Benomyl- or Carbendazim-Induced Androgen Receptor Disrupting Might Lead to Spinal and Bulbar Muscular Atrophy in Multi-Generations of Rats
We have reported that benomyl and its metabolite carbendazim induced reproductive and developmental toxicity and endocrine-disrupting activity in rats. The exactly underlying mechanism of reproductive and developmental toxicity and endocrine-disrupting activity still remain unclear. This manuscript aimed to review and infer the conclusion based on some research, our unpublished data and previous reports. Androgen receptor plays an important role in benomyl- and carbendazim-induced reproductive and developmental toxicity and endocrine-disrupting activity. The evidences were (1) androgen- and androgen receptor-dependent mechanisms are possibly involved in carbendazim-induced toxicity; (2) Our unpublished data showed that the antiandrogen flutamide can completely recover the reproductive and developmental toxicity including embryolethality induced by in utero exposure to benomyl and carbendazim in rats. Based on the previous reports of relationship between androgen receptor expression and spinal and bulbar muscular atrophy (SBMA) or Kennedy’s disease, our unpublished data supported this hypothesis by showing carbendazim-induced spinal and bulbar muscular atrophy in Wistar offspring rats. The molecular underlying mechanism of reproductive and developmental toxicity, endocrine-disrupting activity and spinal and bulbar muscular atrophy induced by benomyl and carbendazim through androgen receptor need to be further investigated.
Introduction
Previous reports showed that co-localization of androgen receptor and nitric oxide synthase in the ventral premammillary nucleus of the newborn rat: an immunohistochemical study [1]. Vito, et al. reported androgen and estrogen receptors expressed in embryonic and neonatal rat brain [2]. The most important finding was that ontogeny of androgen receptor expression was found in spinal nucleus of the bulbocavernosus motoneurons and their target muscles in male mice [3]. Later, Scaramuzzino, et al. reported that protein arginine methyltransferase 6 enhances polyglutamine-expanded androgen receptor function and toxicity in spinal and bulbar muscular atrophy [4]. Based on the issue some researchers was tried to identify neuron selective androgen receptor inhibitors. To identify neuron- selective androgen receptor (AR) signaling inhibitors, which could be useful in the treatment of spinal and bulbar muscular atrophy (SBMA), or Kennedy's disease, a neuromuscular disorder in which deterioration of motor neurons leads to progressive muscle weakness [5]. Furthermore, researchers showed immunohistochemical analysis of huntingtin-associated protein 1 in adult rat spinal cord and its regional relationship with androgen receptor [6].
As reported carbendazim (methyl-2-benzimidazole carbamate) is used to be a systemic fungicide [7]. Both carbendazim and its parent benomyl [methyl 1- (butylcarbamoyl)-2-benzimidazole carbamate] are exhibiting low acute toxicity [8]. In contrast, carbendazim and benzimidazole chemicals induced severe reproductive and developmental toxicity in rodents [9, 10, 11, 12]. Reports showed that carbendazim and benomyl exhibited testicular toxicity such as sloughing of immature spermatids [13, 14], inhibition of microtubule assembly [15], seminiferous tubular atrophy [16], and testicular atrophy and infertility [17] in male rats. Both carbendazim and benomyl induced developmental toxicity in rodents. Prenatal treatment of carbendazim to rats during pregnancy exhibited embryonic death, growth retardation, and developmental abnormalities including exencephaly, microphthalmia and hydronephrosis in offspring [9, 18]. Treatment of benomyl to pregnant rats induced craniocerebral and systemic malformations such as cleft palate, hydrocephalus, and exencephaly in offspring of male and female rats [19]. In contrary to the more reports available on the reproductive and developmental toxicity of carbendazim and benomyl, studies for endocrine-disrupting activity or mode of Lu SY and Tsai WR. Benomyl- or Carbendazim-Induced Androgen Receptor Disrupting Might Lead to Spinal and Bulbar Muscular Atrophy in Multi- Generations of Rats. Pharm Res 2020, 4(2): 000198.
action of these two fungicides remains unclear. The earlier report on endocrine activity for carbendazim might be Rehnberg, et al. [20]. They showed that administration of male rats with carbendazim raised testosterone concentration and the levels of androgen binding protein in the interstitial and seminiferous tubule fluid, meaning an association between endocrine disruption activity and carbendazim toxicity [20]. Recently Rama, et al. reviewed and reported that carbendazim induced reproductive toxicity and possible hormonal effects in rats [21].
They reviewed the previous reports and generalized that carbendazim have androgenic effects acting directly in the androgen receptors and/or increasing the expression of androgen receptors. Some chemicals were reported to increase or decrease AR expression. Bisphenol A was reported to increase AR expression [22] while di-n-butyl phthalate (DBP) [23, 24] and sodium valproate [25] decrease it. We found out that it is common for estrogen receptor agonist and androgen receptor antagonist but not for androgen agonist in pesticides. Based on the chemical structure it seems to determine the AR agonist or antagonist. The degree of increase or decrease of AR expression might be depended on the chemical structure, which shared with nature ligand dihydrotestosterone. This manuscript would like to combine our unpublished data and previous studies to infer that androgen receptor plays an important role in benomyl- and carbendazim-induced reproductive and developmental toxicity, endocrine-disrupting activity and spinal and bulbar muscular atrophy (SBMA), or Kennedy's disease in rats.
Materials and Methods
Study I
Animals and related preparation
The animal use protocol was reviewed and approved by the Institutional Animal Care and Use Committee of the Taiwan Agricultural Chemicals and Toxic Substances Research Institute. Both male and female SD rats, 3–4- week old, were obtained from the National Laboratory Animal Breeding and Research Center, Taipei, Taiwan. All rats were kept in specific-pathogen-free animal facility in Taiwan Agricultural Chemicals and Toxic Substances Research Institute, Taichung, Taiwan. The animal rooms were kept on a 12-h light and dark cycle, 23 ± 2°C, and 50 ± 10% relative humidity. When they were transported to animal room, the rats were quarantined for at least 1 Copyright© Lu SY and Tsai WR.
week and opened on the bases of enough body weight gain and without clinical signs of disease of injury. Both carbendazim and benomyl with 99% pure were a gift from Sinon Co., Taichung, Taiwan. These two pesticides were suspended in polyethylene glycol 200 and treated to animals orally by gavage in a volume of 2 ml/kg body weight, once daily. In reproductive toxicity studies, male rats were administered with these two pesticides and/or flutamide for 28 days. In developmental toxicity studies, both male and female rats were administered with 200 mg/kg carbendazim or 100 mg/kg benomyl for 28 days. Then the female rats were mated with male within each treatment group for 14 days. No treatment was carried out during the mating period. Pregnant dams with plug detected were kept to deliver the offspring at term and conception rates were calculated. All rat offspring were weaned at 21 days postnatal and then fed up to 6- week old [26]. Organ and Tissue Weight, Morphology, and Histopathological Examination Both testis and epididymis were weighed by right and left sides. The incidence of abnormal morphology was recorded. The half of testis or epididymis each was fixed in 10% neutral phosphate-buffered formalin solution for subsequent histopathological examinations. Tissues of testis and epididymis were processed by standard histopathological processes and stained using haematoxylin and eosin for light-microscopic examinations. Both testis and epididymis sections were stained with the Giemsa staining periodic acid-Schiff methods and then counterstained using haematoxylin as reported in Simoes and Schoning [27]. Both testis and epididymis histology were evaluated and histopathological findings were scored according to Oakberg [28] and Hess [29].
In Vitro Androgen Receptor Binding Assay
The ligand binding assay was processed to determine the concentration of androgen receptor in rat tissue according to Nonneman, et al. [30]. The ligand [1, 2, 3, 5, 6, 7-3H(N)]-5α-androstan-17β-ol-3-one (dihydrotestosterone, 5α-DHT) (110–150 Ci/mmol) was obtained from NEN Life Science Products, Inc., Boston. Nonlabelled 5α-DHT was obtained from Sigma Chemical Company, St. Louis, MO, and recrystallized from ethanol prior to use. Both rat testis and epididymis were homogenized in ice-cold low-salt TEDG buffer, pH 7.4, consisting of 10 mM Tris, 1.5 mM ethylenediaminetetraacetic acid (EDTA), 10% glycerol, and 1 mM each of dithiothreitol, phenylmethylsulfonyl Lu SY and Tsai WR. Benomyl- or Carbendazim-Induced Androgen Receptor Disrupting Might Lead to Spinal and Bulbar Muscular Atrophy in Multi- Generations of Rats. Pharm Res 2020, 4(2): 000198.
fluoride, and sodium molybdate as described by Hardy, et al. [31]. Tissue homogenates were centrifuged at 30,000 × g for 1 h and the supernatant were processed to use as the low-salt extract. Before analysis, the endogenous steroids were removed from the low-salt extract by incubation with dextran-coated charcoal. The binding of [3H]-5α- DHT to androgen receptor of testis and epididymis extract each was determined by competitive inhibition binding using nonlabelled 5α-DHT. Charcoal-treated testicular and epididymal extract each was incubated with 1 nM [3H]-5α-DHT at 4°C for 24 h. The nonspecific binding was carried out with incubating the extract with 100-fold excess nonlabelled 5α-DHT. Unbinding [3H]-5α-DHT was isolated from the binding steroid by adding the extract to packed hydroxyapatite in the low-salt TEDG buffer. Mixture were incubated for 30 min with several mixings and then centrifuged at 600 × g for 3 min at 4°C. After that the supernatant was aspirated. An aliquot of the packed HAP was washed 4 times with ice-cold 50 mM Tris buffer, pH 7.3. In determination of total binding, the binding [3H]-5α-DHT was extracted from HAP with ethanol and counted for radioactivity using a Beckman model LS6000 TA liquid scintillator. For specific binding of testis and epididymis extracts they were determined by subtracting nonspecific binding from total binding and corrected for protein concentration. The protein concentration was determined according to Lowry, et al. [32]. Analysis of effect of carbendazim on androgen receptor binding, specific binding of [3H]-5α-DHT to testis extract was carried out with incubation of the charcoal-treated testis extract with [3H]-5α-DHT in the presence of carbendazim at 4°C for 24 h. Incubation mixtures were carried out to the same procedures as before in the androgen receptor binding assay. Statistical analysis All these data were expressed as mean ± SE. All data were processed to analysis of variance followed by Student’s t-test. The level of significance was set at p < 0.05. Results The male rats were co-treatment with 675 mg/kg carbendazim and 50 or 100 mg/kg flutamide, an androgen receptor antagonist, once daily for 28 days decreased testis weight induced by treatment with carbendazim alone. Co-treatment of carbendazim and flutamide blocked losses of spermatozoa and cell morphology and decrease of sperm concentration induced by carbendazim. An important evidence for endocrine disrupting activity induced by carbendazim Copyright© Lu SY and Tsai WR.
and benomyl was that premating treatment of male and female rats with 200 mg/kg carbendazim for 28 days resulted in androgenic effects including incomplete development of uterine horn, enlargement of urethra, absence of vagina, and induction of seminal vesicles in female offspring, without significant effects in male offspring. Also, premating treatment with 100 mg/kg benomyl, the parent compound of carbendazim, produced incomplete development of uterine horn and absence of vagina in female offspring and induced testis and epididymis atrophy in male offspring. When male rats were treated with 25, 50, 100, 200, 400, and 800 mg/kg carbendazim for 56 days androgen receptor concentrations were increased in testis and epididymis with dose dependent. Furthermore, additions of 5, 50, and 500 M carbendazim to testis extract from untreated rats substituted binding of [3H]5 -dihydrotestosterone to androgen receptor with concentration dependent. This study illustrated that reproductive toxicity exhibited by carbendazim is relieved by an androgen receptor antagonist flutamide in male rats and developmental toxicity of the fungicide shows androgenic properties in female offspring. The authors concluded that androgen- and androgen receptor-dependent mechanisms are quite possibly involved in carbendazim-induced toxicity.
Study II
Animals and related preparation
The animal use protocol was reviewed and approved by the Institutional Animal Care and Use Committee of the Taiwan Agricultural Chemicals and Toxic Substances Research Institute. Both male and female Wistar rats, 5- week old, were obtained from the BioLASCO (Taipei, Taiwan, ROC). All rats were kept in specific-pathogen-free animal facility in Taiwan Agricultural Chemicals and Toxic Substances Research Institute, Taichung, Taiwan. The animal rooms were kept on a 12-h light and dark cycle, 23 ± 2°C, and 50 ± 10% relative humidity. When they were transported to animal room, the rats were quarantined for at least 1 week and opened on the bases of enough body weight gain and without clinical signs of disease of injury. Both carbendazim (doses: 0, 6.25, 12.5, 25 and 50 mg/kg/day) and benomyl (doses: 0, 6.25, 12.5, 25, and 50 mg/kg/day) with 99% pure were a gift from Sinon Co., Taichung, Taiwan. These two pesticides were suspended in corn oil and treated to animals orally by gavage in a volume of 2.5 ml/kg body weight, once daily. At 7-week old female rats were transferred into breeding cages in the evening and cohabited overnight with males at 1:1 ratio within treatment. The females were examined the Lu SY and Tsai WR. Benomyl- or Carbendazim-Induced Androgen Receptor Disrupting Might Lead to Spinal and Bulbar Muscular Atrophy in Multi- Generations of Rats. Pharm Res 2020, 4(2): 000198.
next morning for vaginal plugs and sperm in vaginal smears. Successful copulation was confirmed by the presence of vaginal plugs and /or sperm in vaginal smears. Gestation (GD) 0 was defined as the day that plugs or sperm was observed in the vaginal of the female following mating. The pregnant female rats were administered with 0, 6.25, 12.5, 25 and 50 mg/kg/day carbendazim or 0, 6.25, 12.5, 25, and 50 mg/kg/day benomyl for once a day from gestation days 0 through 20. Pregnant dams were kept to deliver the offspring at term and conception rates were calculated. All rat offspring were weaned at 21 days postnatal and then fed up to 8-week old. At 8-week old the F1 male and female rats were mated within treatment and different parents. During the pregnant period the F1 female were not treated as parents. F2 and F3 generations were produced as F1 without treatment. The study was continued to produce offspring until F3 generation is weaned at 21 days postnatal and fed up to 8-week old.
Results
In the first mating, male and female rat treatment with 6.25 mg/kg/day carbendazim within this dose induced 3/8 male and 1/5 female F3 offsprings with spinal and bulbar muscular atrophy (SBMA), or Kennedy's disease. In the second mating, female rats treatment with 6.25 mg/kg/day and male rat treatment with 50 mg/kg/day induced 2/11 male and 2/4 female F3 offsprings with spinal and bulbar muscular atrophy.
Future Work and Recommendations
As we know OECD takes much effort to promote adverse outcome pathways (AOP) methodology. Androgen receptor-mediated reproductive and developmental toxicity, endocrine disrupting activity and spinal and bulbar muscular atrophy (SBMA) would be a novel AOP. It is an approach to support the use of a mode (and/or mechanism) of action basis for understanding the adverse effects of chemicals and other stressors. AR mediated reproductive and developmental toxicity and endocrine disrupting activity would be a novel future application. Specific molecular signals of AR mediated effects would be the future work.
Diagram/Schematic Figure
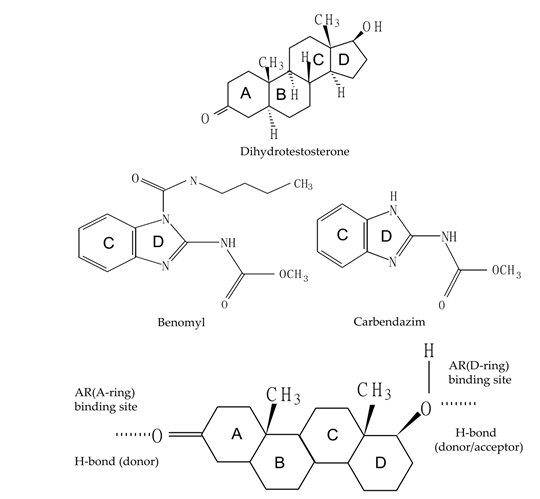
In the principle of AOP, functional group in the chemical structure plays an important role in inducing adverse outcome in mammals. In the respect of chemical structure, benomyl and carbendazim shared the same C Copyright© Lu SY and Tsai WR.
and D ring structure with the natural ligand, dihydrotestosterone (Figure 1). We made a schematic labeling of the benomyl, carbendazim mimicking the main ligand interaction features of the natural ligand, dihydrotestosterone, with the androgen receptor referred to the previous report by Tamura, et al. [33].
Conclusion
Based on the previous study firstly it proved that reproductive toxicity produced by carbendazim is relieved by an androgen receptor antagonist in male rats and developmental toxicity of the pesticide showed androgenic properties in female offspring. We concluded that androgen- and androgen receptor-dependent mechanisms are quite possibly complicated in carbendazim-produced toxicity. Secondly findings show that carbendazim exposure in utero displays a transient and weak androgenic effect and reduces flutamide antiandrogenicity in male rats. Thirdly we concluded that antagonistic effect of flutamide was on the carbendazim androgenic effect on mRNA and protein levels. The results would help us to illustrate the mechanism of carbendazim- and chemical-induced developmental Lu SY and Tsai WR. Benomyl- or Carbendazim-Induced Androgen Receptor Disrupting Might Lead to Spinal and Bulbar Muscular Atrophy in Multi- Generations of Rats. Pharm Res 2020, 4(2): 000198.
toxicity and endocrine disrupting activity. Fourthly benomyl and carbendazim exhibit an androgenic effect, leading to increased weight of ventral prostate and seminal vesicles and uterine fluid retention in young adult rats. Based on the previous reports of relationship between androgen receptor expression and spinal and bulbar muscular atrophy (SBMA) or Kennedy’s disease, our unpublished data supported this hypothesis by showing carbendazim-induced spinal and bulbar muscular atrophy in Wistar offspring rats.
Acknowledgements
The author would like to acknowledge the financial assistance of the Bureau of Animal and Plant Health Inspection and Quarantine, Council of Agriculture, Executive Yuan, Taipei, Taiwan, R.O.C. through the project Copyright© Lu SY and Tsai WR.
104AS-10.8.1-BQ-B1. The authors are grateful to Sinon Co., Taichung, Taiwan, for providing the benomyl and carbendazim standard material used in this study.
References
-
Yokosuka M, Prins GS, Hayashi S (1997) Co- localization of androgen receptor and nitric oxide synthase in the ventral premammillary nucleus of the newborn rat: an immunohistochemical. Brain Res Dev Brain Res 99(2): 226-233.
-
Vito CC, Fox TO (1981) Androgen and estrogen receptors in embryonic and neonatal rat brain. Brain Res 254(1): 97-110.
-
Smith MR, Hamson DK, Poort JE, Jordan CL, Breedlove SM (2012) Ontogeny of androgen receptor expression in spinal nucleus of the bulbocavernosus motoneurons and their target muscles in male mice. Neurosci Lett 513(2): 119-123.
-
Scaramuzzino C, Casci I, Parodi S, Lievens PMJ, Polanco MJ, et al. (2015) Protein arginine methyltransferase 6 enhances polyglutamine- expanded androgen receptorfunction and toxicity in spinal and bulbar muscular atrophy. Neuron 85(1): 88-100.
-
Otto-Duessel M, Tew BY, Vonderfecht S, Moore R, Jones JO (2017) Identification of neuron selective androgen receptor inhibitors. World J Biol Chem. 8(2): 138-150.
-
Islam MN, Takeshita Y, Yanai A, Imagawa A, Jahan MR, et al. (2017) Immunohistochemical analysis of huntingtin-associated protein 1 in adult rat spinal cord and its regional relationship with androgen receptor. Neuroscience 340: 201-217.
-
Vettorazzi GII (1976) Carbamate and organophosphorus pesticides used in agriculture and public health. Residue Reviews 63: 1-76.
-
Seiler JP (1975) Toxicology and genetic effects of benzimidazole compounds. Mutation Research 32(2): 151-168.
-
Cummings AM, Harris ST, Rehnberg GL (1990) Effects of methyl benzimidazolecarbamate during early pregnancy in the rat. Fundam Appl Toxicol 15(3): 528-535. Lu SY and Tsai WR. Benomyl- or Carbendazim-Induced Androgen Receptor Disrupting Might Lead to Spinal and Bulbar Muscular Atrophy in Multi- Generations of Rats. Pharm Res 2020, 4(2): 000198.
-
Gray LE Jr, Ostby J, Linder R, Goldman J, Rehnberg G, et al. (1990) Carbendaziminduced alterations of reproductive development and function in the rat and hamster. Fundame Appl Toxicol 15(2): 281-297.
-
Nakai M, Hess RA, Moore BJ, Guttroff RF, Strader LF, et al (1992) Acute and long-term effects of a single dose of the fungicide carbendazim (methyl 2- benzimidazole carbamate) on the male reproductive system in the rat. Journal of Andrology 13(6): 507- 518.
-
Perreault SD, Jeffay S, Poss P, Laskey JW (1992) Use of the fungicide carbendazim as a model compound to determine the impact of acute chemical exposure during oocyte maturation and fertilization on pregnancy outcome in the hamster. Toxicology and Applied Pharmacology 114(2): 225-231.
-
Parvinen M, Kormano M (1974) Early effects of antispermatogenic benzimidazole derivatives U 32.422 and U 32.104 on the seminiferous epithelium of the rats. Andrologia 6(3): 245-253.
-
Hess RA, Moore BJ, Forrer J, Linder RE, Abuel-Atta AA (1991) The fungicide benomyl(methyl-1- (butylcarbamoyl)-2-benzimidazolecarbamate) causes testicular dysfunction by inducing the sloughing of germ cells and occlusion of efferent ductules. Fundame Appl Toxicol 17(4): 733-745.
-
Lim J, Miller MG (1997) Role of testis exposure levels in the insensitivity of prepubertal rats to carbendazim-induced testicular toxicity. Fundam Appl Toxicol 37(2):158-167.
-
Carter SD, Laskey JM (1982) Effect of benomyl on reproduction in the male rat. Toxicology Letters 1: 87-94.
-
Carter SD, Hess RA, Laskey JW (1987) The fungicide methyl 2-benzimidazole carbamate causes infertility in male Sprague-Dawley rats. Biology of Reproduction 37(3): 709-717.
-
Cummings AM, Ebron-McCoy MT, Rogers JM, Barbee BD, Harris ST (1992) Developmental effects of methyl benzimidazolecarbamate following exposure during early pregnancy. Fundam Appl Toxicol 18(2): 288- 293. Copyright© Lu SY and Tsai WR.
-
Ellis WG, Semple JL, Hoogenboom ER, Kavlock RJ, Zeman FJ (1987) Benomyl-induced craniocerebral anomalies in fetuses of adequately nourished and protein-derived rats. Teratog Carcinog Mutagen 7: 357-375.
-
Rehnberg GL, Cooper RL, Goldman JM, Gray LE, Hein JF, et al. (1989) Serum and testicular testosterone and androgen binding protein profiles following subchronic treatment with carbendazim. Toxicol Appl Pharmacol 101(1): 55-61.
-
Rama EM, Bortolan S, Vieira ML, Gerardin DC, Moreira EG (2014) Reproductive and possible hormonal effects of carbendazim. Regul Toxicol Pharmacol 69(3): 476-486.
-
Huang DY, Zheng CC, Pan Q, Wu SS, Su X, et al. (2018) Oral exposure of lowdose bisphenol a promotes proliferation of dorsolateral prostate and induces epithelialmesenchymal transition in aged rats. Scientific Reports 8(1): 490-499.
-
Jiang JT, Xu HL, Zhu YP, Wood K, Li EH, et al. (2015) Reduced Fgf10/Fgfr2 and androgen receptor (AR) in anorectal malformations male rats induced by di-n- butyl phthalate (DBP): A study on the local and systemic toxicology of DBP. Toxicology 338: 77-85.
-
Jiang JT, Zhong C, Zhu YP, Xu DL, Wood K, et al. (2016) Prenatal exposure to di-n-butyl phthalate (DBP) differentially alters androgen cascade in undeformed versus hypospadiac male rat. Reproductive Toxicology 61: 75-81.
-
Perez-Pouchoulen M, Miquel M, Saft P, Brug B, Toledo R, Hernandez ME, Manzo J (2016) Prenatal exposure to sodium valproate alters androgen receptor expression in the developing cerebellum in a region and age specific manner in male and female rats. Int J Dev Neurosci 53: 46-52.
-
Lu SY, Liao JW, Kuo ML, Wang SC, Hwang JS, et al. (2004) Endocrine-disrupting activity in carbendazim- induced reproductive and developmental toxicity in rats. J Toxicol Environ Health A 67(19): 1501-1515.
-
Simoes JP, Schoning P (1994) Canine mast cell tumors: A comparison of staining techniques. J Vet Diagn Invest 6(4): 458-465.
-
Oakberg EF (1956) A description of spermatogenesis in the mouse and its use in analysis of the cycle of the seminiferous epithelium and germ cell renewal. Am J Anat 99(3): 391-413.
-
Hess RA (1990) Quantitative and qualitative characteristics of stages and transitions in the cycle of the rat seminiferous epithelium: Light microscopic observations of perfusion fixed and plastic-embedded testes. Bio Reprod 43(3): 525-542.
-
Nonneman DJ, Ganjam VK, Welshons WV, Vom Saal FS (1992) Intrauterine position effects on steroid metabolism and steroid receptors of reproductive organs in male mice. Bio Reprod 47: 723-729.
-
Hardy MP, Gelber SJ, Zhou Z, Penning T, Ricigliano JW, et al. (1991) Hormonal control of Leydig cell differentiation. Ann N Y Acad Sci 637: 152-163.
-
Lowry OH, Rosebrough NJ, Farr AL, Randall RL (1951) Protein measurement with the Folin phenol reagent. Journal Biol Chem 193(1): 265-275.
-
Tamura H, Yoshikawa H, Gaido KW, Rose SM, DeLisle RK, et al. (2003) Interaction of organophosphate pesticides and related compounds with androgen receptor. Environ Health Perspect 111(4): 545-552. Lu SY and Tsai WR. Benomyl- or Carbendazim-Induced Androgen Receptor Disrupting Might Lead to Spinal and Bulbar Muscular Atrophy in Multi- Generations of Rats. Pharm Res 2020, 4(2): 000198. Copyright© Lu SY and Tsai WR.
- Acido Labile or Gastro Irritant Apis and Enteric Release in Galenic Practice: An Overview
- A Study on Knowledge, Attitude and Practice of Hand Hygiene among Healthcare Professionals at a Tertiary Care Hospital, India
- Influence of Inoculum Concentration on In Vivo Incubation Period of Emmia lacerata, Pathogenesis and Management of Wilt in Pepper (Capsicum annuum L.)
- Vanilla’s Chemistry
- Marine Anti-Cancer Compounds and Adverse Effects of Global Warming on Oceans: An Overview
- Serological Investigation of Chikungunya Virus Antibody among Malaria-Suspected Febrile Patients in Some Healthcare Facilities in Rivers State