [27]. Furthermore, at a population density that causes the amount of formed signals in the surrounding atmosphere to be induced, it modifies the expression of bacterial genes and initiates cooperative responses through the activation of signalling pathways [28]. Numerous virulence factors, including pyocyanin, proteases, elastases, and exoenzymes, are encoded by these types of genes. There is a difference between gram-positive and negative bacteria in the molecular mechanism that underlies QS, which had been thoroughly studied [29, 30, 31].
Autoinducer peptides (AIPs) were secreted by gram- positive bacteria into the environment. AIP binds to the kinase receptors on the bacteria’s membrane to send signals to the proper transcriptional components when concentrations rise, which ultimately activated the expression of related genes involving RNAIII as well as accessory gene regulator (Agr). The most prevalent gram-positive bacterium, Staphylococcus aureus, has a well-studied Agr system that plays a significant role in the synthesis of factors that promote virulence, such as degradative exoenzymes (proteases SspA, SspB, Spl, etc.) and toxins (including phenol-soluble modulins, PSMs, delta- toxin (hld), etc.) [30]. However, when the concentration of autoinducer acyl-homoserine lactones (AHLs) autoinducers in the bacterial community increased, in order to modify the expression of specific genes, the AHLs that are frequently generated in gram-negative bacteria’s interaction attach to cytoplasmic receptors. The production of virulence factors involving pyocyanin, lectin, elastase, proteases, and toxin is affected through the Luxl/luxR transcriptional factors that constitute the canonical QS system in gram-negative bacteria, these factors can be activated by AHLs. Additionally, various types of gram-negative bacteria had different autoinducers (Pseudomonas quinolone signal (PQS), CAI-1, AI-2, etc.) along with gens/QS receptors (LasI/LasR, RhlI/RhlR, CqsS and LuxPQ, etc.) [29]. On top of that, QS has been demonstrated to impact the architecture of the biofilm and offer built-in defence against outside influences like host immunity and antibiotic therapy [32].
Potential Antibacterial Agents Derived from Natural or Medicinal Plant Extracts
Nowadays, infectious diseases cause a significant number of morbidity and mortality worldwide especially with the emergence of antimicrobial resistance. New approaches to obtaining medications must be employed in place of antibiotics that, due to antibiotic resistance, demonstrate low efficacy in treating diseases in humans and animals [33]. There are (28,187) medicinal plant species have been reported having antimicrobial activity and are used as a way for treating different human and animal infectious diseases [34]. More than 1340 plants have been found to possess antimicrobial activity, as well as greater than 30,000 antimicrobial compounds were extracted from plants [35]. Medicinal plant extracts have been shown to have a variety of biological properties, including antioxidant, antibacterial, and anti-inflammatory characteristics. It has been suggested that medicinal plants’ antimicrobial compounds are a more potent means of preventing the formation of biofilms than antimicrobials that have been in use recently. Additionally, plants are a great source of antimicrobials with little adverse effects during medical care.
Recent study carried out by Blando, et al. [36] who observed how polyphenolic extracts from Opuntia ficus- indica (L.) Mill. cladodes inhibited the development of certain enterobacteria and Staphylococcus aureus ability to form biofilms. The research team examined the phenolic content and antioxidant activity, through the use of ex vivo assays for cellular antioxidant activity in red blood cells and in vitro assays for oxygen radical absorbance capacity and trolox equivalent antioxidant capacity. The activities of native plants (Larrea divaricata, Tagetes minuta, Tessaria absinthioides, Lycium chilense, and Schinus fasciculatus) as antimicrobial and anti-biofilm were reported by Romero, et al. [37]. Based on ethnobotanical knowledge from specific plants in northwest Argentina, the aforementioned five species are a potential source of alternative medicinal products made from extracts of plants [37].
The antibacterial and anti-biofilm properties of 13 medicinal plants gathered in northeastern Thailand were investigated by Saeloh and Visutthi [38]. The broth micro- dilution method was employed to evaluate the crude ethanolic extracts of these plants against Staphylococcus aureus (ATCC25923) and Escherichia coli (ATCC25922). According to their findings, the ethanolic extract from piper betle leaves significantly inhibited the growth of biofilms against the strains they tested; the reduction in growth ranged from 70–85% for S. aureus and 50–85% for E. coli. The growth, proliferation, and biofilm formation of Streptococcus mutans are inhibited by chlorella vulgaris extract, based on a study conducted by Jafari, et al. [39]. The findings indicated that the extract had a minimum bacterial concentration between 25 and 50 mg/mL. Furthermore, at a concentration of 50 mg/mL, it was effective in inhibiting biofilm formation and evaluated toxicity at a concentration of 100 mg/mL.
A different study examined the effects of cornflower extract (Centaurea cyanus) on Pseudomonas aeruginosa, Bacillus cereus, Streptococcus pneumoniae, and Staphylococcus aureus biofilms. The findings demonstrated that the type of solvent used and the extract’s concentration impacted the extract’s inhibitory effect on the biofilm structure of bacteria. The strongest inhibitory effect was observed on the development of biofilm bacteria from E. coli (84.26%) and Streptococcus pneumoniae (83.14%), respectively [40]. Based on the above-mentioned review of the literature, for many infections and diseases, plant extract is an excellent source of complementary therapies. Numerous secondary metabolites, or phytochemicals, found in plants include tannins, alkaloids, polyphenols, flavonoids, and terpenoids. These compounds have been experimentally demonstrated to have potent antibacterial effects. Recent study carried out by Wijesundara and Vasantha [41], which found that licorice root, purple coneflower flower, purple coneflower stem, sage leaves and slippery elm inner bark ethanol extract were effective on bacterial growth and biofilm formation of Streptococcus pyogenes, with minimum inhibitory concentrations (MIC) and minimum bactericidal concentrations (MBC) of 62.5 μg/ mL and 125 μg/mL, respectively.
Natural and Synthetic Substances that Influence Biofilm Formation
In vitro experiments showed that several natural compounds of plant sources had antibacterial effects. Natural products anti-biofilm impacts primarily depend on the suppression of cell adhesion, interruption of extracellular matrix creation and reduction of virulence factor synthesis, which together prevent the formation of quorum sensing networks and biofilms [42]. There are numerous compounds having antibacterial potential that are derived from plant sources, including garlic is thought to be a rich source of several antibacterial compounds, garlic extract has been demonstrated to possess inhibitory activity on QS. Tobramycin was able to inhibit Pseudomonas aeruginosa in a mouse lung infection model when the extract from garlic was added [43]. It was found that garlic extracts inhibited the growth of biofilms in six clinical bacterial isolates [44].
According to a study achieved by Kim and his co- workers, Cocculus trilobus and Coptis chinensis medicinal plant extracts can prevent bacteria from adhering to surfaces that have been coated with fibronectin. By blocking the membrane enzyme sortase from catalysing the covalent attachment of surface proteins to peptidoglycan in gram- positive bacteria, they have anti-adhesin impacts during the adherence phase of biofilm development [45]. The ethyl acetate fraction of Cocculus trilobus showed the strongest activity in preventing bacterial attachment by inhibiting the sortase enzyme out of the two plant fractions that were evaluated: water and ethyl acetate separately [45]. Ginkgo biloba extract, at 100 µg/ml, inhibited the growth of bacteria but did not affect the formation of E. coli O157:H7 biofilms on nylon, polystyrene, or glass membrane surfaces. The ways in which of inhibiting activity in E. Coli O157:H7 revealed that ginkgolic acid suppressed the prophage and curli genes, that has been linked to a reduction in the formation of biofilm and fimbriae [46, 47]. AHLs synthesised by adding additional aromatic moieties [N-(indole-3-butanoyl)-L-HSL and N-(4- bromo-phenylacetanoyl)-L-HSL] have been indicated to exhibit inhibitory effects on the LasR-based QS system and the production of biofilm in P. aeruginosa PAO1 [48]. Synthetic AHL analogues, which replace the homoserine lactone ring with a cyclohexanone ring, suppress the expression of LasI AHL synthase. This results in a decrease in the level of expression of virulence genes such as pyocyanin and elastase, as well as a change in the shape and phenotype of biofilms [49]. N-acyle cyclopentyl amides, which are non- hydrolysable cyclopentyl analogues of AHLs, prevent the development of biofilms, the expression of lasI and rhlA, along with the process for producing factors that promote virulence such as elastase, pyocyanin, and rhamnolipids, while not influencing the growth of bacteria [50].
Motivated by naturally occurring compounds generated from the marine macroalga Delisea pulchra, halogenated furanones (specifically, C-30 and C-56) demonstrate a biofilm decrease and target the P. aeruginosa las and rhl systems [51, 52, 53]. Furthermore, they have been shown to inhibit P. aeruginosa-infected the mouse lungs from colonising with bacteria, which improves the removal of bacteria from the host and lessens tissue destruction [54]. Azithromycin, a macrolide antibiotic that is produced from Saccharopolyspora erythraea, became the subject of extensive research due to its potent anti-QS and biofilm- inhibiting properties against P. aeruginosa [55, 56, 57]. It has been demonstrated that two secondary fungal metabolites from Penicillium species, penicillic acid and patulin, influence QS-controlled expression of genes in P. aeruginosa. This is most likely due to their posttranscriptional effects on the RhlR along with LasR regulatory proteins. P. aeruginosa PAO1 biofilms administered with both patulin and tobramycin had been significantly highly vulnerable towards antibiotics than control biofilms subjected to both patulin and tobramycin independently according to in vitro investigations [58].
Nevertheless, the authors did not suggest any predictions regarding how it works of action, and treatment with patulin alone had no effect on the biofilm’s growth. Patulin’s gene toxicity undoubtedly restricts its potential applications [59]. One of the main components of turmeric roots (Curcuma longa L.) is the phenolic compound curcumin, which blocks the attachment of P. aeruginosa PAO1 to polypropylene surfaces by reducing the factors associated with virulence pyocyanin, elastase, and protease. This has been associated with a reduction in the synthesis of 3-oxo-C12-HSL [60]. According to in vitro and in vivo settings, the root of Ocimum basilicum L. produces rosmarinic acid, a naturally occurring phenolic compound, in response to P. aeruginosa infection. It prevents the formation of biofilm but cannot pass through mature biofilm [61]. Virtual assessments conducted using structure against the LasR and RhlR protein receptors successfully demonstrate that rosmarinic acid could serve as a QS blocker [62]. AHLs have been experimentally demonstrated to concurrently inhibit the levels of expression of the lasIR and rhlIR genes when exposed to derivatives products of ellagic acid from Terminalia chebula Retz. This attenuates the generation of virulence factors and increases the susceptibility of biofilm for tobramycin [63]. In a V. harveyi bioassay, Girennavar, et al. demonstrated that the autoinducers AI-1 (N-3 hydroxybutanoyl- homoserine lactone) and AI-2 (furanosyl borate diester) are inhibited by the phenolic compounds bergamottin and dihydroxybergamottin from grapefruit juice. Moreover, the aforementioned researchers demonstrated that AI-1 and AI-2 prevent P. aeruginosa, Salmonella typhimurium, and E. coli O157:H7 from forming biofilms without compromising the development of bacteria.
Conclusion and Future Perspectives
A variety of natural and medicinal plant extracts were presented in this review based on their medical history and mechanisms of action. When the antibacterial effect of extracts from natural plant sources was compared to antibiotics, plant extracts had a greater impact on biofilm formation than antibiotics. This makes the possibility of developing extracts that specifically target and treat infections caused on by virulent biofilm bacterial species, opening up new potential for the development of novel treatments for biofilms. The next step is to look for new plants that have not been used in the treatment of diseases caused by biofilm bacterial species, since this could be a novel treatment.
Declarations
Competing Interests: The author declare that they have no competing interests.
Funding: This research received no external funding.
Acknowledgement
The author is grateful to University of Fallujah, College of Veterinary Medicine in Iraq – the province of Anbar for providing encouragement.
References
-
Lynch AS, Robertson GT (2008) Bacterial and fungal biofilm infections. Annu Rev Med 59: 415-428.
-
Palombo EA (2011) Traditional medicinal plant extracts and natural products with activity against oral bacteria: potential application in the prevention and treatment of oral diseases. Evid Based Complement Alternat Med 2011: 680354.
-
Nittayananta W, Limsuwan S, Srichana T, Sae-Wong C, Amnuaikit T (2018) Oral spray containing plant-derived compounds is effective against common oral pathogens. Arch Oral Biol 90: 80-85.
-
Lugo-Flores MA, Quintero-Cabello KP, Palafox-Rivera P, Silva-Espinoza BA, Cruz-Valenzuela MR, et al. (2021) Plant-derived substances with antibacterial, antioxidant, and flavoring potential to formulate oral health care products. Biomedicines 9(11): 1669.
-
Hosseinzadeh S, Jafarikukhdan A, Hosseini A, Armand R (2015) The application of medicinal plants in traditional and modern medicine: a review of Thymus vulgaris. International Journal of Clinical Medicine 6(9): 635-642.
-
Pardhan BK, Badola HK (2008) Ethnomedicinal plants use by Lepcha Tribe of Dzongu Valley bordering Khangchendzonga biosphere reserve in North Sikkim, India. J Ethnobiol Ethnomed 4: 22.
-
Lim SH, Seo JS, Yoon YJ, Kim KW, Yoo SY, et al. (2003) Effect of leaf-extract from Camellia sinensis and seed-extract from Casia tora on viability of mutans streptococci isolated from the interface between orthodontic brackets and tooth surfaces. Korean Journal of Orthodontics 33(5): 381-389.
-
Limsong J, Benjavongkulchai E, Kuvatanasuchati J (2004) Inhibitory effect of some herbal extracts on adherence of Streptococcus mutans. J Ethnopharmacol 92(2-3): 281- 289.
-
Song JH, Yang TC, Chang KW, Han SK, Yi HK, et al. (2007) In vitro effects of a fraction separated from Polygonum cuspidatum root on the viability, in suspension and biofilms, and biofilm formation of mutans streptococci. J Ethnopharmacol 112(3): 419-425.
-
Sampaio FC, Socorro MDVP, Dias CS, Costa VCO, Conde NC, et al. (2009) In vitro antimicrobial activity of Caesalpinia ferrea Martius fruits against oral pathogens. J Ethnopharmacol 124(2): 289-294.
-
Donlan RM (2002) Biofilms: microbial life on surfaces. Emerg Infect Dis 8(9): 881-890 .
-
Kostakioti M, Hadjifrangiskou M, Hultgren SJ (2013) Bacterial biofilms: development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb Perspect Med 3(4): a010306.
-
Smith M (1983) Microbial adhesion to surfaces. Berkeley RCW (Ed.), Amer Chemical Soc, 16th Street, Washington, DC, USA, pp: 1155.
-
Dunne WM (2002) Bacterial adhesion: seen any good biofilms lately? Clin Microbiol Rev 15(2): 155-166.
-
Vallet I, Diggle SP, Stacey RE, Cámara M, Ventre I, et al. (2004) Biofilm formation in Pseudomonas aeruginosa: fimbrial cup gene clusters are controlled by the transcriptional regulator MvaT. J Bacteriol 186(9): 2880- 2890.
-
O’Toole G, Kaplan HB, Kolter R (2000) Biofilm formation as microbial development. Annu Rev Microbiol 54: 49- 79.
-
Davey ME, O’toole GA (2000) Microbial biofilms: from ecology to molecular genetics. Microbiol Mol Biol Rev 64(4): 847-867.
-
Vu B, Chen M, Crawford RJ, Ivanova EP (2009) Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 14(7): 2535-2554.
-
Sutherland IW (2001) The biofilm matrix–an immobilized but dynamic microbial environment. Trends Microbiol 9(5): 222-227.
-
Rutherford ST, Bassler BL (2012) Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb Perspect Med 2(11): a012427.
-
Williams P, Winzer K, Chan WC, Camara M (2007) Look who’s talking: communication and quorum sensing in the bacterial world. Philosophical Transactions of the Royal Society B: Biological Sciences 362(1483): 1119- 1134.
-
Fuqua C, Parsek MR, Greenberg EP (2001) Regulation of gene expression by cell-to-cell communication: acyl- homoserine lactone quorum sensing. Annu Rev Genet 35: 439-468.
-
Parsek MR, Greenberg EP (2000) Acyl-homoserine lactone quorum sensing in gram-negative bacteria: a signaling mechanism involved in associations with higher organisms. Proc Natl Acad Sci USA 97(16): 8789- 8793.
-
Klausen M, Heydorn A, Ragas P, Lambertsen L, Aaes‐ Jørgensen A, et al. (2003) Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol Microbiol 48(6): 1511-1524.
-
Lappin-Scott HM, Bass C (2001) Biofilm formation: attachment, growth, and detachment of microbes from surfaces. Am J infect Control 29(4): 250-251.
-
Sauer K, Stoodley P, Goeres DM, Hall-Stoodley L, Burmølle M, et al. (2022) The biofilm life cycle: expanding the conceptual model of biofilm formation. Nature Reviews Microbiology 20(10): 608-620.
-
Abisado RG, Benomar S, Klaus JR, Dandekar AA, Chandler JR (2018) Bacterial quorum sensing and microbial community interactions. mBio 9(3): e02331-17.
-
Holm A, Vikström E (2014) Quorum sensing communication between bacteria and human cells: signals, targets, and functions. Front Plant Sci 5: 309.
-
Le KY, Otto M (2015) Quorum-sensing regulation in staphylococci—an overview. Front Microbiol 6: 1174.
-
Papenfort K, Bassler BL (2016) Quorum sensing signal– response systems in Gram-negative bacteria. Nat Rev Microbiol 14(9): 576-588.
-
Jakobsen TH, Tolker-Nielsen T, Givskov M (2017) Bacterial biofilm control by perturbation of bacterial signaling processes. Int J Mol Sci 18(9): 1970.
-
Romha G, Admasu B, Gebrekidan TH, Aleme H, Gebru G (2018) Antibacterial activities of five medicinal plants in Ethiopia against some human and animal pathogens. Evid Based Complement Alternat Med 2018: 2950758.
-
Vaou N, Stavropoulou E, Voidarou C, Tsigalou C, Bezirtzoglou E (2021) Towards advances in medicinal plant antimicrobial activity: A review study on challenges and future perspectives. Microorganisms 9(10): 2041.
-
Tajkarimi MM, Ibrahim SA, Cliver DO (2010) Antimicrobial herb and spice compounds in food. Food control 21(9): 1199-1218.
-
Blando F, Russo R, Negro C, Bellis LD, Frassinetti S (2019) Antimicrobial and antibiofilm activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. cladode polyphenolic extracts. Antioxidants (Basel) 8(5): 117.
-
Romero CM, Vivacqua CG, Abdulhamid MB, Baigori MD, Slanis AC, et al. (2016) Biofilm inhibition activity of traditional medicinal plants from Northwestern Argentina against native pathogen and environmental microorganisms. Rev Soc Bras Med Trop 49(6): 703-712.
-
Saeloh D, Visutthi M (2021) Efficacy of Thai plant extracts for antibacterial and anti-biofilm activities against pathogenic bacteria. Antibiotics (Basel) 10(12): 1470.
-
Jafari S, Mobasher M, Najafipour S, Ghasemi Y, Mobasher N, et al. (2015) Evaluation of chlorella vulgaris extract inhibitory effects on growth, proliferation and biofilm formation by streptococcus mutans and evaluation of its toxicity.
-
Mohsenipour M, Hassanshahian Z (2014) Investigating the effectiveness of Centaureacyanus extracts on planktonic growth and biofilm structures of six pathogenic bacteria. SSU_Journals 22(4): 1358-1370.
-
Wijesundara NM, Rupasinghe HPV (2019) Bactericidal and anti-biofilm activity of ethanol extracts derived from selected medicinal plants against Streptococcus pyogenes. Molecules 24(6): 1165.
-
Lu L, Hu W, Tian Z, Yuan D, Yi G, et al. (2019) Developing natural products as potential anti-biofilm agents. Chinese medicine 14: 11.
-
Bjarnsholt T, Jensen PØ, Rasmussen TB, Christophersen L, Calum H, et al. (2005) Garlic blocks quorum sensing and promotes rapid clearing of pulmonary Pseudomonas aeruginosa infections. Microbiology (Reading) 151(Pt 12): 3873-3880.
-
Persson T, Hansen TH, Rasmussen TB, Skindersø ME, Givskov M, et al. (2005) Rational design and synthesis of new quorum-sensing inhibitors derived from acylated homoserine lactones and natural products from garlic. Org Biomol Chem 3(2): 253-262.
-
Kim SW, Chang IM, Oh KB (2002) Inhibition of the bacterial surface protein anchoring transpeptidase sortase by medicinal plants. Biosci Biotechnol Biochem 66(12): 2751-2754.
-
He J, Wang S, Wu T, Cao Y, Xu X, et al. (2013) Effects of ginkgoneolic acid on the growth, acidogenicity, adherence, and biofilm of Streptococcus mutans in vitro. Folia Microbiol (Praha) 58(2): 147-153.
-
Lee JH, Kim YG, Ryu SY, Cho MH, Lee J (2014) Ginkgolic acids and Ginkgo biloba extract inhibit Escherichia coli O157: H7 and Staphylococcus aureus biofilm formation. Int J Food Microbiol 174: 47-55.
-
Geske GD, Wezeman RJ, Siegel AP, Blackwell HE (2005) Small molecule inhibitors of bacterial quorum sensing and biofilm formation. J Am Chem Soc 127(37): 12762- 12763.
-
Smith KM, Bu Y, Suga H (2003) Library screening for synthetic agonists and antagonists of a Pseudomonas aeruginosa autoinducer. Chem Biol 10(6): 563-571.
-
Ishida T, Ikeda T, Takiguchi N, Kuroda A, Ohtake H, et al. (2007) Inhibition of quorum sensing in Pseudomonas aeruginosa by N-acyl cyclopentylamides. Appl Environ Microbiol 73(10): 3183-3188.
-
Ruer S, Stender S, Filloux A, Bentzmann SD (2007) Assembly of fimbrial structures in Pseudomonas aeruginosa: functionality and specificity of chaperone- usher machineries. J Bacteriol 189(9): 3547-3555.
-
Hentzer M, Riedel K, Rasmussen TB, Heydorn A, Andersen JB, et al. (2002) Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology (Reading) 148(Pt 1): 87-102.
-
Hentzer M, Wu H, Andersen JB, Riedel K, Rasmussen TB, et al. (2003) Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J 22(15): 3803-3815.
-
Wu H, Song Z, Givskov M, Høiby N (2004) Effects of quorum-sensing on immunoglobulin G responses in a rat model of chronic lung infection with Pseudomonas aeruginosa. Microbes Infect 6(1): 34-37.
-
Skindersoe ME, Alhede M, Phipps R, Yang L, Jensen PO, et al. (2008) Effects of antibiotics on quorum sensing in Pseudomonas aeruginosa. Antimicrob Agents Chemother 52(10): 3648-3663.
-
Tateda K, Comte R, Pechere JC, Köhler T, Yamaguchi K, et al. (2001) Azithromycin inhibits quorum sensing in Pseudomonas aeruginosa. Antimicrob Agents Chemother 45(6): 1930-1933.
-
Kumar M, Rao M, Mathur T, Barman TK, Joshi V, et al. (2021) Azithromycin exhibits activity against Pseudomonas aeruginosa in chronic rat lung infection model. Front Microbiol 12: 603151.
-
Rasmussen TB, Skindersoe ME, Bjarnsholt T, Phipps RK, Christensen KB, et al. (2005) Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology (Reading) 151(Pt 5): 1325-1340.
-
Glaser N, Stopper H (2012) Patulin: Mechanism of genotoxicity. Food Chem Toxicol 50(5): 1796-1801.
-
Rudrappa T, Bais HP (2008) Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. J Agric Food Chem 56(6): 1955-1962.
-
Walker TS, Bais HP, Déziel E, Schweizer HP, Rahme LG, et al. (2004) Pseudomonas aeruginosa-plant root interactions. Pathogenicity, biofilm formation, and root exudation. Plant Physiol 134(1): 320-331.
-
Annapoorani A, Umamageswaran V, Parameswari R, Pandian SK, Ravi AV (2012) Computational discovery of putative quorum sensing inhibitors against LasR and RhlR receptor proteins of Pseudomonas aeruginosa. J Comput Aided Mol Des 26(9): 1067-1077.
-
Sarabhai S, Sharma P, Capalash N (2013) Ellagic acid derivatives from Terminalia chebula Retz. Down regulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS One 8(1): e53441.
-
Girennavar B, Cepeda ML, Soni KA, Vikram A, Jesudhasan P, et al. (2008) Grapefruit juice and its furocoumarins inhibits autoinducer signaling and biofilm formation in bacteria. Int J Food Microbiol 125(2): 204-208.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania