The Role of Bacterial Pathogens in Autoimmune Diseases: Breaking Self-Tolerance
Autoimmune diseases (ADs) arise when the immune system erroneously attacks self-tissues, often due to a breakdown in selftolerance. These conditions, influenced by genetic and environmental factors, are increasingly linked to bacterial infections as significant triggers. A healthy immune system protects the body against infections. However, when the immune system malfunctions, it attacks healthy cells, tissues, and organs of the body. This malfunction or dysfunction is called autoimmune disease and can affect any part of the body, impairing psychological function and potentially becoming fatal. This review explores how bacterial pathogens such as Helicobacter pylori, Campylobacter jejuni, and Mycobacterium tuberculosis contribute to autoimmunity through mechanisms like molecular mimicry, bystander activation, and epitope spreading. These processes provoke cross-reactive immune responses, amplify immune dysregulation, and exacerbate tissue damage. Epidemiological and experimental studies reveal strong associations between bacterial infections and diseases like Guillain-Barré syndrome, rheumatoid arthritis, and sarcoidosis. By analyzing the interactions between these pathogens and immune mechanisms, the review highlights the pivotal role of bacteria in disrupting immune tolerance and driving autoimmune disease progression. Moreover, therapeutic strategies such as antibiotics, immunomodulators, and vaccines targeting bacterial-induced autoimmunity offer promising avenues for prevention and treatment. Understanding the mechanisms of bacterial action provides valuable insights into the development of effective diagnostics, preventive measures, and therapies. This review emphasizes the need for continued research on bacterial pathogenesis in autoimmunity, particularly among genetically susceptible populations, to refine targeted clinical approaches and improve outcomes in these complex diseases.
Joseph CE¹*, Zakarriyah AA², Maidambe BB², Ojonuba GE², Ibrahim SO², Akuson SA² and Musa AD³
¹Benue State University Teaching Hospital, Nigeria ²Department of Microbiology, Ahmadu Bello University, Nigeria ³Nassarawa State University, Nigeria
Abbreviations
ADS: Autoimmune Diseases; GBS: Guillain-Barré Syndrome; MHC: Major Histocompatibility Complex; HLAs: Human Leukocyte Antigens; APCs: Activate Antigen-Presenting Cells; DCs: Dendritic Cells; CagA: cytotoxin-Associated Gene A; and Vacuolating Cytotoxin A; RA: Rheumatoid Arthritis; ITP: Immune Thrombocytopenic Purpura; RHD: Rheumatic Heart Disease; Mtb: Mycobacterium Tuberculosis; SLE: Systemic Lupus Erythematosus; IFN-γL:
Interferon-gamma; IL-17: Interleukin-17; HBHA: heparin- binding Hemagglutinin Adhesin; SAG: Superantigen; TCR: T Cell Receptor; APCs: Antigen-presenting Cells; LCMV: Lymphocytic Choriomeningitis Virus; Cx: Connexins; NOD: Non-obese Diabetic; R-EAE: Relapsing Experimental Autoimmune Encephalomyelitis; TLRs: Toll-like Receptors; PAMPs: Pathogen-Associated Molecular Patterns; Tregs: Regulatory T Cells; T1D: Type 1 Diabetes; IBD: Inflammatory Bowel Disease; MS: Multiple Sclerosis; ARF: Acute Rheumatic Fever; RHD: Rheumatic Heart Disease; RAV: Rat Autoimmune Valvulitis; OMVs: Outer Membrane Vesicles.
Introduction
Overview of Autoimmune Diseases
Autoimmune diseases occur when the body’s immune system mistakenly attacks its own cells, tissues, and organs, failing to recognize them as part of itself. Paul Ehrlich, a German immunologist, first described this phenomenon in the 20th century as Horror autotoxicus, meaning the body’s immune cells attacking itself [1]. Autoimmune diseases (ADs) involve the overactivation of specific immune cells called T cells and B cells, even when there are no infections or obvious triggers [2]. These diseases are influenced by factors such as lifestyle, environment, genetics, and hormones, although their exact causes remain unclear. Despite advances in diagnosis and treatment, there is still limited understanding of how autoimmune diseases develop and lead to health problems [3]. As of 2015, autoimmune diseases affected 19.1% of people globally, with rheumatologic disorders showing the largest increase [4, 5]. Common examples include rheumatoid arthritis (which damages joints), type 1 diabetes (which targets insulin- producing cells), and systemic lupus erythematosus (which causes widespread inflammation) [2] (Table 1).
| Disease | Affected Organ(s) | Common Symptoms |
|---|---|---|
| Rheumatoid Arthritis | Joints | Joint pain, swelling, stiffness |
| Multiple Sclerosis | Central Nervous System | Numbness, weakness, vision problems |
| Type 1 Diabetes | Pancreas | Frequent urination, excessive thirst |
| Systemic Lupus Erythematosus | Multiple organs | Fatigue, joint pain, skin rashes |
Table 1: Common Autoimmune Diseases and Their Clinical Manifestations.
Role of Self-Tolerance in Immunity
The immune system can usually tell the difference between the body’s own tissues (self) and harmful invaders like bacteria (non-self). This ability is called self-tolerance [6]. Self-tolerance is maintained through two main mechanisms:
Central Tolerance: Immature immune cells that mistakenly recognize the body’s tissues as harmful are eliminated or altered before they fully develop. This process happens in the bone marrow (for B cells) or the thymus (for T cells). Cells that strongly react to self-antigens are either destroyed or modified to prevent them from causing harm [7, 8].
Peripheral Tolerance: Some immune cells that escape central tolerance can move to other parts of the body. Peripheral tolerance ensures these cells remain inactive or are regulated to prevent damage. If this system fails, it can lead to autoimmune diseases, especially those affecting specific organs [9].
Emerging Evidence Linking Bacterial Pathogens to Autoimmunity
Recent research suggests that bacterial infections may play a significant role in triggering autoimmune diseases. Key mechanisms include bystander activation, bacterial superantigens, and molecular mimicry. Bystander Activation: Infections can activate immune cells in a way that doesn’t depend on specific antigens, leading to widespread immune activity and unintended damage to the body’s tissues [10].
Superantigens: Some bacteria produce proteins called superantigens that overstimulate immune cells, causing severe inflammation and damage. This uncontrolled activation can result in symptoms like fever, shock, or even death [11].
Molecular Mimicry: Certain bacterial proteins resemble the body’s own proteins, causing the immune system to attack both the bacteria and the body’s tissues, breaking self- tolerance [12].
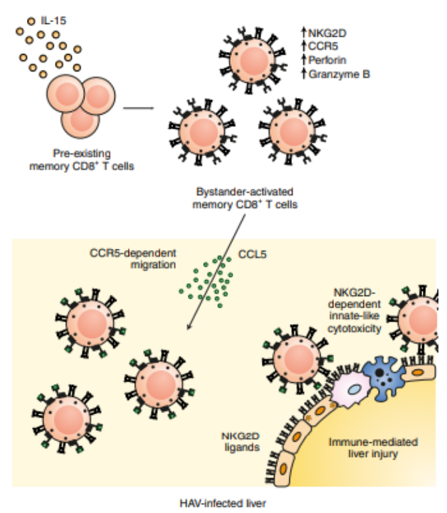
The implication of infections in the cause of autoimmune diseases, has been demonstrated by the study of animal models [13]. Epidemiological evidence indicates that Chagas’ illness is caused by a Trypanosoma cruzi infection, while rheumatic fever follows a streptococcal infection [14]. Numerous autoimmune illnesses can be triggered by an H. pylori infection [15]. Helicobacter pylori can trigger autoimmunity through prolonged interaction with the host immune system, leading to molecular mimicry and activation of cross-reactive T cells [15]. Cross-reactive T cells are activated by molecular mimicry of H. pylori antigens, which may result in autoimmune gastritis. However, there is evidence that H. pylori infection may have a protective impact in certain conditions, such as inflammatory bowel disease [16]. Also, Campylobacter jejuni, a major global cause of gastroenteritis, is known to be the causal agent of Guillain-Barré syndrome (GBS). GBS pathogenesis is significantly influenced by molecular mimicry and a cross- reactive immunological response, at least in situations where ganglioside antibodies and a prior C. jejuni infection are present [17] (Figures 1 & 2).
Mechanisms of Autoimmune Disease Development
Self-Tolerance: Mechanisms and Breakdown
The primary abnormality in ADs is the loss of self- tolerance, which makes immune cells incapable of differentiating between “self” and “non-self” antigens. This means that, aside from its poor target selection, the autoreactive immune system is working “within normal limits” to eradicate threats [18]. When the immune system mounts a defense against foreign antigens, anti-self reactions are typically produced; however, autoimmune illness only arises when autoimmunity is poorly controlled [19].
Genetic and Environmental Factors
Numerous genetic variants have been implicated in various ADs by a significant number of genome-wide association studies. The majority of the variations are found in the regulatory areas of genes whose byproducts are thought to be involved in immunological responses. According to the odds ratio, each gene has a negligible contribution to a given disease, and it is likely that several polymorphisms influence the onset of a patient’s illness [20]. Most autoimmune illnesses are caused by a combination of hereditary variables rather than being monogenic. Despite the fact that several preliminary investigations have linked human autoimmune disorders to the major histocompatibility complex (MHC), which the gene products are termed human leukocyte antigens (HLAs) in humans [21]. Families with Mendelian multi-organ autoimmune disease syndromes were found to have mutations in the gene encoding CTLA4 [22]. Environmental factors like exposure to chemicals, nutrition, and diseases can also influence immune responses resulting to increased risk of ADs, for instance, infections can lead to inflammation or trigger dormant self-reactive cells which in turn cause specific abnormalities in the gut microbiota and might change immunological homeostasis [23].
Role of Gut Microbiome in Autoimmune Diseases
The gut microbiome has recently emerged as a key player in the development of autoimmune diseases. An imbalance in this microbial community, known as dysbiosis, can lead to a cascade of inflammatory and immune-deregulatory effects [24]. Dysbiosis, often triggered by factors like environment, diet, xenobiotics, infection, and inflammation [25], can disrupt the delicate balance between the gut microbiota and the immune system. A healthy gut microbiota is essential for a robust immune response. However, dysbiosis can lead to aberrant immunological responses [26]. Beyond inflammation and oxidative stress, gut dysbiosis can also contribute to insulin resistance and dysregulated immune responses. Chronic dysbiosis and the subsequent leakage of microbiota and their metabolites can increase the risk of various health conditions, including type 2 diabetes, cardiovascular disease, autoimmune diseases, inflammatory bowel disease, and certain cancers [27].
Several mechanisms link gut microbiota imbalance to autoimmune diseases. For instance, dysbiosis can alter the host immune response, activate antigen-presenting cells (APCs) like dendritic cells (DCs), and trigger antigen presentation and cytokine production. These factors can influence T cell differentiation and function [28]. Conditions such as rheumatoid arthritis, multiple sclerosis, type 1 diabetes, and systemic lupus erythematosus have been associated with gut microbiota imbalances [29].
Molecular Mimicry: A Key Driver in Autoimmune Pathogenesis
Molecular mimicry is one of the main ways that chemical or viral agents can cause autoimmunity. It happens when foreign and self-peptide similarities favor a foreign-derived antigen’s activation of autoreactive T or B cells in a susceptible person [30]. Molecular mimicry involves a group of cross- reactions that target specific host proteins implicated in autoimmune reactions and illnesses. An autoimmune disease may therefore be triggered by molecular mimicry that is first triggered as a host reaction to a virus or microbial infection, but can also cross-react with a suitable host-antigen [31]. Numerous molecular mimicry pathways between H. pylori and the host have been suggested as a pathogen tactic to influence the host’s immune system in order to evade detection and prevent eradication [32].
Bacterial Pathogens Implicated in Autoimmunity
Helicobacter pylori and Autoimmune Gastritis
Helicobacter pylori is a spiral-shaped, Gram-negative, flagellated, microaerophilic, extracellular bacterium that resides in the human gastric submucosa. It possesses a unique ability to colonize the gastric mucosa despite the stomach’s acidic pH, facilitated by virulence factors such as the urease enzyme and flagella, which are critical for colonization [33].
H. pylori colonizes the gastric mucosa despite the stomach’s acidic pH, aided by virulence factors such as urease, flagella, and toxins like cytotoxin-associated gene A (CagA) and vacuolating cytotoxin A (VacA), which are crucial to its pathogenicity [34]. These factors are associated with various gastrointestinal conditions, including chronic gastritis, ulcers, and gastric cancers [33].
H. pylori infection triggers robust immune responses, including the recruitment of neutrophils and macrophages, leading to chronic inflammation and persistent infection [35]. By employing mechanisms like molecular mimicry, where H. pylori antigens resemble host antigens, it can provoke autoimmune responses. This results in the production of autoantibodies and inflammatory cytokines, such as TNF-α, IL-6, and IL-8 [33], contributing to systemic inflammation and immune dysregulation [33]. Specifically, CagA-positive H. pylori strains have been implicated in autoimmune gastritis. These strains induce systemic inflammatory responses that damage gastric parietal cells, impair acid secretion, and contribute to chronic atrophic gastritis. The interplay between bacterial virulence factors and host immune responses is now recognized as pivotal in the progression of both gastric and autoimmune diseases [36, 37].
Rheumatoid Arthritis: (RA) is an autoimmune, chronic inflammatory disorder that causes irreversible joint deformities and functional impairment. The exact cause of Rheumatoid Arthritis, like other connective tissue illnesses, remains unknown. Several autoimmune diseases, including rheumatoid arthritis, immune thrombocytopenic purpura (ITP), and autoimmune thyroid diseases, have been linked to pylori H [33]. Specific conditions where H. pylori play a suspected or proven role include chronic urticaria, Sjögren’s syndrome, and psoriasis [38]. H. pylori have been shown to perform a protective role, in developed countries, the prevalence of H. pylori has decreased while childhood asthma and other allergy disorders have increased. Various research has related these phenomena. According to Lim JH, et al. [39], there is an inverse relationship between H. pylori infection and asthma in young adults. The study suggests that the immune mechanism triggered by H. pylori infection may impact allergic reactions associated with asthma in this population due to its low prevalence [40]. The relationship between H. pylori and autoimmune diseases varies geographically and among populations, indicating that host and environmental factors significantly influence outcomes [34]. Similarly, Streptococcus pyogenes has been implicated in autoimmunity through molecular mimicry. Its M-proteins exhibit structural similarities with human tissues, such as cardiac myosin and valvular endothelium, leading to cross- reactive immune responses. This phenomenon underpins the pathogenesis of rheumatic heart disease (RHD), where immune- mediated valve destruction occurs. Genetic predispositions further exacerbate autoimmune outcomes in individuals, and ongoing vaccine research aims to mitigate these post-infection complications [36, 41].
Mycobacterium Tuberculosis and Autoimmune Diseases
Chronic infection with Mycobacterium tuberculosis (Mtb) has been implicated in autoimmune diseases, particularly systemic lupus erythematosus (SLE) and sarcoidosis. The relentless stimulation by mycobacterial antigens produces an acquired state of immune activation to induce self-tolerance through a cascade of events involving overly active T cells, chronic antigen presentation, and disturbed control of the immune response [42, 43]. Mtb infection evokes Th1 and Th17 pathways and both have been implicated in autoimmunity. The Th1 pathway, induced by interferon-gamma (IFN-γ) secretion, leads to inflammatory responses, while the Th17 pathway, mediated by the production of interleukin-17 (IL-17), sustains neutrophilic inflammation and preserve tissue injury [44]. Overall, these create a pro-inflammatory setting for autoimmune diseases. Granuloma formation, a defining feature of Mtb infection, shares some similarities in the pathogenesis of tissue damage of sarcoidosis [45, 46]. Similarly, the production of autoantibodies in SLE patients with latent or active tuberculosis can be explained by molecular mimicry and epitope spreading [47].
Granulomas balance the containment of latent Mtb and the risk of tissue damage. This balance depends on the dynamic interplay between M1 macrophages (pro- inflammatory) and M2 macrophages (anti-inflammatory). Immune dysfunction destabilizes this equilibrium, leading to granuloma necrosis, caseation, and increased tissue injury [48, 49]. Mtb’s survival mechanisms, such as inhibiting phagosome maturation and modulating cytokines through proteins like mammalian cell entry proteins (Mce) and heparin-binding hemagglutinin adhesin (HBHA), contribute to its persistence and immune evasion [50, 51].
The systemic dissemination of Mtb accentuates its involvement in extrapulmonary diseases and its role in chronic, multi-system conditions. Genetic differences can affect how the body responds to infections like tuberculosis and the risk of developing autoimmune diseases [52]. Recent advances in immunological profiling provide detailed insights into the interplay between chronic Mtb infection and immune-mediated pathology, emphasizing the importance of addressing latent infections in autoimmune disease management [53, 54] (Table 2).
| Bacterial Pathogen | Mechanism | Associated Conditions |
|---|---|---|
| Mycobacterium tuberculosis. | Mtb invokes both Th1 Th17 pathways, overall both creates a pro-inflammatory setting for autoimmune disease. | SLE, Sarcoidosis. |
| Helicobacter pylori. | H. pylori colonizes the gastric mucosa aided by urease, flagella, CagA and VaCA which triggers immune resonses. | Chronic gastritis, ulcers, gastric cancers. |
| Helicobacter pylori (although not a clearly known causal agent). | It is suspected that the mechanism involves infiltration of inflammatory cells into the joints, which results in destruction of bones and cartilages. | Rheumatoid arthritis, ITP, autoimmune thyroid disease, psoriasis, chronic urticarial. |
Table 2: Bacterial Pathogens, Mechanisms, and Associated Autoimmune Diseases.
Mechanisms by which Bacteria Contribute to Autoimmunity
Molecular Mimicry and Cross-Reactivity
The concept of molecular mimicry, first proposed by Raymond Damian in 1964, describes antigen sharing between pathogens and hosts. Pathogens develop structures resembling host components, aiding immune evasion. However, this resemblance can lead to autoimmunity when the immune system targets host tissues sharing structural similarities with pathogen antigens [55, 56]. Molecular mimicry initially referred to microbial mimicry of host antigens to avoid immune responses. However, it has now been expanded to include the triggering of autoimmune reactions due to structural similarities between pathogen antigens and host proteins. These similarities can provoke the production of autoreactive antibodies, leading to tissue damage, particularly when the mimicked self-determinants play essential biological roles [57, 58].
Four distinct types of mimicry have been identified:
- Sequence and structural similarity in full-length proteins or domains.
- Structural similarity without sequence homology,
- Mimicry of short protein motifs.
- Interface mimicry, where binding surface architectures resemble each other despite sequence differences [55].
Advances in bioinformatics have revealed shared epitopes between pathogens and hosts, linking mimicry to autoimmune diseases such as multiple sclerosis, type 1 diabetes, and systemic lupus erythematosus. For example, Srinivasappa showed that 3.5% of monoclonal antibodies against viruses cross-reacted with normal tissues in uninfected mice [55, 58, 59, 60, 61].
Superantigens and Polyclonal T Cell Activation
The term superantigen (SAG), introduced by White, et al. [62], describes bacterial proteins that bind the variable region of the T cell receptor (TCR) β chain and MHC-II molecules on antigen-presenting cells (APCs). This interaction causes widespread CD4+ and CD8+ T cell proliferation, triggering intense immune activation. Acute effects of SAGs include food poisoning and toxic shock syndrome, while chronic exposure has been linked to autoimmune diseases, such as multiple sclerosis, and immunodeficiencies, including those associated with HIV. SAGs disrupt immune regulation, delaying the development of pathogen-specific immunity [62, 63]. Superantigens, typically 22–29 kDa microbial proteins, can bypass conventional antigen processing to directly stimulate up to 20–30% of T cells with specific Vβ regions—10,000 times more than classical antigens. This immense activation raises concerns about their role in triggering autoimmunity by activating autoreactive T cells [64, 65]. SAGs have two domains: an NH2-terminal β-barrel for MHC-II binding and a COOH-terminal α-barrel for TCR interaction [66].
SAGs are produced by Gram-positive bacteria, such as Staphylococcus aureus and Streptococcus pyogenes, Gram- negative bacteria like Yersinia pseudotuberculosis, and wall-less bacteria like Mycoplasma arthritidis. Notable SAG families include staphylococcal enterotoxins (SEA–SEI), toxic shock syndrome toxin-1 (TSST-1), and streptococcal pyrogenic exotoxins (SPE A, C, F), along with non-pyrogenic types like staphylococcal exfoliative toxins [63, 67].
SAGs have been proposed as triggers of autoimmune diseases due to their ability to activate self-reactive T cells. In multiple sclerosis, for example, bacterial and viral peptides activate T cells specific to myelin basic protein. This persistent Vβ-specific T cell proliferation may amplify clones that cross-react with self-antigens, inducing autoimmunity even after SAG exposure ends [67, 68]. Mechanisms by which SAGs contribute to autoimmunity include:
- Aberrant Th-B cell interactions leading to autoantibody production.
- Direct activation of B cells independent of T cells.
- Sustained activation of self-reactive T cells [66]. SAGs have been implicated in autoimmune conditions like Kawasaki syndrome, diabetes, rheumatoid arthritis, and atopic dermatitis, although direct evidence of their role remains elusive [63].
Bystander Activation and Inflammatory Responses
Bystander activation refers to the indirect or nonspecific activation of autoreactive B and T lymphocytes due to inflammatory conditions during infections. The phenomenon first reported in 1996 by Tough, et al. can trigger a chain reaction where activation of one immune system arm stimulates the other, mediated by indirect signals such as co- signaling receptor ligands, cytokines, chemokines, pathogen- associated molecular patterns, and extracellular vesicles with microbial particles [61, 69]. Bystander T cells were initially identified in lymphocytic choriomeningitis virus (LCMV), vaccinia virus, and vesicular stomatitis virus murine infection models, where diverse polyclonal T cells proliferated and expanded. Bacterial products like lipopolysaccharide (LPS) have also been linked to nonspecific T cell stimulation, as demonstrated by LPS injections in mice [70].
Under normal conditions, bystander activation contributes to processes like B cell development, homeostasis, and apoptotic particle clearance. In some infectious diseases, it is crucial for counteracting microbial substances that inhibit or hijack the immune response. However, bystander activation can also lead to autoimmune diseases by triggering autoreactive lymphocytes. Evidence links this mechanism to autoimmune conditions such as type 1 diabetes, rheumatoid arthritis, autoimmune thyroid disease, multiple sclerosis, systemic lupus erythematosus, and autoimmune hepatitis [71].
Mechanisms of Bystander Activation in Bacterial Infections
Cellular Coreceptor Induction: Bacterial LPS can stimulate non-antigen-specific T cells and other immune cells like dendritic cells (DCs) by upregulating CD86 and promoting IFN-γ production. Gap Junction Communication: Bacteria use gap junctions composed of connexins (Cx) to transmit pathogen recognition signals between adjacent cells, enhancing the bystander effect. Soluble Mediator Activation: Bacteria like Legionella pneumophila and Chlamydia trachomatis activate neighboring cells via soluble mediators. Particle Exchange: Macrophages infected with Bacillus Calmette-Guérin or Mycobacterium tuberculosis release bacteria-containing exosomes that promote cytokine production in naïve macrophages [69]. Epitope Spreading and Tissue Damage Epitope spreading occurs when chronic infections lead to microbial antigens being taken up by antigen-presenting cells (APCs). These APCs present microbial antigens to T cells, initiating an inflammatory response that causes tissue damage. This damage exposes self-antigens, which are also taken up by APCs and presented to autoreactive T cells, resulting in immune attacks on the body’s tissues. This process extends the immune response from microbial antigen epitopes to non-cross-reactive self-epitopes, contributing to autoimmunity [61, 69, 72].
Epitope spreading is when the immune system expands its attack from one specific target to other similar targets in the body over time. This broadening enhances immune efficiency by optimizing antigen recognition and neutralizing antibody functions. However, in autoimmune diseases, ES varies among individuals and may follow a hierarchical structure based on factors like antigen processing, MHC restriction, and epitope-specific lymphocyte availability [73, 74].
In autoimmune conditions, ES often involves immune responses to self-antigens in previously inaccessible compartments. These hidden or cryptic epitopes become immunologically visible due to tissue damage during chronic infection or autoimmunity. Immune responses can spread intramolecularly, within the same molecule, or intermolecularly, across different molecules [71]. Although ES is a normal immune process that enhances antibacterial and antitumor responses, it becomes harmful when directed at self-antigens. ES strengthens immune defense by targeting multiple molecular structures, making it harder for pathogens to evade immunity through mutations. This mechanism also promotes persistent immunological memory and robust secondary responses, preventing immune evasion and accelerating cross-reactive antigen responses [72].
However, when ES involves self-antigens, it can bypass clonal selection mechanisms, leaving the immune system vulnerable to attacking the body. For example, in type 1 diabetes, initial autoimmunity targets the B9-23 region of insulin but spreads to other beta-cell antigens as the disease progresses. Mechanisms like endocytic processing, antigen presentation, and somatic hypermutation contribute to the broadening of immune responses in such diseases [75]. Animal models have provided valuable insights into ES in autoimmune diseases. Studies using murine relapsing experimental autoimmune encephalomyelitis (R-EAE) and non-obese diabetic (NOD) mice have shown that:
- Intra- and intermolecular epitope spreading can occur.
- ES follows a predictable cascade.
- Tissue damage is essential for initiating ES.
- ES plays a functional role in disease progression and relapse [73, 76]. Autoimmune diseases involving ES include bullous pemphigoid, systemic lupus erythematosus, multiple sclerosis, type 1 diabetes, rheumatoid arthritis, scleroderma, Graves’ disease, Sjögren’s syndrome, Heymann nephritis, cardiomyopathy, and paraneoplastic pemphigus [71, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101].
Geographic and Demographic Influences on Autoimmune Disease Risk
Autoimmune diseases show notable variability in prevalence and etiology across geographic regions and demographic groups, reflecting disparities in exposure to bacterial pathogens, environmental conditions, and genetic predispositions. For instance, Helicobacter pylori infection is significantly more prevalent in developing countries compared to developed nations. This difference stems from factors such as sanitation standards, healthcare access, and dietary habits. Interestingly, while H. pylori has been implicated in autoimmune conditions like gastritis and rheumatoid arthritis, it has also been associated with a reduced risk of allergic disorders, such as asthma, particularly in populations from developed countries. This paradoxical effect highlights the complex interplay between bacterial exposure and immune regulation [15, 39, 40].
Population-Based Studies: Epidemiological studies highlight how local factors shape autoimmune risk. A large cohort study conducted in Sweden found a 40% increased likelihood of rheumatoid arthritis development in individuals with prior Prevotella copri infections. This gut bacterium, linked to intestinal dysbiosis, is more prevalent in Western diets rich in processed foods [102, 103, 104, 105, 106, 107, 108, 109]. Conversely, in South Asia, the high prevalence of Mycobacterium tuberculosis has been linked to autoimmune conditions like sarcoidosis and systemic lupus erythematosus. The immune system’s response to chronic tuberculosis infection, including granuloma formation and persistent antigen presentation, may create a pro-inflammatory environment conducive to autoimmunity [42, 46, 110].
Meta-Analyses: Meta-analyses provide broader insights into these variations. A review of Campylobacter jejuni-associated Guillain-Barré syndrome showed a higher incidence in populations from South America and Southeast Asia. These were associated with differences in bacterial strains along with genetic susceptibility to immune-mediated nerve damage [17, 103]. Likewise, differences in the virulence factors of H. pylori, including cytotoxin-associated gene A (CagA) and vacuolating cytotoxin A (VacA), are influential determinants of the outcomes of autoimmune-related conditions. CagA-positive strains, common in East Asia, have been associated with a higher risk of autoimmune gastritis and other systemic autoimmune diseases than strains more common in Western countries [15, 33, 34] (Figure 3).
Immune Dysregulation Triggered by Bacterial Infections
Impact of Bacterial Components on the Immune System
Bacterial components such as the cell wall, membrane, and cytoplasmic structures play distinct roles in modulating immune responses. These components interact with the two major divisions of the immune system: innate and adaptive immunity [78].
Innate Immune System: The innate immune response is facilitated by the epithelial cells at mucosal surfaces and phagocytic cells in the blood, such as monocytes, granulocytes and macrophages [79]. Immune sensors recognize unique patterns on bacteria, called microbial patterns, to fight infections [80, 81]. PRRs also detect host-derived signals of damage or infection, triggering a cascade of immune responses, including the activation of cellular defenses, the release of pro-inflammatory cytokines, and recruitment of additional immune cells [82, 83]. Notably, pathogens often engage multiple PRRs simultaneously, eliciting specific signal transduction pathways that optimize the immune response to each pathogen [84, 85].
Adaptive immune System: Bacterial components can also initiate the activation of adaptive immunity, involving the production of antibodies by B cells, the cytotoxic actions of T cells and helper T cells-mediated responses. The formation of memory B and T during this process ensures long-term protection against subsequent infections with the same pathogen [86].
Role of TLRs (Toll-Like Receptors) and Innate Immune Activation
Toll-like receptors (TLRs) are key members of the PRR family, responsible for recognizing pathogen- associated molecular patterns (PAMPs) and initiating immune responses. TLRs are evolutionarily conserved and particularly effective in extracellular pathogen recognition [87, 88]. Upon activation, TLRs have been demonstrated to induce the expression of pro-inflammatory cytokines and mechanisms that bridge innate and adaptive immunity, facilitating pathogen elimination and immunological memory formation [89]. Excessive TLR activation, however, can result in systemic inflammation and severe disease, as seen in cases of bacterial sepsis or chronic infections [90].
Disruption of Regulatory T Cells (Tregs) and Immune Homeostasis
Regulatory T Cells (Tregs): Regulatory T cells (Tregs) help keep the immune system in check by stopping other immune cells from overreacting [91]. Disruption of Tregs function can lead to a loss of immune homeostasis, manifesting as autoimmune diseases such as: Type 1 diabetes (T1D): characterized by the infiltration into pancreatic islets leads to β-cell destruction and loss of insulin production [92].
Rheumatoid arthritis (RA): RA is characterized by chronic inflammation and polyarthritis, often involving controversial alterations in Treg populations that fail to control the immune response [93, 94, 95, 96].
Inflammatory bowel disease (IBD): IBD is a genetically predisposing, chronic disease with breakdown of intestinal immune homeostasis, showing an exaggerated inflammatory response to the gut microbiota [97].
Immune Homeostasis: Immune homeostasis relies on a delicate balance between activation and suppression, mediated by cellular players and chemical factors [98]. Treg disruption shifts this balance, leading to unrestrained immune activation and chronic inflammation.
Clinical Evidence Supporting Bacteria- Induced Autoimmunity
Conventionally, bacterial pathogens have been considered to induce autoimmune diseases by breaching mechanisms of immune self-tolerance. Clinical evidence to support this is reviewed in the next section which, for convenience, is grouped into three categories: case reports combined with epidemiological findings, animal model data and studies in humans which relate bacterial infection to autoimmune disease development.
Case Studies and Epidemiological Data
Case reports and epidemiological studies link bacterial infection with the onset of autoimmune diseases. For instance, Streptococcus pyogenes infection has been steadily associated with rheumatic fever, a model of autoimmune disease in which cross-reactivity between bacterial M proteins and cardiac myosin epitopes leads to immune-mediated cardiac tissue damage [99, 100]. Along this line, Helicobacter pylori infection is associated with autoimmune gastritis, in which antigenic mimicry between antigens of H. pylori and gastric epithelium provoke self-directed immune responses [101]. Epidemiological studies confirm such findings. Indeed, a large-scale cohort study conducted in Sweden in 2020 showed that a history of infection with Prevotella copri, a gut bacterium implicated in intestinal dysbiosis, was associated with an increased subsequent RA development 40% by Rodríguez D, et al. [102]. Other population-based studies of postinfectious autoimmune syndromes show increased incidences of Guillain-Barré syndrome after Campylobacter jejuni infections [103]. Thus, these studies highlight the theory that bacterial pathogens may induce autoimmunity in general across different populations.
Case Studies and Clinical Data: Real-World Implications
Case Studies: Helicobacter pylori and Autoimmune Gastritis: Autoimmune gastritis, a chronic inflammatory condition characterized by the immune-mediated destruction of gastric parietal cells, has been extensively linked to Helicobacter pylori infection. A notable case study conducted in Japan reported on a 45-year-old woman diagnosed with autoimmune gastritis following a prolonged H. pylori infection. Serological tests revealed high titers of autoantibodies against the H+/K+ ATPase of gastric parietal cells. The study demonstrated that eradication therapy for H. pylori led to a significant reduction in autoantibody levels and an improvement in gastric histology, illustrating a potential therapeutic avenue [33, 36].
In a broader cohort, A study conducted in 2022 examined 250 patients with autoimmune gastritis and found that 65% had concurrent H. pylori infections [36]. The study highlighted molecular mimicry as a driving factor, where H. pylori antigens mimicked host proteins, triggering autoimmunity. Patients undergoing H. pylori eradication therapy exhibited delayed progression of autoimmune gastritis, further supporting the pathogen’s role in disease onset and management [37].
Clinical Data: Campylobacter jejuni and Guillain- Barré Syndrome
Guillain-Barré Syndrome (GBS), an acute autoimmune neuropathy, is a well-documented consequence of Campylobacter jejuni infections. A multicenter study conducted across Europe analyzed 1,100 cases of GBS, finding that approximately 40% of patients reported recent C. jejuni infections. Serological testing confirmed the presence of ganglioside antibodies, indicating a cross-reactive immune response initiated by molecular mimicry between bacterial lipooligosaccharides and human gangliosides [17, 103]. Similarly, a meta-analysis of 35 studies demonstrated that C. jejuni-associated GBS cases exhibited more severe clinical outcomes, including prolonged recovery times and higher rates of mechanical ventilation dependency. These findings underscore the critical need for early diagnosis and intervention in populations at risk [17, 103].
Animal Models of Bacteria-Associated Autoimmune Diseases
Animal models are robust experimental systems that provide insight into the mechanisms by which bacterial pathogens trigger autoimmune diseases. For instance, in a mouse model of autoimmune arthritis, oral administration of Prevotella copri was shown to exacerbate joint inflammation via activation of Th17-driven pathways and thus demonstrated its pathogenic contribution to the development of rheumatoid arthritis [104]. Along the same line, a mouse model infected with Yersinia enterocolitica suggested that the LPS induced autoimmune thyroiditis in the infected mice through TLR-dependent inflammatory pathway [105]. It has also been demonstrated that the model of NOD-a model for Type 1 Diabetes-accelerated pancreatic β-cell destruction via colonization with B. fragilis was driven by cross-reactive T cell responses against bacterial antigens [106]. These models further strengthen the causative role played by bacterial infection in the induction and propagation of autoimmunity through mechanisms such as molecular mimicry, epitope spreading, and bystander activation.
Human Studies: Correlation between Infection and Autoimmune Risk
Human studies confirm evidence that bacterial infection predisposes one to autoimmune disease. A 2021 meta- analysis involving 25 studies demonstrated a positive correlation existing between the infection with Chlamydia pneumoniae and an increased risk for multiple sclerosis (MS) [107]. Furthermore, clinically relevant studies identified a key role that is played by periodontal pathogens such as Porphyromonas gingivalis in RA, hypothesizing that there might be a driving of autoantibody production by citrullination of bacterial proteins [108].
The larger part of the evidence comes from longitudinal studies, especially those following patients after infection. One such study published in 2022 by the UK Biobank showed a higher incidence of SLE in patients who had previously suffered from recurrent urinary tract infections due to Escherichia coli infection; the study consequently hinted that prolonged exposure to bacteria might be a major risk factor [109]. Furthermore, high-throughput sequencing studies have reported the presence of bacterial DNA on autoimmune lesion sites; for example, Mycobacterium tuberculosis DNA in sarcoidosis granulomas supports that bacterial presence is linked to localized autoimmune pathology directly [110].
Therapeutic Approaches and Interventions
Autoimmune diseases arise when the immune system mistakenly attacks the body’s own tissues, often triggered by infections. Bacterial pathogens are increasingly implicated in initiating autoimmune responses by disrupting self- tolerance as reported by Alam Y, et al. [111]. This review explores key therapeutic strategies, including antibiotics, immunomodulatory treatments, and vaccination, aimed at managing or preventing infection-driven autoimmunity.
Antibiotics in Managing Bacteria-Triggered Autoimmune Conditions
Antibiotics are commonly considered for autoimmune diseases associated with bacterial pathogens. Rheumatoid Arthritis (RA) is one such condition where bacterial involvement has been observed, potentially contributing to disease pathogenesis. Minocycline and Doxycycline: Tetracycline antibiotics, especially minocycline and doxycycline, have shown effectiveness in RA by modulating immune responses [112].
Minocycline, for instance, exhibits both antibacterial and anti-inflammatory properties, which may help in reducing RA activity. Stone, et al. [113] noted significant improvements in RA symptoms with minocycline therapy. Additionally, a randomized controlled trial by Dell R, et al. [114] found that minocycline significantly reduced disease severity in RA patients compared to placebo. Another study by Dell R, et al. [115] demonstrated that doxycycline, when used with methotrexate, provided enhanced therapeutic outcomes for RA patients, though benefits were more apparent in early disease stages.
Macrolides: Macrolides which are a group of antibiotics that contain a distinctive macrocyclic lactone ring combined with sugars (clandinose, desosamine) like clarithromycin and roxithromycin have also been explored for inflammatory diseases like RA as stated by Kwiatkowska B and Maślińska M [116]. Clarithromycin was shown by Ogrendik M [117] to reduce disease activity in RA patients. Similarly, roxithromycin improved early RA outcomes compared with placebo, as reported in a randomized, double-blind study by Ogrendik M and Karagoz N [118].
Levoflaxacin: which is an oral fluoroquinolone antibacterial agent, is the optical s-(-) isomer of ofloxacin as discussed in detail by Wimer SM, et al. [119]. An experiment demonstrated by Ogrendik M [120] in patients with active rheumatoid arthritis undergoing methotrexate therapy in addition to levofloxacin significantly improved the signs and symptoms of rheumatoid arthritis, suggesting its potential role in improving outcomes in such cases.
Immunomodulatory Treatments Targeting Infection-Driven Autoimmunity
Immunomodulatory therapies are crucial in conditions where bacterial infections are thought to influence autoimmunity. TNF Inhibitors: TNF inhibitors, such as infliximab and adalimumab, have transformed autoimmune disease management by targeting TNF-α, a cytokine involved in inflammation [121, 122] found that TNF inhibitors significantly improve disease activity in RA and Crohn’s disease, which may involve bacterial triggers. Tanaka Y, et al. [123] reported sustained remission in RA patients treated with infliximab, underscoring the efficacy of TNF blockade.
B-Cell Depletion Therapy: Rituximab, targeting CD20- positive B cells, has shown efficacy in RA and systemic lupus erythematosus (SLE) [124]. Bag-Ozbek A, et al. [125] reported remission in patients with SLE. Similarly, Korhonen R, et al. [126] suggested that rituximab had a beneficial effect in the treatment of RA in combination with cyclophosphamide. Schioppo T, et al. [127] demonstrated similar benefits in patients with RA, SLE and other autoimmune diseases. Toll- Like Receptor (TLR) Modulators: Overactivation of TLRs by bacterial components can lead to autoimmunity. HMGB1 and TLRs are crucial biomarkers, with ongoing clinical trials investigating drugs aimed at these targets, while targeting HMGB1 and TLRs has shown therapeutic potential, various limitations and challenges remain [128].
Vaccination Strategies and Prevention of Pathogen-Induced Autoimmunity
Vaccination aims to prevent bacterial infections that could trigger autoimmune diseases, thereby supporting immune tolerance, as current treatments primarily depend on broad immunosuppression which increases patients’ susceptibility to infections and requires lifelong therapy [129].
Inverse Vaccine: Inverse vaccines represent an antigen- specific method to promote tolerance to a particular antigen. Unlike traditional vaccines, which train the immune system to identify and attack pathogens such as viruses or bacteria, inverse vaccines function therapeutically by teaching the immune system to recognize certain antigens as harmless, preventing it from targeting the body’s own tissues [130]. The innovation of inverse vaccines lies in their ability to specifically target and erase the immune system’s memory of self-antigens that it erroneously attacks in autoimmune diseases. Instead of broadly suppressing the immune system, these vaccines leverage the liver’s natural mechanisms for promoting immune tolerance [131]. Recent developments in immune tolerance and therapies for multiple sclerosis (MS) have focused on innovative strategies to regulate the immune response. Early studies, such as those by Warren and Catz, explored the administration of Myelin Basic Protein 8298 (MBP 8298) without adjuvants to train the immune system to ignore brain proteins. Building on this, Roland Martin, in collaboration with Stephen Miller, introduced artificial peripheral blood leukocytes bound to multiple myelin peptides, aiming to regulate T cell activity. Opexa Therapeutics also contributed by developing patient-specific vaccines targeting malignant T cells. In a breakthrough, Steinman demonstrated that a plasmid cocktail of four myelin proteins (MBP, PLP, MOG, and myelin-associated glycoprotein) significantly reduced relapse severity in MS mouse models [132]. More recently, advanced approaches have focused on leveraging peripheral immune tolerance in the liver by tagging antigens with N-acetylgalactosamine (pGal), showing promise in reversing autoimmune conditions like MS. Additionally, research into advanced delivery systems, such as PLGA microspheres and liposomes, aims to enhance antigen stability and release. However, challenges remain in identifying autoantigens, ensuring specificity, and managing the long timelines and costs associated with clinical trials, which continue to delay the commercialization of these therapies [131].
Streptococcal vaccines: The development of a vaccine for Streptococcus pyogenes, the bacterium responsible for rheumatic fever, has been a major area of research due to the potential for widespread protection against related autoimmune diseases [133]. Despite extensive research, no commercial Streptococcus pyogenes vaccine has been developed due to risks like autoimmune responses and inflammation with live-attenuated or inactivated bacteria. Peptide-based subunit vaccines, targeting M proteins and non-M protein factors, have shown promise in preclinical and clinical trials. Examples such as StreptAnova and MJ8VAX demonstrated safety and immunogenicity in animal models and Phase I trials [134]. S. pyogenes vaccines are categorized into M-protein vaccines, which focus on conserved M protein regions, and non-M-protein vaccines, targeting other common virulence factors [135]. Another promising vaccine candidate, the StreptInCor vaccine targets S. pyogenes using a 55-amino-acid structure and has shown strong immune responses across various HLA class II types without causing adverse effects in multiple animal models, indicating its potential as a universal and safe vaccine [136, 137]. Safety remains a critical concern for these vaccines due to the potential for molecular mimicry, where S. pyogenes antigens cross-react with human tissues and lead to autoimmune conditions like Acute Rheumatic Fever (ARF) and Rheumatic Heart Disease (RHD). For example, the VaxiStrep vaccine, which uses the SpeAB fusion protein (containing inactivated forms of SpeA and SpeB), has been shown to be safe and effective in preclinical studies, with no adverse effects observed in mice and rabbits [131]. In terms of experimental models, a unique rat autoimmune valvulitis (RAV) model was used to assess the potential for autoimmune responses in vaccine trials. This model, which mimics both cardiac and neurobehavioral pathology following S. pyogenes infection, demonstrated the safety of synthetic peptide-based vaccines, such as those incorporating the p*17 peptide from the M protein and the K4S2 peptide from SpyCEP, which targets virulence factors involved in immune evasion [138]. Vaccine trials for S. pyogenes have a long history, with more than 200,000 participants involved in over 100 years of research. In the 1960s, safety concerns regarding the association between vaccination and ARF led to regulatory restrictions on the use of S. pyogenes antigens in vaccines. However, these restrictions were lifted in 2005, and since then, multiple clinical trials have been conducted using M-protein- based vaccines, such as the Hexavalent, StreptAvax, and MJ8VAX candidates. Despite the ongoing challenges, recent advancements in vaccine development continue to show promise in providing an effective, safe solution for preventing S. pyogenes infections and related autoimmune diseases [139].
Helicobacter Pylori Vaccine: Developing an effective vaccine for Helicobacter pylori (H. pylori) remains a significant challenge due to factors such as genetic diversity, immune tolerance mechanisms, and the bacterium’s ability to hide within cells. Despite being linked to autoimmune gastritis and conditions like gastric cancer, current vaccines targeting H. pylori have not advanced beyond early clinical trials [140]. Multiple vaccine approaches, including whole bacterial, subunit, and live vector vaccines, are being explored, focusing on antigens like urease, neutrophil-activating protein, and adhesion proteins such as BabA and OipA. While promising in animal models, these vaccines have not been successful in clinical settings, with issues around adjuvant efficacy and safety hindering progress [141]. Multi-epitope vaccines, which target multiple antigens, hold potential for better immunogenicity, though challenges like the large molecular size of H. pylori proteins persist. Epitope-based vaccines and advancements in immunoinformatics, including reverse vaccinology, are promising strategies for overcoming these obstacles and enhancing vaccine development [142]. In addition, new techniques like the ANTIGENome approach have identified novel antigens, such as HP1341, which could help improve immunogenicity and expand the pool of vaccine candidates [143]. To formulate an effective vaccine, key issues like selecting an optimal antigen and adjuvant need to be addressed [144]. While some adjuvants, such as aluminum hydroxide and LT, have been widely used, safer and more effective alternatives, including α-GalCer, cGAMP, and outer membrane vesicles (OMVs), are being explored. Nanomaterial-based delivery systems offer significant advantages in terms of antigen stability and prolonged immune response, though improvements in stability and controlled release are still necessary. Furthermore, the development of vaccines suitable for children, considering factors like oral dosage and alternative administration routes, remains an important focus for future research [140].
Preventive Vaccination in Susceptible Populations: The identification of individuals with genetic susceptibility, such as specific Human Leukocyte Antigen (HLA) types, and targeting them with tailored bacterial vaccines could help prevent autoimmune responses. Vadalà M, et al. [145] proposed this strategy as a potential means of reducing the risk of autoimmune diseases in genetically predisposed populations. However, they also emphasized the need for further research to refine these approaches and better understand how genetic factors influence immune responses to vaccines [146] (Table 2).
| Treatment | Mechanism | Advantages | Disadvantages |
|---|---|---|---|
| Immunosuppressants | Reduces immune activity | Effective in early stages | Risk of infections |
| Biologics | Targets specific cytokines | Fewer side effects | Expensive |
| Lifestyle Interventions | Diet, exercise, stress management | Low cost, holistic | Requires compliance |
Challenges and Future Directions
Limitations in Current Understanding
The association between bacterial pathogens and autoimmune diseases remains a complex and partially understood field. Although several mechanisms are implicated, including molecular mimicry, bystander activation, and superantigen-induced immune responses, variability in pathogen behavior and immune system responses makes a unified understanding difficult. For instance, while molecular mimicry has been implicated in conditions such as Guillain-Barré syndrome (often triggered by Campylobacter jejuni), researchers have found that not all bacterial infections result in autoimmunity, suggesting that additional factors, such as genetic predisposition and environmental exposures, play a substantial role [147, 148]. Additionally, research techniques face limitations: traditional serology, though valuable, can lack the specificity to detect low-abundance autoantibodies reliably. More recent methods like high-throughput sequencing and proteomic profiling have enhanced our understanding but are still costly and not universally accessible. As such, understanding how bacterial infections initiate autoimmunity, particularly across different diseases, remains limited by these methodological and knowledge-based barriers [149].
The Need for More Research on Pathogen- Associated Autoimmune Mechanisms
Further studies on mechanisms of pathogen-driven autoimmunity are necessary to clearly target those immune responses that might be provoked by bacterial pathogens, including streptococcal and Mycoplasma species, implicated in MS and RA, respectively, which exhibit molecular mimicry with self-antigens but whose exact cascades of immune events remain unknown. Recent research suggests that infections may trigger immune responses that evolve into chronic autoimmunity via epitope spreading or immune cell reprogramming [150, 151]. For example, Mycobacterium tuberculosis may interact with host immune cells in ways that alter immune tolerance thresholds, though studies are still needed to solidify these pathways [152]. Such mechanisms might be further unraveled with the help of further research which may involve induced pluripotent stem cells and organ- on-chip systems. Besides, more epidemiological studies concerning the pattern of infections among genetically predisposed individuals may further point out new bacterial associations that can lead towards pathogen-specific markers for diagnosis and prevention in the future [153].
Potential for Personalized Medicine in Treating Autoimmune Diseases Linked to Infections
The bridge between pathogens and autoimmunity opens new vistas for personalized medicine. Recent advances in genomic and proteomic analyses, combined with microbiome profiling, currently allow for a stratification of patients regarding their susceptibility to pathogen-induced autoimmunity. With this knowledge regarding genetic predisposition and the composition of a patient’s microbiome, it may well be feasible for the clinician to predict, in a not- so-distant future, the likelihood of autoimmune responses against certain pathogens and to intervene accordingly. For example, patients identified with a genetic susceptibility to Campylobacter-triggered autoimmunity could be monitored for early immune response changes or receive preventive treatments following exposure [147, 154].
Moreover, targeted immunotherapies and microbiome modulation therapies are being explored as ways to prevent and treat autoimmune diseases with known pathogen associations. For instance, manipulating the gut microbiome to promote beneficial bacteria could reduce pathogen colonization, potentially mitigating autoimmune risk in genetically susceptible individuals. This approach could offer a pathway for prevention as well as personalized treatment, thereby reducing reliance on generalized immunosuppressive therapies that can compromise immune defense against infections [149].
Conclusion
Summary of the Role of Bacterial Pathogens in Autoimmune Disease Development
Recent evidence implicates bacterial pathogens as key contributors to autoimmune disease onset through mechanisms that disrupt immune tolerance, such as molecular mimicry, bystander activation, and epitope spreading. Molecular mimicry occurs when immune responses to bacterial antigens inadvertently target structurally similar self-antigens, initiating autoimmunity [155, 156]. Studies have shown that bystander activation, where immune cells are non-specifically activated in pathogen presence, and epitope spreading, wherein immune responses extend to self-antigens post-infection, also play critical roles in autoimmune progression [157, 158]. These findings highlight a complex interplay between bacterial pathogens and autoimmunity, reinforcing their significant, albeit indirect, role in disease development.
Implications for Diagnosis, Treatment, and Prevention of Autoimmunity Recognizing bacterial involvement in autoimmune diseases has notable clinical implications. Diagnostically, understanding pathogen triggers linked to specific autoimmune conditions could enhance biomarkers for early detection [159, 160]. For treatment, targeting pathogen-induced immune dysregulation may provide novel therapeutic approaches, potentially reducing autoimmunity severity [161]. Interventions such as targeted antimicrobials or probiotics could help modulate immune responses and restore tolerance in susceptible individuals. Furthermore, preventive strategies like vaccination may reduce autoimmune risk among genetically predisposed populations. Continued research on specific bacterial strains and host interactions is essential for advancing diagnostic and therapeutic strategies in autoimmune diseases.
Conflicts of Interest
The authors declare no conflict of interest.
References
-
Koppala SN, Guruprasad V (2023) Overview of Autoimmunity: Classification, Disease Mechanisms, and Etiology. Turkish Journal of Immunology 11(3): 93-105.
-
Anne D, Diamond B (2001) Autoimmune diseases. New Engl J Med 345(5): 340-350.
-
Lifeng W, Wang FS, Gershwin ME (2015) Human autoimmune diseases: a comprehensive update. Journal of Internal Medicine 278(4): 369-395.
-
Lerner A, Jeremias P, Matthias T (2015) The world incidence and prevalence of autoimmune diseases is increasing. Int J Celiac Dis 3(4): 151-155.
-
Frederick WM (2023) The increasing prevalence of autoimmunity and autoimmune diseases: an urgent call to action for improved understanding, diagnosis, treatment, and prevention. Current Opinion in Immunology 80: 102266.
-
Herman W (2010) Tolerance: an overview and perspectives. Nature Reviews Nephrology 6(10): 569- 576.
-
Zhang P, Lu Q (2018) Genetic and epigenetic influences on the loss of tolerance in autoimmunity. Cellular & Molecular Immunology 15(6): 575-585.
-
Mezgebu E, Aliy A, Worku T (2023) Autoimmunity and immune tolerance: A review. Microbiology Research International 11(2): 23-35.
-
Bretscher PA (2022) An integrated view of immunological tolerance. Scandinavian Journal of Immunology 96(3): e13207.
-
Pacheco Y, Acosta-Ampudia Y, Monsalve DM, Chang C, Gershwin ME, et al. (2019) Bystander activation and autoimmunity. J Autoimmun 103: 102301.
-
Proft T, Fraser JD (2008) The bacterial superantigen and superantigen-like proteins. Immunol Rev 225: 226-243.
-
Cusick MF, Libbey JE, Fujinami RS (2012) Molecular mimicry as a mechanism of autoimmune disease. Clinical Reviews in Allergy & Immunology 42(1): 102-111.
-
Jean-François B (2005) Infections and autoimmune diseases. J Autoimmun 25: 74-80.
-
Rose NR (1998) The role of infection in the pathogenesis of autoimmune disease. Semin Immunol 10(1): 5-13.
-
Wang L, Cao ZM, Zhang LL, Dai XC, Liu ZJ, et al. (2022) Helicobacter pylori and autoimmune diseases: involving multiple systems. Frontiers in Immunology 13: 833424.
-
Hasni S, Ippolito A, Illei GG (2011) Helicobacter pylori and autoimmune diseases. Oral Diseases 17(7): 621-627.
-
Nyati KK, Nyati R (2013) Role of Campylobacter jejuni infection in the pathogenesis of Guillain‐Barre syndrome: An update. Biomed Res Int 2013: 852195.
-
Bolon B (2012) Cellular and molecular mechanisms of autoimmune disease. Toxicologic Pathology 40(2): 216- 229.
-
Fairweather D (2007) Autoimmune disease: mechanisms. Els.
-
Rosenblum MD, Remedios KA, Abbas AK (2015) Mechanisms of human autoimmunity. J Clin Invest 125(6): 2228-2233.
-
Wang L, Wang FS, Gershwin ME (2015) Human autoimmune diseases: a comprehensive update. Journal of Internal Medicine 278(4): 369-395.
-
Marson A, Housley WJ, Hafler DA (2015) Genetic basis of autoimmunity. J Clin Invest 125(6): 2234-2241.
-
Vojdani A (2014) A potential link between environmental triggers and autoimmunity. Autoimmune Diseases 2014(1): 1-18.
-
Shaheen WA, Quraishi MN, Iqbal TH (2022) Gut microbiome and autoimmune disorders. Clinical and Experimental Immunology 209(2): 161-174.
-
Levy M, Kolodziejczyk AA, Thaiss CA, Elinav M (2017) Dysbiosis and the immune system. Nature Reviews Immunology 17(4): 219-232.
-
Round JL, Mazmanian SK (2009) The gut microbiota shapes intestinal immune responses during health and disease. Nature reviews immunology 9(5): 313-323.
-
Yoo JY, Groer M, Dutra SVO, Sarkar A, McSkimming DI (2020) Gut microbiota and immune system interactions. Microorganisms 8(10): 1587.
-
Xu H, Liu M, Cao J, Li X, Fan D, et al. (2019) The dynamic interplay between the gut microbiota and autoimmune diseases. Journal of immunology research 2019(1): 1-14.
-
Oliveira GLVD, Leite AZ, Higuchi BS, Gonzaga MI, Mariano VS (2017) Intestinal dysbiosis and probiotic applications in autoimmune diseases. Immunology 152(1): 1-12.
-
Rojas M, Restrepo-Jiménez P, Monsalve DM, Pacheco Y, Acosta-Ampudia Y, et al. (2018) Molecular mimicry and autoimmunity. J Autoimmun 95: 100-123.
-
Oldstone MBA (2014) Molecular mimicry: its evolution from concept to mechanism as a cause of autoimmune diseases. Monoclonal Antibodies in Immunodiagnosis and Immunotherapy 33(3): 158-165.
-
Chmiela M, Gonciarz W (2017) Molecular mimicry in Helicobacter pylori infections. World Journal of Gastroenterology 23(22): 3964-3974.
-
Yula E, Köksal F (2016) Autoimmune extraintestinal manifestations of Helicobacter pylori infection: A bundle of conflicts. Journal of Immunology and Clinical Microbiology 1(1): 22-30.
-
Yula E, Nagiyev T, Kaya OA, İnci M, Çelik MM, et al. (2013) Detection of primary clarithromycin resistance of Helicobacter pylori and association between cagA+ status and clinical outcome. Folia Microbiologica 58(2): 141-146.
-
Papamichael K, Konstantopoulos P, Mantzaris GJ (2014) Helicobacter pylori infection and inflammatory bowel disease: Is there a link?. World JGastroenterol 20(21): 6374-6385.
-
Tonegato M, Panozzo MP, Antico A, Bizzaro N (2024) Improving the Diagnosis of Autoimmune Gastritis: From Parietal Cell Antibodies to H+/K+ ATPase Antibodies. Diagnostics (Basel) 14(16): 1721.
-
Cunningham MW (2014) Rheumatic fever, autoimmunity, and molecular mimicry: the streptococcal connection. International Review of Immunology 37(4): 314-329.
-
Campanati A, Ganzetti G, Martina E, Giannoni M, Gesuita R, et al. (2015) Helicobacter pylori infection in psoriasis: results of a clinical study and review of the literature. International Journal of Dermatology 54(5): e109-e114.
-
Lim JH, Kim N, Lim SH, Kwon JW, Shin CM, et al. (2016) Inverse Relationship between Helicobacter pylori Infection and Asthma among Adults Younger than 40 Years. Medicine 95(8): e2609.
-
Ram M, Barzilai O, Shapira Y, Anaya JM, Tincani A, et al. (2013) Helicobacter pylori serology in autoimmune diseases - fact or fiction?. Clinical Chemistry and Laboratory Medicine 51(5): 1075-1082.
-
Schubert-Unkmeir F (2023) Group A streptococcal M proteins: From molecular mimicry to targeted vaccine development. Infection and Immunity 90(4): 421-422.
-
Cambier CJ, Falkow S, Ramakrishnan L (2014) Host evasion and exploitation schemes of Mycobacterium tuberculosis. Cell 159(7): 1497-1509.
-
Parbhoo T, Mouton JM, Sampson SL (2022) Phenotypic adaptation of Mycobacterium tuberculosis to host- associated stressors that induce persister formation. Frontiers in Cellular and Infection Microbiology 12: 956607.
-
Rittershaus ES, Baek SH, Sassetti CM (2013) The normalcy of dormancy: common themes in microbial quiescence. Cell Host & Microbe 13(6): 643-651.
-
Fang C, Huang H, Xu Z (2016) Immunological evidence for the role of mycobacteria in sarcoidosis: a meta- analysis. PLoS One 11(8): e0154716.
-
Weeratunga P, Moller DR, Ho LP (2024) Immune mechanisms of granuloma formation in sarcoidosis and tuberculosis. The Journal of Clinical Investigation 134(1).
-
Ribeiro FM, Goldenberg T (2015) Mycobacteria and autoimmunity. Lupus 24(4-5): 374-381.
-
Ncube P, Bagheri B, Goosen WJ, Miller MA, Sampson SL (2022) Evidence, challenges, and knowledge gaps regarding latent tuberculosis in animals. Microorganisms 10(9): 1845.
-
WHO (2017) Global Tuberculosis Report 2017. World Health Organization Press, Geneva, Switzerland.
-
Alcantara CA, Glassman I, Nguyen KH, Parthasarathy A, Venketaraman V (2023) Neutrophils in Mycobacterium tuberculosis. Vaccines 11(3): 631.
-
Maio FD, Squeglia F, Goletti D, Delogu G (2019) The mycobacterial HBHA protein: a promising biomarker for tuberculosis. Current Medicinal Chemistry 26(11): 2051-2060.
-
Chandra P, Grigsby SJ, Philips JA (2022) Immune evasion and provocation by Mycobacterium tuberculosis. Nature Reviews Microbiology 20(12): 750-766.
-
Wodelo W, Wampande EM, Andama A, Kateete DP, Ssekatawa K (2024) Polymorphisms in immune genes and their association with tuberculosis susceptibility: An analysis of the African population. Applied Clinical Genetics 17: 33-46.
-
Zhuang L, Yang L, Li L, Ye Z, Gong W (2024) Mycobacterium tuberculosis: immune response, biomarkers, and therapeutic intervention. MedComm 5(1): e419.
-
Christen U (2014) Molecular mimicry. Elsevier eBooks pp: 35-42.
-
Martins YC, Jurberg AD, Daniel-Ribeiro CT (2023) Visiting molecular mimicry once more: pathogenicity, virulence, and autoimmunity. Microorganisms 11(6): 1472.
-
Peterson LK, Fujinami RS (2007) Molecular mimicry. Elsevier eBooks pp: 13-19.
-
Neamțu M, Bild V, Vasincu A, Arcan OD, Bulea D, et al. (2024) Inflammasome molecular insights in autoimmune diseases. Current Issues in Molecular Biology 46(4): 3502-3532.
-
Rojas M, Restrepo-Jiménez P, Monsalve DM, Pacheco Y, Acosta-Ampudia Y, et al. (2018) Molecular mimicry and autoimmunity. Journal of Autoimmunity 95: 100-123.
-
Davies JM (1997) Molecular mimicry: Can epitope mimicry induce autoimmune disease?. Immunology and Cell Biology 75(2): 113-126.
-
Rao AP, Patro D (2023) The intricate dance of infections and autoimmunity: An interesting paradox. The Indian Journal of Pediatrics 91(9): 941-948.
-
Whiteside SK, Snook JP, Williams MA, Weis JJ (2018) Bystander T cells: A balancing act of friends and foes. Trends in Immunology 39(12): 1021-1035.
-
Torres BA, Kominsky S, Perrin GQ, Hobeika AC, Johnson HM (2001) Superantigens: The good, the bad, and the ugly. Experimental Biology and Medicine 226(3): 164- 176.
-
Truant SN, Redolfi DM, Sarratea MB, Malchiodi EL, Fernández MM (2002) Superantigens, a paradox of the immune response. Toxins 14(11): 800.
-
Burns J, Littlefield K, Gill J, Trotter JL (1992) Bacterial toxin superantigens activate human T lymphocytes reactive with myelin autoantigens. Annals of Neurology 32(3): 352-357.
-
Amaya-Uribe L, Rojas M, Azizi G, Anaya JM, Gershwin ME (2019) Primary immunodeficiency and autoimmunity: A comprehensive review. Journal of Autoimmunity 99: 52-72.
-
Vaishnani J (2009) Superantigen. Indian Journal of Dermatology Venereology and Leprology 75(5): 540- 544.
-
Li H, Llera A, Malchiodi EL, Mariuzza RA (1999) The structural basis of T cell activation by superantigens. Annual Review of Immunology 17(1): 435-466.
-
Acha-Orbea H (1993) Bacterial and viral superantigens: Roles in autoimmunity?. Annals of the Rheumatic Diseases 52(Supplement 1): S6-16.
-
Pacheco Y, Acosta-Ampudia Y, Monsalve DM, Chang C, Gershwin ME, et al. (2019) Bystander activation and autoimmunity. Journal of Autoimmunity 103: 102301.
-
Guarneri F (2022) The concept of infection-triggered autoimmunity. Role of Microorganisms in Pathogenesis and Management of Autoimmune Diseases, Singapore, pp: 14-15.
-
Chan LS, Vanderlugt CJ, Hashimoto T, Nishikawa T, Zone JJ, et al. (1998) Epitope spreading: Lessons from autoimmune skin diseases. Journal of Investigative Dermatology 110(2): 103-109.
-
Didona D, Zenzo GD (2018) Humoral epitope spreading in autoimmune bullous diseases. Frontiers in Immunology 9: 779.
-
James JA, Harley JB (1998) B‐cell epitope spreading in autoimmunity. Immunological Reviews 164(1): 185- 200.
-
Floreani A, Leung PSC, Gershwin ME (2015) Environmental basis of autoimmunity. Clinical Reviews in Allergy & Immunology 50(3): 287-300.
-
Powell AM, Black MM (2001) Epitope spreading: protection from pathogens, but propagation of autoimmunity?. Clinical and Experimental Dermatology 26(5): 427-433.
-
Cornaby C, Gibbons L, Mayhew V, Sloan CS, Welling A, et al. (2014) B cell epitope spreading: Mechanisms and contribution to autoimmune diseases. Immunology Letters 163(1): 56-68.
-
Gowen BB, Wong MH, Jung KH, Sanders AB, Mitchell WM, et al. (2007) TLR3 is essential for the induction of protective immunity against Punta Toro virus infection by the double stranded RNA (dsRNA), poly(I), but not poly(I): differential recognition of synthetic dsRNA molecules. Journal of Immunology 178(8): 5200-5208.
-
Medzhitov R, Janeway CA (1997) Innate immunity: impact on the adaptive immune response. Current Opinion in Immunology 9(1): 4-9.
-
Janeway CA, Medzhitov R (2002) Innate immune recognition. Annual Review of Immunology 20(1): 197- 216.
-
Kumar H, Kawai T, Akira S (2011) Pathogen recognition by the innate immune system. Int Rev Immunol 30(1): 16-34.
-
Garg AD, Nowis D, Golab J, Vandenabeele P, Krysko DV, et al. (2010) Immunogenic cell death, DAMPs and anticancer therapeutics: An emerging amalgamation. Biochimica et Biophysica Acta (BBA)- Reviews on Cancer 1805(1): 53-71.
-
Carpenter S, Oneill LAJ (2007) How important are Toll- like receptors for antimicrobial responses?. Cellular Microbiology 9(8): 1891-1901.
-
Cheng TF, Brzostek S, Ando O, Scoy SV, Kumar K, et al. (2006) Differential activation of IFN regulatory factor (IRF)-3 and IRF-5 transcription factors during viral infection. The Journal of Immunology 176(12): 7462- 7470.
-
Mogensen TH (2009) Pathogen recognition and inflammatory signaling in innate immune defenses. Clinical Microbiology Reviews 22(2): 240-273.
-
Wigley P (2013) Immunity to bacterial infection in the chicken. Developmental & Comparative Immunology 41(3): 413-417.
-
Takeuchi O, Akira S (2010) Pattern recognition receptors and inflammation. Cell 140(6): 805-820.
-
Takeda k, Akira S (2015) Toll‐like receptors. Current Protocols in Immunology 1: 109.
-
Chow A, Zhou W, Liu L, Fong Y, Champer Y, et al. (2014) Macrophage immunomodulation by breast cancer- derived exosomes requires Toll-like receptor 2-mediated activation of NF-κB. Sci Rep 4: 5750.
-
Cugno M, Meroni PL, Gualtierotti R, Griffini S, Grovetti E, et al. (2020) Complement activation in patients with COVID-19: A novel therapeutic target. Journal of Allergy and Clinical Immunology 146(1): 215-217.
-
Sakaguchi S, Yamaguchi T, Nomura T, Ono M (2008) Regulatory T cells and immune tolerance. Cell 133(5): 775-787.
-
Kukreja A, Cost G, Marker J, Zhang C, Sun Z, et al. (2002) Multiple immuno-regulatory defects in type-1 diabetes. Journal of Clinical Investigation 109(1): 131-140.
-
Cao D, Vollenhoven RV, Klareskog L, Trollmo C, Malmström V (2004) CD25brightCD4+ regulatory T cells are enriched in inflamed joints of patients with chronic rheumatic disease. Arthritis Research & Therapy 6(4).
-
Moradi B, Schnatzer P, Hagmann N, Rosshirt T, Gotterbarm J, et al. (2014) CD4+CD25+/highCD127low/- regulatory T cells are enriched in rheumatoid arthritis and osteoarthritis joints-analysis of frequency and phenotype in synovial membrane, synovial fluid and peripheral blood. Arthritis Research & Therapy 16(2).
-
Samson M, Audia S, Janikashvili N, Ciudad M, Trad M, et al. (2012) Brief Report: Inhibition of interleukin‐6 function corrects Th17/Treg cell imbalance in patients with rheumatoid arthritis. Arthritis & Rheumatism 64(8): 2499-2503.
-
Zhang X, Zhuang L, Xu C, Li T, Zhang G, et al. (2018) Decreased regulatory T-cell frequency and interleukin-35 levels in patients with rheumatoid arthritis. Experimental and Therapeutic Medicine.
-
Pedros C, Duguet F, Saoudi A, Chabod M (2016) Disrupted regulatory T cell homeostasis in inflammatory bowel diseases. World Journal of Gastroenterology 22(3): 974.
-
Duarte JD, Woods K, Andrews MC, Behren A (2018) The good, the (not so) bad and the ugly of immune homeostasis in melanoma. Immunology and Cell Biology 96(5): 497-506.
-
Brouwer S, Rivera-Hernandez T, Curren BF, Harbison- Price N, Oliveira DDD, et al. (2023) Pathogenesis, epidemiology and control of Group A Streptococcus infection. Nature Reviews Microbiology 21(7): 431-447.
-
Cunningham MW (2019) Molecular mimicry, autoimmunity, and infection: the cross-reactive antigens of group A streptococci and their sequelae. Microbiol Spectr 7(4): 10.
-
Youssefi M, Tafaghodi M, Farsiani H, Ghazvini K, Keikha M (2021) Helicobacter pylori infection and autoimmune diseases; Is there an association with systemic lupus erythematosus, rheumatoid arthritis, autoimmune atrophic gastritis and autoimmune pancreatitis?. A systematic review and meta-analysis study. Journal of Microbiology, Immunology and Infection 54(3): 359-369.
-
Alpízar-Rodríguez D, Finckh A, Gilbert B (2020) The Role of Nutritional Factors and Intestinal Microbiota in Rheumatoid Arthritis Development. Nutrients 13(1): 96.
-
Finsterer J (2022) Triggers of Guillain–Barré syndrome: campylobacter jejuni predominates. International Journal of Molecular Sciences 23(22): L14222.
-
Alpizar-Rodriguez D, Lesker TR, Gronow A, Gilbert B, Raemy, et al. (2019) Prevotella copri in individuals at risk for rheumatoid arthritis. Annals of Rheumatic Diseases 78(5): 590–593.
-
Zangiabadian M, Mirsaeidi M, Pooyafar MH, Goudarzi M, Nasiri MJ (2021) Associations of Yersinia Enterocolitica infection with autoimmune thyroid diseases: a systematic review and meta-analysis. Endocrine, Metabolic & Immune Disorders-Drug Targets 21(4): 682-687.
-
Zheng P, Li Z, Zhou Z (2018) Gut microbiome in type 1 diabetes: A comprehensive review. Diabetes/ Metabolism Research and Reviews 34(7): e3043.
-
Jacobs BM, Giovannoni G, Cuzick J, Dobson R (2020) Systematic review and meta-analysis of the association between Epstein–Barr virus, multiple sclerosis and other risk factors. Mult Scler 26(11): 1281-1297.
-
Molon RS, Rossa C, Thurlings RM, Cirelli JA, Koenders MI (2019) Linkage of periodontitis and rheumatoid arthritis: current evidence and potential biological interactions. Int J Mol Sci 20(18): 4541.
-
Battaglia M, Garrett-Sinha LA (2021) Bacterial infections in lupus: Roles in promoting immune activation and in pathogenesis of the disease. J Transl Autoimmun 4: 100078.
-
Yamaguchi T, Costabel U, McDowell A, Guzman J, Uchida K, et al. (2021) Immunohistochemical detection of potential microbial antigens in granulomas in the diagnosis of sarcoidosis. J Clin Med 10(5): 983.
-
Alam J, Kim YC, Choi Y (2014) Potential role of bacterial infection in autoimmune diseases: a new aspect of molecular mimicry. Immune Network 14(1): 7-13.
-
Park CS, Kim SH, Lee CK (2020) Immunotherapy of autoimmune diseases with nonantibiotic properties of tetracyclines. Immune Netw 20(6): e47.
-
Stone M, Fortin PR, Pacheco-Tena C, Inman RD (2003) Should tetracycline treatment be used more extensively for rheumatoid arthritis?. Meta-analysis demonstrates clinical benefit with reduction in disease activity. The Journal of Rheumatology 30(10): 2112-2122.
-
Dell JR, Paulsen G, Haire CE, Blakely K, Palmer W, et al. (1999) Treatment of early seropositive rheumatoid arthritis with minocycline: Four-year follow-up of a double-blind, placebo-controlled trial. Arthritis & Rheumatism 42(8): 1691-1695.
-
Dell JRO, Elliott JR, Mallek JA, Mikuls TR, Weaver CA, et al. (2006) Treatment of early seropositive rheumatoid arthritis: doxycycline plus methotrexate versus methotrexate alone. Arthritis & Rheumatism: Official Journal of the American College 54(2): 621-627.
-
Kwiatkowska B, Maślińska M (2012) Macrolide therapy in chronic inflammatory diseases. Mediators of Inflammation 2012: 636157.
-
Ogrendik M (2007) Effects of clarithromycin in patients with active rheumatoid arthritis. Curr Med Res Opin 23(3): 515-522.
-
Ogrendik M, Karagoz N (2011) Treatment of rheumatoid arthritis with roxithromycin: A randomized trial. Postgrad Med 123(5): 220-227.
-
Wimer SM, Schoonover L, Garrison MW (1998) Levofloxacin: A therapeutic review. Clin Ther 20(6)6: 1049-1070.
-
Ogrendik M (2007) Levofloxacin treatment in patients with rheumatoid arthritis receiving methotrexate. South Med J 100(2): 135-139.
-
Adegbola SO, Sahnan K, Warusavitarne J, Hart A, Tozer P (2018) Anti-TNF therapy in Crohn’s disease. Int J Mol Sci 19(8): 2244.
-
Feldmann M, Brennan F, Paleolog E, Cope A, Taylor P, et al. (2004) Anti-TNFα therapy of rheumatoid arthritis: What can we learn about chronic disease?. In: Chadwick DJ, et al. (Eds.), Cancer and Inflammation. Novartis Foundation, pp: 53-73.
-
Tanaka Y, Takeuchi T, Mimori T, Saito K, Nawata M, et al. (2010) Discontinuation of infliximab after attaining low disease activity in patients with rheumatoid arthritis: RRR (remission induction by Remicade in RA) study. Ann Rheum Dis 69(7): 1286-1291.
-
Zian Z, Berry SPD, Bahmaie N, Ghotbi D, Kashif A, et al. (2021) The clinical efficacy of Rituximab administration in autoimmunity disorders, primary immunodeficiency diseases and malignancies. Int Immunopharmacol 95: 107565.
-
Ozbek B, Hui-Yuen JS (2021) Emerging B-cell therapies in systemic lupus erythematosus. Ther Clin Risk Manag 17: 39-54.
-
Korhonen R, Moilanen E (2010) Anti-CD20 antibody rituximab in the treatment of rheumatoid arthritis. Basic Clin Pharmacol Toxicol 106: 13-21.
-
Schioppo T Ingegnoli F (2017) Current perspective on rituximab in rheumatic diseases. Drug Des Devel Ther 11: 2891-2904.
-
Ren W, Zhao L, Sun Y, Wang X, Shi X (2023) HMGB1 and Toll-like receptors: Potential therapeutic targets in autoimmune diseases. Mol Med 29: 117.
-
Kim A, Fang X, Abed OA, Moon JJ (2023) Vaccines for immune tolerance against autoimmune disease. Adv Drug Deliv Rev 203: 115140.
-
Leslie M (2023) Inverse vaccine could help tame autoimmune diseases. Science.
-
Tenchov R (2024) Are inverse vaccines the cure for autoimmune diseases. American Chemical Society.
-
Dolgin E (2010) The inverse of immunity. Nature Medicine 16(7): 740–743.
-
Batzloff MR, Pandey M, Olive C, Good MF (2006) Advances in potential M-protein peptide-based vaccines for preventing rheumatic fever and rheumatic heart disease. Immunol Res 35: 233-248.
-
Fan J, Toth I, Stephenson RJ (2024) Recent Scientific Advancements towards a Vaccine against Group A Streptococcus. Vaccines 12(3): 272.
-
Pandey M, Batzloff MR, Good MF (2012) Vaccination against rheumatic heart disease: A review of current research strategies and challenges. Curr Infect Dis Rep 14: 381-390.
-
Guilherme L, Ferreira FM, Köhler KF, Postol E, Kalil J (2013) A vaccine against Streptococcus pyogenes: The potential to prevent rheumatic fever and rheumatic heart disease. AJ Cardiovasc Drugs 13(1): 1-4.
-
Troese MJ, Burlet E, Cunningham MW, Alvarez K, Bentley R, et al. (2023) Group A Streptococcus Vaccine Targeting the Erythrogenic Toxins SpeA and SpeB Is Safe and Immunogenic in Rabbits and Does Not Induce Antibodies Associated with Autoimmunity. Vaccines 11(9): 1504.
-
Reynolds S, Rafeek RAM, Hamlin A, Lepletier A, Pandey M, et al. (2023) Streptococcus pyogenes vaccine candidates do not induce autoimmune responses in a rheumatic heart disease model. npj Vaccines 8(1): 9.
-
Asturias EJ, Excler JL, Ackland J, Cavaleri M, Fulurija A, et al. (2023) Safety of Streptococcus pyogenes Vaccines: Anticipating and Overcoming Challenges for Clinical Trials and Post-Marketing Monitoring. Clinical Infectious Diseases 77(6): 917-924.
-
Li S, Zhao W, Xia L, Kong L, Yang L (2023) How Long Will It Take to Launch an Effective Helicobacter pylori Vaccine for Humans?. Infection and Drug Resistance 16: 3787-3805.
-
Zhang H, Liu Z, Li Y, Tao Z, Shen L, et al. (2024) Adjuvants for Helicobacter pylori vaccines: Outer membrane vesicles provide an alternative strategy. Virulence.
-
Cui M, Ji X, Guan F, Su G, Du L (2024) Design of a Helicobacter pylori multi-epitope vaccine based on immunoinformatics. Frontiers in Immunology 15.
-
Aebischer T, Meyer TF, Andersen LP (2010) Inflammation, immunity, and vaccines for Helicobacter. Helicobacter 15(S1): 21-28.
-
Abadi ATB (2016) Vaccine against Helicobacter pylori: Inevitable approach. World Journal of Gastroenterology 22(11): 3150.
-
Vadalà M, Poddighe D, Laurino C, Palmieri B (2017) Vaccination and autoimmune diseases: Is prevention of adverse health effects on the horizon?. EPMA J 8(3): 295- 311.
-
Brouwer S, Rivera-Hernandez T, Curren BF, Harbison-Price N, Oliveira DMPD (2023) Pathogenesis, epidemiology and control of Group A Streptococcus infection. Nat Rev Microbiol 21(7): 431-447.
-
De Luca F, Shoenfeld Y (2019) The microbiome in autoimmune diseases. Clin Exp Immunol 195(1): 74-85.
-
Christen U (2019) Pathogen infection and autoimmune disease. Clin Exp Immunol195(1): 10-14.
-
Cusick MF, Libbey JE, Fujinami RS (2012) Molecular mimicry as a mechanism of autoimmune disease. Clin Rev Allergy Immunol 42(1): 102–111.
-
Hou Y, Lin X, Shen W, Wang Y (2022) Emerging role of molecular mimicry in autoimmune diseases associated with infection. Infect Genet Evol 93: 104924.
-
Silva MT (2012) Immune evasion strategies of Mycobacterium tuberculosis. Trends Microbiol 20 (12): 602-610.
-
Xiang Y, Zhang M, Jiang D, Su Q, Shi J (2023) The role of inflammation in autoimmune disease: A therapeutic target. Front Immunol 14: 1267091.
-
Nicastro L, Tukel C (2019) Bacterial amyloids: The link between bacterial infections and autoimmunity. Trends Microbiol 27(11): 954-963.
-
Monteleone G, Moscardelli A, Colella A, Marafini I, Salvatori S (2023) Immune-mediated inflammatory diseases: Common and different pathogenic and clinical features. Autoimmunity Reviews 22(10): 103410.
-
Wildner G (2023) Antigenic mimicry-The key to autoimmunity in immune privileged organs. J Autoimmun137: 102942.
-
Brossart P (2020) The role of antigen spreading in the efficacy of immunotherapies. Clin Cancer Res 26(17): 4442-4447.
-
Venkatesha SH, Durai M, Moudgil KD (2024) Epitope spreading in autoimmune diseases. In: Mahroum N, et al. (Eds.), Infection and Autoimmunity. 3rd(Edn.), Academic Press pp: 61-89.
-
Pisetsky DS (2020) Evolving story of autoantibodies in systemic lupus erythematosus. J Autoimmun 110: 102356.
-
Glassner KL, Abraham BP, Quigley EM (2020) The microbiome and inflammatory bowel disease. J Allergy Clin Immunol 145(1): 16-27.
-
Serrano-Gómez G, Mayorga L, Oyarzun I, Roca J, Borruel N, et al. (2021) Dysbiosis and relapse-related microbiome in inflammatory bowel disease: A shotgun metagenomic approach. Comput Struct Biotechnol J 19: 6481-6489.
-
Murdaca G, Greco M, Borro M, Gangemi S (2021) Hygiene hypothesis and autoimmune diseases: A narrative review of clinical evidences and mechanisms. Autoimmunity Reviews 20(7): 102845.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania