Study of Urease gene and O-Antigen Cluster Gene of Proteus Mirabilis Isolated from Urinary Stones Patients
The aims form this study were to determine the serotypes of Proteus mirabilis isolated from urinary stone patients by using Nuclear magnetic resonance (NMR) spectroscopy and genotype characterizations of UreR gene as well as O-antigen cluster of these isolates via polymerase chain reaction (PCR) technique and sequences alignment process of these genes. The samples were collected from urinary stone patients, bacterial isolates were identified by traditional methods, Vitek 2 system technique and PCR technique, DNA samples were extracted according to the manufacture kit. The lipopolysaccharide and O-antigen of bacterial isolates were isolated, extracted, purified and electrophoresed. The Serotyping of bacterial isolates was determined by using NMR spectroscopy. The genotype study was included two steps, first the detection of UreR gene and O-antigen by PCR technique and seconds the determination the sequencing of these two genes according to Sanger and Coulson method. The determination of Proteus mirabilis serogroups showed that four different types of serogroups for the tested isolates, which included O3, O16, O18, and O20. These serogroups were tested by Nuclear magnetic resonance (NMR) spectroscopy via the determination of the chemical structure of O-antigen repeating units of tested isolates. The detection of the UreR gene and O-antigen by using polymerase chain reaction (PCR) showed that all the tested isolates of P. mirabilis possessed the two genes, where the produced band of UreR gene and O-antigen was 405 bp and 342 bp, which represents the presence of the UreR gene and O-antigen, respectively. The sequencing and the constructed phylogenetic tree results of UreR gene and O-antigen showed that the P. mirabilis isolates were generally closely related to their respective type. NMR spectroscopy was considered a suitable method for determine the serogroup of P. mirabilis based on the determination of chemical structure of O-antigen repeating units. The UreR and O-antigen gene sequence comparison seems to be an appropriate method for inferring genetic relationships within the P. mirabilis isolates on a molecular basis. The UreR gene and O-antigen gene cluster sequences of P. mirabilis isolates allow rapidly a specific PCR assays to be designed.
Mohanad Jawad Kadhim1*, Adnan H Al-Hamadany2 and Eman M Al-
Jarallah3
Engineering, Iraq
Genetic Engineering, Iraq, Tel: 9647801188364; Email: dr.mohanad@biotech.uoqasim.edu.iq polymerase chain reaction (PCR) technique and sequences alignment process of these genes.
seconds the determination the sequencing of these two genes according to Sanger and Coulson method.
determination of the chemical structure of O-antigen repeating units of tested isolates.
O-antigen showed that the P. mirabilis isolates were generally closely related to their respective type.
cluster sequences of P. mirabilis isolates allow rapidly a specific PCR assays to be designed.
Keywords: Struvite stones; NMR spectroscopy; Serotyping; UreR gene; O-antigen cluster
Introduction
Proteus belongs to the Enterobacteriaceae family within the Proteobacteria. They are Gram-negative rods with peritrichous flagella, polymorphic, with a diameter ranging between 0.4 – 0.8 μm, and characterized by rapid motility and by production of the urease enzyme. The genus Proteus currently consists of five named species (P. mirabilis, P. penneri, P. vulgaris, P. myxofaciens and P. hauseri) and three unnamed genomospecies (Proteus genomospecies 4, 5, and 6). Proteus from Homer’s Odyssey was pursued by mortals and gods alike for his ability to foretell the future, but he evaded pursuers by taking the shape of animals, plants, water, or even fire. The term Proteus therefore refers to readily changing appearance: “and has the gift of endless transformation” [1]. P. mirabilis is most often associated with urinary stones and catheter encrustation and is rarely a cause of acute cystitis, hospital-acquired UTIs or recurrent UTIs, but is more often associated with complicated UTIs, intermittent catheterization, condom catheter, and ileal loop [2]. The ability of Proteus spp. to produce urease and to alkalinize the urine by hydrolyzing urea to ammonia makes it effective in producing an environment in which it can survive. This leads to precipitation of organic and inorganic compounds, which leads to struvite stone formation. Struvite stones are composed of a combination of magnesium ammonium phosphate (struvite) and calcium carbonate-apatite [3]. The persistence of a P.mirabilis infection is compounded by the ability of this organism to cause the formation of urinary stones and encrust indwelling catheters, indeed the formation of stones around the organism can make antibiotic treatment ineffective, stone formation requires urease, which catalyzes the hydrolysis of urea into carbon dioxide and ammonia which raises the environmental pH, and mediates precipitation of normally soluble polyvalent ions from the urine, specifically precipitation of magnesium, ammonium, phosphate and calcium ions results in formation of the struvite and carbonate hydroxyapatite crystals that comprise urinary stones [4]. The urease enzyme is comprised of three structural subunits, UreA, UreB, and UreC, assembled as a homotrimer of individual UreABC heterotrimers (means that there are three copies of each subunit) [5]. The lipopolysaccharide (LPS) is considered as an important virulence factor of P. mirabilis. The polysaccharide chain (O-polysaccharide or O antigen) and sometimes the core region of the LPS define the serological specificity of these bacteria [6]. Based on the serospecificity of the lipopolysaccharide (O-antigen), strains of P. mirabilis and P. vulgaris are divided into 49 O-serogroups and later 11 additional O-serogroups have been proposed [7]. The O-specific polysaccharide (O-PS) or O-antigen represents the polymer chain of LPS, it is exposed on the bacterial surface bound, frequently, to the terminal residue of the outer core [8]. The structural diversity of O antigens is remarkable; more than 60 monosaccharides and 30 different non-carbohydrate components have been recognized [9]. In Proteus, the O antigen gene cluster is located between the two housekeeping genes, cpxA (encoding two-component system sensor kinase) and secB (encoding a pre´protein translocase subunit). Similar to the E. coli and Salmonella O antigen gene cluster, the Proteus cluster also contains three major gene classes: sugar biosynthetic pathway genes, sugar transferase genes, and O antigen-processing genes. All of the O antigen-processing genes analyzed were wzx and wzy, indicating that the assembly of Proteus O antigen was likely to be Wzx/Wzy dependent [10]. The molecular characterization of the putative gene clusters responsible for Proteus O antigen biosynthesis in five different serogroups has been reported [10]. Although the O-specific polysaccharides have been identified in most Proteus O antigens, a study of the genetic locus associated with Proteus O antigens has never been carried out [10] as well as the genome sequence of P. mirabilis was published for the first time in 2008 [11].
Material and Methods
Samples Collection
A total of 25 stone of suitable size were collected in sterilized containers and taken to the Department of Microbiology in the College of Veterinary Medicine for bacterial analysis. Stones were washed with sterile saline and crushed under aseptic conditions, then cultured in nutrient broth Nemoy and Stanley. After overnight incubation at 37 °C, they were sub-cultured on MacConkey agar, blood agar and nutrient agar and incubated at 37 oC for 18-24 hr. Colonies growing on MacConkey agar were considered Gram-negative bacteria.
Mid-stream urine of 100 specimens was collected in sterilized screw-cap containers. Urine specimens were centrifugated at 2500 rpm for 15 minutes. The supernant was discarded and sediments were cultured in MacConkey broth and incubated at 37°C for 24 h, followed by streaking on MacConkey agar, blood agar and nutrient agar then incubated at 37°C for 24 h.
Isolation and Identification
The P. mirabilis isolates were identified using macro- microscopic, biochemical test, Vitek 2 system and polymerase chain reaction (PCR) technique by using target gene 16S rRNA, also, the urease activity and other virulence factors of bacterial isolates were previously investigated [12]. LPS and O-antigen were previously isolated and purified [13].
examined in D2O at 45 °C using internal acetone (δH 2.225, δC 31.45) as reference. 1H and13C NMR spectra were recorded with a Bruker DRX-500 spectrometer equipped with an SGI INDY computer workstation. NMR experiments were performed using standard Bruker software, and XWINNMR program (Bruker) was used to acquire and process data. A mixing time of 200 ms was used in TOCSY and ROESY experiments. Also, the results were analyzed by using MestRec software and Bacterial Carbohydrate Structure Database (CSDB) NMR searches with the using of reference strains [14].
Molecular Investigation of UreR and O-antigen genes by PCR
The detection of UreR gene by polymerase chain reaction technique was done as described by Dattelbaum, et al. [15]. the primer (F- 5' GCTGGCGGTTTATCACGAAG – 3'), (R- 5' CGCCATTGTTGCTATTGCATTC – 3') with product size 405 bp. PCR was performed in a 25μl of reaction volume which contained (12.5 µl master mix; 1 µl forward, reverse primer and template DNA; 9.5 PCR water. The PCR conditions were as the following table:
- Detection of P. Mirabilis Serotype by NMR
- The serotype of 10 isolates of P. mirabilis was determine by using
- Nuclear magnetic resonance spectroscopy (NMR) as follow: The O antigen samples
- (2.5 mg) were solubilized in 650 μl deuterated water
- (D2O) and transferred to 5-mm NMR tubes. Then
- PCR cycle repeat
- Temp.
- Time
- Initial denaturation
- 1
- 94 oC
- 5 min
- Denaturation
- 94 oC
- 30 sec.
- Annealing
- 52 oC
- 30 sec.
- Extension
- 72 oC
- 40 sec
- Final extension
- 1
- 72 oC
- 5 min
- Hold
- -
- 4 oC
- -
- 30
Table 1: The PCR conditions of the detection of UreR gene.
- PCR cycle repeat
- Temp.
- Time
- Initial denaturation
- 1
- 94 oC
- 5 min
- Denaturation
- 94 oC
- 40 sec.
- Annealing
- 60 oC
- 1 min.
- Extension
- 72 oC
- 1 min
- Final extension
- 1
- 72 oC
- 5 min
- Hold
- -
- 4 oC
- -
- 30
Table 2: The PCR conditions of the detection of O-antigen cluster.
Also PCR product was visualized according to Sanger F, et al. [16] by using electrophoresis technique.
DNA Sequencing
Both UreR gene and O-antigen were sequenced according to Sanger method with one single strand of primer [18]. DNA sequencing compared with those available in the GenBank database by using basic alignment search tool, BLAST of the NCBI to identify whether they aligned with closely related organisms [19, 20].
Phylogenetic Mapping
Phylogenetic analysis was carried out using Sequence Alignments Explorer CLUSTALW 2.1, and using the neighbors-joining method to reconstruct the phylogenetic tree with the option of complete deletion of gaps [21].
Results
Determination of Serogroup of P. Mirabilis Isolates
After acid degradation of O antigen of P. mirabilis isolates, the NMR spectroscopy was investigated to determine the serogroup of P. mirabilis isolates. The results showed that P. mirabilis isolates number 1,6,10, and 16 were classified under O18 serogroup, while isolates number 3 and 13 were classified under O16 serogroup. The isolates number 18 and 23 were classified under O20 serogroup, while the isolate number 25 and 14 were classified under O3 serogroup as shown in the table 7. P. Mirabilis O16 Serogroup: The 13C NMR spectrum of oligosaccharide was contained signals for three anomeric carbon at δ 100, 95.6, and 95.3; three carbons bearing nitrogen at δ 51.2, 53.6, and 54; five HOCH2 C groups at δ 59- 60.6; also, 12 other carbons bearing oxygen belonging to sugar pyranose rings and ribitol in the region δ 67- 80 and three N-acetyl groups at δ 20.5-20.7 (CH3) and δ 170.6-171.6 (CO) as shown in table 3. The results of 1H NMR spectrum of oligosaccharide showed that oligosaccharide was contained signals for three anomeric protons at δ 5.53, 6.63, and 6.12; and three N-acetyl groups at δ 2- 2.2 as shown in table 3. The 13C and 1H NMR spectrum data were suggested that O- polysaccharide had pentasaccharide-repeating unit, which contained two N-acetylgalactosamine (GalNAc) residues and one residue each of N-acetylglucosamine (GlcNAc) and ribitol. The sequence of residues of the oligosaccharide O16 was shown in the table 5.
| Sugar residue | H1 | H2 | H3 | H4 | H5 | H6 | C1 | C2 | C3 | C4 | C5 | C6 | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β-GalpNAc*-(1→ | 4.69 | 3.92 | 3.77 | 3.93 | 3.67 | 3.8 | 100 | 51.8 | 70 | 68.3 | 75.5 | 60.5 | ||||||||||||||||||||||||||
| →4)-α-GalpNAc*-(1→ | 5.36 | 4.14 | 3.92 | 4.18 | 3.87 | 3.79 | 95.6 | 49.5 | 67 | 75 | 70.5 | 59.2 | ||||||||||||||||||||||||||
| →3)-α-GlcpNAc*-(1→ | 5.11 | 4.05 | 3.89 | 3.69 | 3.8 | 3.71 | 95.3 | 51.4 | 76 | 73.5 | 74.5 | 59.4 | ||||||||||||||||||||||||||
| →4)-Rib-ol | 3.64 | 3.75 | 3.91 | 3.96 | 3.83 | 3.9 | 60.6 | 70.5 | 71 | 80 | 59 |
Table 3: 1H and 13C NMR chemical shifts (δ) of the O-polysaccharide of Proteus mirabilis O16 (δ = ppm).
*Additional chemical shifts for NAc were δH (2-2.2) for CH3 and CO; δC (20.5-20.7) for CH3 and (170.6-171.6) for CO.
P. Mirabilis O18 Serogroup: The results of 13C NMR spectrum of O-polysaccharide of P. mirabilis O18 revealed that O-polysaccharide contained signals for five carbons at δ 90.5 and 103.3; three nitrogen-bearing carbons at δ 50.3 and 51; seven signals for CH2 groups at δ 60.5, 61.3, and 65; and 19 signals for sugar-ring oxygen-bearing carbons in the region δ 65 and 80.5. Signals at δ 20.5, 171.5, and 172.5 were belonged to two N-acetyl groups, and an intense signal at δ 52.5, together with two additional signals for CH2 groups at δ 59 and 65 that were split due to coupling to phosphorus, indicated the presence of choline as shown in table 4. The 1H NMR results of O-polysaccharide of P. mirabilis O18 were shown in the table 4. The results revealed that O-polysaccharide was contained signals for five anomeric protons at δ 5.42 and 6.52; two N-acetyl groups at δ 2.1 and 2.1 and methyl group of choline at δ 4.52. The 13C and 1H NMR spectrum data were suggested that O-polysaccharide had a pentasaccharide-repeating unit, which contained three hexoses residues (two Glcp and one Galp), two N-acetyl hexosoamines residues (GlcpNAc), a choline residue (ChoP), and two phosphate groups. The sequence of residues of the oligosaccharide O18 shown in the table 7.
| Sugar residue | H1 | H2 | H3 | H4 | H5 | H6 | C1 | C2 | C3 | C4 | C5 | C6 | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| → 3,4)-α-GlcpNAc*-l-P- | 6.85 | 5.52 | 5.31 | 5.31 | 4.65 | 4.62 | 90.5 | 50.3 | 75.8 | 72.6 | 72.8 | 60.5 | ||||||||||||||||||||||
| → 3)-β-GlcpNAc*-(1→ | 5.53 | 4.67 | 4.77 | 4.57 | 4.25 | 4.66 | 100.3 | 51 | 80.5 | 68.5 | 74.5 | 60.7 | ||||||||||||||||||||||
| → 3)-β-Galp-(1→ | 5.42 | 4.56 | 4.67 | 5 | 4.65 | 4.67 | 103 | 70 | 79 | 67 | 76 | 61.3 | ||||||||||||||||||||||
| → 4,6)-β-Glcp-(1→ | 5.57 | 4.21 | 4.58 | 4.47 | 4.83 | 5.16 | 103.3 | 73 | 75.5 | 77.7 | 73 | 65 | ||||||||||||||||||||||
| α-Glcp-(1→ | 6.52 | 4.96 | 4.81 | 4.63 | 4.85 | 4.86 | 100 | 71 | 73.5 | 69.5 | 73 | 61 | ||||||||||||||||||||||
| ChoP | 5.67 | 4.75 | 4.52 | 65 | 59 | 52.5 |
Table 4: 1H and 13C NMR chemical shifts (δ) of the O-polysaccharide of Proteus mirabilis O18 (δ = ppm).
*Additional chemical shifts for NAc were δH 2.05 for CH3 and CO; δC 20.5 for CH3 and (171.5, 172.5) for CO. P. Mirabilis O20 Serogroup: The results of 13C NMR spectrum of O-polysaccharide of P. mirabilis O20 were shown in the table 5. The results showed that O- polysaccharide was contained signals for five anomeric carbons at δ 105.5, 104.8, 101.5 and 100; two nitrogen- bearing carbons at δ 50.5 and 51.5; five non-substituted HOCH2-C groups at δ 60.2-61.5; and oxygen-bearing carbons at δ 67.5-80.5. Signals δ at 21.3 and 21.6 (CH3), as well as at δ 174.2 and 174.7 were belonged to two N- acetyl groups.
The results of 1H NMR spectrum of O-polysaccharide of P. mirabilis O20 revealed that O-polysaccharide was contained signals for five anomeric protons at δ 5.5-6.83 and two signals for N-acetyl at δ 2.1 and 2.3 as shown in table 5. The 13C and 1H NMR spectrum data were suggested that O-polysaccharide had a pentasaccharide- repeating unit, which contained three residues of hexoses (two Glc_p_ and one Gal_p_) and two residues of N-acetyl hexosoamines (Glc_p_NAc). The sequence of residues of the oligosaccharide O20 shown in the table 7.
| Sugar residue | H1 | H2 | H3 | H4 | H5 | H6 | C1 | C2 | C3 | C4 | C5 | C6 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α-Glcp-(1→ | 5.33 | 3.58 | 3.12 | 3.43 | 4.02 | 3.82 | 102 | 70.3 | 71.5 | 70 | 72 | 60.2 | |||||||||||||||||||||
| → 2)-β-Galp-(1→ | 4.7 | 3.7 | 3.72 | 3.94 | 3.68 | 3.79 | 105 | 74.9 | 70.7 | 68 | 74.3 | 61 | |||||||||||||||||||||
| → 3,4)-α-GlcpNAc*-(1→ | 5.27 | 4.12 | 4.19 | 4.12 | 3.94 | 3.8 | 100 | 50.5 | 68.5 | 73.5 | 70.5 | 60.2 | |||||||||||||||||||||
| → 4)-β-Glcp-(1→ | 4.45 | 3.31 | 3.62 | 3.73 | 3.79 | 3.75 | 106 | 70.6 | 73.6 | 75 | 75.5 | 61.5 | |||||||||||||||||||||
| → 3)-β-GlcpNAc*-(1→ | 4.94 | 3.84 | 3.76 | 3.44 | 3.39 | 3.67 | 100 | 51.5 | 80.5 | 67.5 | 76 | 61 |
Table 5: 1H and 13C NMR chemical shifts (δ) of the O-polysaccharide of Proteus mirabilis O20 (δ = ppm).
Mirabilis O3 Serogroup: The results of 13C NMR spectrum table 6 revealed that O-polysaccharide was contained signals for four anomeric carbons at δ 99.5- 103.6; one non-substituted (δ 60.2) and one substituted (δ 64.5) C-CH2OH groups; two carboxyl groups at δ 172.5 and 170.2; two carbons-bearing at δ 50.1 and 51; 14 sugar ring carbons-bearing oxygen at δ 66-78.4; N-acetyl groups (CH3 at δ 20.5, CO at δ 174.5 and 174.9); and six carbons of lysine at δ 175.5, 52.5, 30, 20, 21.4 and 40.
The results of 1H NMR spectrum of O-polysaccharide showed that O-polysaccharide was contained signals for four anomeric protons at δ 5.12-6.5; two N-acetyl groups at δ 2.1 and 2.4; and signals for lysine at δ 1.52, 1.69, 1.71, 2.5 and 4.4 as shown in table 6. The 13C and 1H NMR spectrum data were suggested that O-polysaccharide had a tetrasaccharide-repeating unit, which contained one residue each of D-glucuronic acid (D-GlcA) and D- galacturonic acid (D-GalA); two residues of D-GalNAc; and L-lysine. The sequence of residues of the oligosaccharide O3shown in the table 7.
| Sugar residue | H1 | H2 | H3 | H4 | H5 | H6 | C1 | C2 | C3 | C4 | C5 | C6 | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| → 3)-β- -GalpNAc*-(1→ D | 5.16 | 4.98 | 4.57 | 5.83 | 4.37 | 3.78 | 100.5 | 50.1 | 80 | 66 | 75.3 | 60.2 | |||||||||||||||||||||||||
| → 6)-β- -GalpNAc*-(1→ D | 5.12 | 4.96 | 4.5 | 5.1 | 4.45 | 3.89 | 100.9 | 51 | 70 | 74.5 | 72.5 | 64.5 | |||||||||||||||||||||||||
| → 4)-β- -GlcpA-(1→ D | 5.13 | 4.73 | 4.21 | 4.41 | 4.36 | 103.6 | 70.5 | 71.7 | 78.4 | 75.7 | 173 | ||||||||||||||||||||||||||
| α- -GalpA-(1→ D | 6.5 | 4.93 | 5.1 | 5.6 | 5.76 | 99.5 | 65.7 | 66.5 | 65 | 67.9 | 170 | ||||||||||||||||||||||||||
| -Lys L | 4.4 | 1.69 | 1.52 | 1.71 | 2.5 | 175.5 | 52.5 | 30 | 20 | 21.5 | 40 |
Table 6: 1H and 13C NMR chemical shifts (δ) of the O-polysaccharide of Proteus mirabilis O3 (δ = ppm).
| No. of local isolates | Serogroup | O-antigen chemical structure | ||||||
|---|---|---|---|---|---|---|---|---|
| 1, 6, 10, 16 | O18 | α-D-Glcp 1 ChoP ↓ | 4 4 6)-β-D-Glcp-(1→3)-β-D-Galp-(1→3)-β-D-GlcpNAc-(1→3)-α-D-GlcpNAc-l-P- | ||||||
| 3, 13 | O16 | β-D-GalpNAc-(1→4)-α-D-GalpNAc-(1→3)-α-D-GlcpNAc-(1→4)- Rib-ol | ||||||
| 18, 23 | O20 | β- -Galp-(1→2)-α- -Glcp D D 1 ↓ 4 → 3)-α- -GlcpNAc-(1→4)-β- -Glcp-(1→3)-β- -GlcpNAc-(1→ D D D | ||||||
| 14, 25 | O3 | α- -GalpA6 ( -Lys) D L 1 ↓ 4 → 3)-β- -GalpNAc-(1→6)-β- -GalpNAc-(1→4)-β- -GlcpA-(1→ D D D |
Table 7: Chemical structure of O-polysaccharide of 10 P. mirabilis isolated from urinary stones patients.
Detection of UreR gene among P. Mirabilis Isolates using PCR Technique
Results in the figure 1 indicated that P. mirabilis isolates, which contain UreR gene, exhibited positive PCR products on gel which all of the isolates contained that gene. Moreover, the produced band was of 405 bp, which represents the presence of UreR gene.
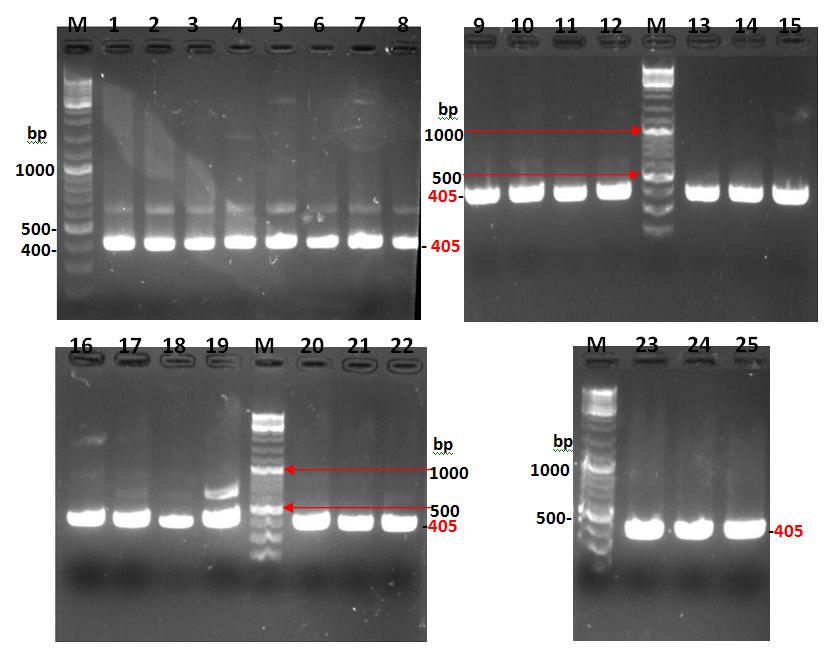
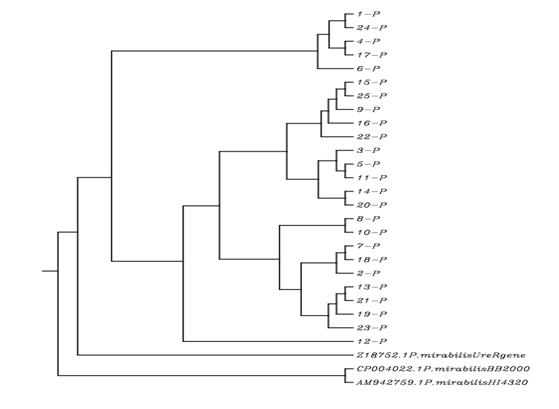
Sequencing of UreR gene of P. Mirabilis Isolates: After the detection of UreR gene of P. mirabilis isolates, the sequencing of isolates had been investigated. The results of sequencing and alignments of these isolates were shown in the table 7, 8 and 9 which obtained by using BLAST and ClustalW 2.1 sites, respectively. The results of NCBI blast of UreR gene sequencing of P. mirabilis isolates showed that the local isolates were had a sequencing significance alignment with three standard strains; P. mirabilis BB2000, P. mirabilis HI4320 and P. mirabilis UreR gene. The identity percentages were ranged from (97.55%-99.44%), (97.28%-98.60%) and (95.66%-97.48%) with P. mirabilis BB2000, P. mirabilis HI4320 and P. mirabilis UreR gene, respectively. The expected value (E-value) was (0.0) with P. mirabilis BB2000, and (0.0) with P. mirabilis HI4320 except for isolates number 16 and 25 where, the E-values were 2e- 179 and 8e-179, respectively. The E-value was ranged from (0.0 to 6e-180) with P. mirabilis UreR gene as shown in the table 7, 8 and 9. The phylogenetic tree results showed that the P. mirabilis isolates were formed a cluster together with Z18752.1 P. mirabilis UreR gene, CP004022.1 P. mirabilis BB2000, and AM942759.1 P. mirabilis HI4320 as shown in figure 2. Thus, the present results were suggested that the UreR gene operon has a common ancestor with these bacterial species.
- Local isolates
- Identity
- Alignment length
- Mismatches
- Gap opens
- Q. start
- Q. end
- S. start
- S. end
- E-value bit score
- 1-P
- 96.74
- 368
- 10
- 2
- 2
- 369
- 11
- 376
- 6e-180
- 612
- 2-P
- 96.65
- 358
- 12
- 0
- 1
- 358
- 5
- 362
- 6e-175
- 595
- 3-P
- 96.91
- 356
- 10
- 1
- 2
- 357
- 7
- 361
- 6e-175
- 595
- 4-P
- 97.02
- 369
- 9
- 2
- 1
- 369
- 14
- 380
- 0
- 619
- 5-P
- 96.74
- 368
- 11
- 1
- 2
- 369
- 18
- 384
- 6e-180
- 612
- 6-P
- 97.01
- 368
- 9
- 2
- 2
- 369
- 16
- 381
- 0
- 617
- 7-P
- 96.69
- 362
- 10
- 2
- 1
- 362
- 12
- 371
- 1e-176
- 601
- 8-P
- 96.92
- 357
- 10
- 1
- 2
- 358
- 14
- 369
- 2e-175
- 597
- 9-P
- 96.21
- 369
- 12
- 2
- 1
- 369
- 12
- 378
- 4e-177
- 603
- 10-P
- 96.21
- 369
- 13
- 1
- 1
- 369
- 11
- 378
- 4e-177
- 603
- 11-P
- 96.48
- 369
- 11
- 2
- 1
- 369
- 14
- 380
- 8e-179
- 608
- 12-P
- 96.48
- 369
- 12
- 1
- 1
- 369
- 13
- 380
- 8e-179
- 608
- 13-P
- 96.93
- 358
- 10
- 1
- 1
- 358
- 11
- 367
- 5e-176
- 599
- 14-P
- 96.48
- 396
- 11
- 2
- 1
- 369
- 15
- 381
- 8e-179
- 608
- 15-P
- 96.48
- 369
- 11
- 2
- 2
- 369
- 15
- 382
- 8e-179
- 608
- 16-P
- 95.66
- 369
- 11
- 5
- 2
- 369
- 13
- 377
- 1e-172
- 588
- 17-P
- 97.48
- 357
- 8
- 1
- 2
- 358
- 12
- 367
- 8e-179
- 608
- 18-P
- 96.2
- 368
- 12
- 2
- 2
- 369
- 10
- 375
- 1e-176
- 601
- 19-P
- 96.93
- 358
- 10
- 1
- 1
- 358
- 12
- 368
- 5e-176
- 599
- 20-P
- 96.48
- 369
- 11
- 2
- 1
- 369
- 14
- 380
- 8e-179
- 608
- 21-P
- 96.93
- 358
- 10
- 1
- 1
- 358
- 11
- 367
- 5e-176
- 599
- 22-P
- 96.47
- 368
- 11
- 2
- 2
- 369
- 15
- 380
- 3e-178
- 606
- 23-P
- 96.48
- 369
- 12
- 1
- 1
- 369
- 13
- 380
- 8e-179
- 608
- 24-P
- 96.75
- 369
- 10
- 2
- 1
- 369
- 13
- 379
- 2e-180
- 614
- 25-P
- 96.35
- 356
- 13
- 0
- 14
- 369
- 2
- 357
- 4e-172
- 586
- %
Table 8: NCBI blast results of UreR gene of P. mirabilis local isolates compared with Z18752.1 P. mirabilis UreR gene
Program: blastn, Iteration: 0, RID: 3X99KZ3B114, Database: n/a, 25 hits found.
Local isolates
Identity
Alignment
length Mismatches Gap opens
bit score 1-P 97.55 368 7 2 1 368 376 11 0 628 2-P 99.16 358 3 0 12 369 362 5 0 645 3-P 99.44 356 1 1 13 368 361 7 0 645 4-P 97.83 369 6 2 1 369 380 14 0 636 5-P 99.18 368 2 1 1 368 384 18 0 662 6-P 97.83 368 6 2 1 368 381 16 0 634 7-P 99.17 362 1 2 8 369 371 12 0 651 8-P 99.44 357 1 1 12 368 369 14 0 647 9-P 98.64 369 3 2 1 369 378 12 0 652 10-P 98.64 369 4 1 1 369 378 11 0 652 11-P 98.92 369 2 2 1 369 380 14 0 658 12-P 98.92 369 3 1 1 369 380 13 0 658 13-P 99.44 358 1 1 12 369 367 11 0 649 14-P 98.92 369 2 2 1 369 381 15 0 658 15-P 98.92 369 2 2 1 368 382 15 0 658 % Q. start Q. end S. start S. end E- value
| 16-P | 98.1 | 369 | 2 | 5 | 1 | 368 | 377 | 13 | 0 | 638 |
|---|---|---|---|---|---|---|---|---|---|---|
| 17-P | 98.32 | 357 | 5 | 1 | 12 | 368 | 367 | 12 | 0 | 625 |
| 18-P | 98.64 | 368 | 3 | 2 | 1 | 368 | 375 | 10 | 0 | 651 |
| 19-P | 99.44 | 358 | 1 | 1 | 12 | 369 | 368 | 12 | 0 | 649 |
| 20-P | 98.92 | 369 | 2 | 2 | 1 | 369 | 380 | 14 | 0 | 658 |
| 21-P | 99.44 | 358 | 1 | 1 | 12 | 369 | 367 | 11 | 0 | 649 |
| 22-P | 98.91 | 368 | 2 | 2 | 1 | 368 | 380 | 15 | 0 | 656 |
| 23-P | 98.92 | 369 | 3 | 1 | 1 | 369 | 380 | 13 | 0 | 658 |
| 24-P | 97.56 | 369 | 7 | 2 | 1 | 369 | 379 | 13 | 0 | 630 |
| 25-P | 98.88 | 356 | 4 | 0 | 1 | 356 | 357 | 2 | 0 | 636 |
Table 9: NCBI blast results of UreR gene of P. mirabilis local isolates compared with CP004022.1 P. mirabilis BB2000
- Standard strain.
- Program: blastn, Iteration: 0, RID: 3X99KZ3B114, Database: N/A, 25 hits found.
- Local isolates
- Identity
- Alignment length
- Mismatches
- Gap opens
- Q. start
- Q. end
- S. start
- S. end
- E-value bit score
- 1-P
- 97.83
- 368
- 6
- 2
- 1
- 368
- 376
- 11
- 0
- 634
- 2-P
- 97.77
- 358
- 8
- 0
- 12
- 369
- 362
- 5
- 0
- 617
- 3-P
- 98.03
- 356
- 6
- 1
- 13
- 368
- 361
- 7
- 0
- 617
- 4-P
- 98.1
- 369
- 5
- 2
- 1
- 369
- 380
- 14
- 0
- 641
- 5-P
- 97.83
- 368
- 7
- 1
- 1
- 368
- 384
- 18
- 0
- 634
- 6-P
- 98.1
- 368
- 5
- 2
- 1
- 368
- 381
- 16
- 0
- 640
- 7-P
- 97.79
- 362
- 6
- 2
- 8
- 369
- 371
- 12
- 0
- 623
- 8-P
- 98.04
- 357
- 6
- 1
- 12
- 368
- 369
- 14
- 0
- 619
- 9-P
- 97.29
- 369
- 8
- 2
- 1
- 369
- 378
- 12
- 0
- 625
- 10-P
- 97.29
- 369
- 9
- 1
- 1
- 369
- 378
- 11
- 0
- 625
- 11-P
- 97.56
- 369
- 7
- 2
- 1
- 369
- 380
- 14
- 0
- 630
- 12-P
- 97.56
- 369
- 8
- 1
- 1
- 369
- 380
- 13
- 0
- 630
- 13-P
- 98.04
- 358
- 6
- 1
- 12
- 369
- 367
- 11
- 0
- 621
- 14-P
- 97.56
- 369
- 7
- 2
- 1
- 369
- 381
- 15
- 0
- 630
- 15-P
- 97.56
- 369
- 7
- 2
- 1
- 368
- 382
- 15
- 0
- 630
- 16-P
- 96.75
- 369
- 7
- 5
- 1
- 368
- 377
- 13
- 2e-179
- 610
- 17-P
- 98.6
- 357
- 4
- 1
- 12
- 368
- 367
- 12
- 0
- 630
- 18-P
- 97.28
- 368
- 8
- 2
- 1
- 368
- 375
- 10
- 0
- 623
- 19-P
- 98.04
- 358
- 6
- 1
- 12
- 369
- 368
- 12
- 0
- 621
- 20-P
- 97.56
- 369
- 7
- 2
- 1
- 369
- 380
- 14
- 0
- 630
- 21-P
- 98.04
- 358
- 6
- 1
- 12
- 369
- 367
- 11
- 0
- 621
- 22-P
- 97.55
- 368
- 7
- 2
- 1
- 368
- 380
- 15
- 0
- 628
- 23-P
- 97.56
- 369
- 8
- 1
- 1
- 369
- 380
- 13
- 0
- 630
- 24-P
- 97.83
- 369
- 6
- 2
- 1
- 369
- 379
- 13
- 0
- 636
- 25-P
- 97.47
- 356
- 9
- 0
- 1
- 356
- 357
- 2
- 8e-179
- 608
- %
Table 10: NCBI blast results of UreR gene of P. mirabilis local isolates compared with AM942759.1 P. mirabilis HI4320
Program: blastn, Iteration: 0, RID: 3X99KZ3B114, Database: n/a, 25 hits found.
Detection of O-antigen among P. Mirabilis Isolates using PCR Technique
The results of O-antigen detection showed that P. mirabilis isolates were positive to O-antigen and all tested isolates revealed the same PCR product on agarose gel (342 bp) as shown in the figure 3. Also, according to the product size of primers the results showed that the O- antigen of P. mirabilis was Wzy genes.
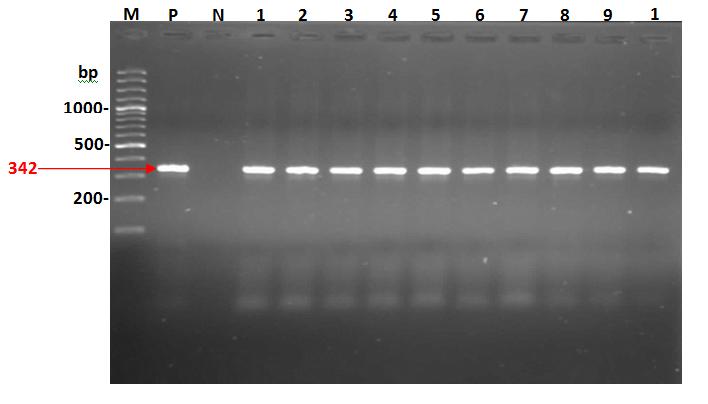
Sequencing of O-antigen of P. Mirabilis Isolates
The results of NCBI blast alignment in the table 10 showed that the local isolates of P. mirabilis had identity percentages with standard P. mirabilis strains. The high identity percentage of isolate No.1 was 83% with P. mirabilis strain Prk 20/57, P. mirabilis strain BB2000, P. mirabilis strain CCUG 4637 and P. mirabilis strain HI4320. The high identity percentage was 99% with each of P.
mirabilis strain CCUG 4637, P. mirabilis strain Prk 20/57, P. mirabilis strain Prk 50/57 and P. mirabilis strain BB2000 for isolates No. 2, 3, 4 and 10. The isolate No.6 revealed high identity percentage (99%) with P. mirabilis strain BB2000, while isolate No.8 had the same identity percentage with P. mirabilis strain Prk 20/57 and P. mirabilis strain HI4320. The high identity percentage of isolate No.9 was 87% with P. mirabilis BB2000.
| NCBI blast alignment | |||||||||||||||||
| No. of local | |||||||||||||||||
| isolates | P. mirabilis | P. mirabilis | P. mirabilis | P. mirabilis | P. mirabilis | ||||||||||||
| CCUG4637 | BB2000 | Prk20/57 | HI4320 | Prk50/57 | |||||||||||||
| Identity % | |||||||||||||||||
| 1 | 83 | 83 | 83 | 83 | 82 |
| 2 | 99 | 98 | 99 | 99 | 99 |
|---|---|---|---|---|---|
| 3 | 99 | 98 | 99 | 99 | 99 |
| 4 | 99 | 98 | 99 | 99 | 99 |
| 5 | 87 | 88 | 88 | 88 | 88 |
| 6 | 98 | 99 | 98 | 98 | 98 |
| 7 | 98 | 96 | 98 | 98 | 97 |
| 8 | 96 | 98 | 99 | 99 | 98 |
| 9 | 84 | 87 | 86 | 86 | 84 |
| 10 | 99 | 98 | 99 | 99 | 99 |
Table 12: NCBI blast alignment of O-antigen sequences of P. mirabilis local isolates.
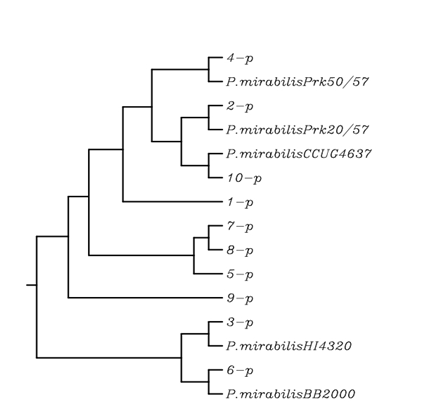
The phylogenetic tree analysis showed that the P. mirabilis local isolates were generally closely related to their respective type strain. The results revealed that there were six cluster groups of P. mirabilis local isolates 4, 1, [2 and 10], [7, 8 and 5], 9 and [3 and 6] as shown in the figure 4.
Discussion
Determine Serogroup of P. Mirabilis Isolates
After LPS extraction and purification as well as Acid degradation of LPS to purify O antigen of isolated P.
mirabilis, the serogroups of isolates were determined by using NMR spectroscopy. The results from the table 3 to the table 7 showed that the tested isolates were belonged to the serogroup O16, O18, O20, and O3, respectively.
The present results were in agreement with the study of Sidorczyk [22] who study the structure of O- polysaccharide and classified of P. mirabilis strain in Proteus serogroup O3, and he reported that O- polysaccharide O3 has branched tetrasaccharide O-unit containing one GlcA and two GalNAc residues in the main chain and an amide of GalA with L-lysine. Toukach [23] showed that a teichoic acid like OPS of P. mirabilis O16 has two phosphate substituents, ribitol 5-phosphate in the main chain and ethanolamine phosphate (EtNP) linked to a part of (~ 65%) of GlcNAc residues, and the similar result was also obtained by Toukach [24]. The OPS of P. mirabilis O18 has branched pentasaccharide phosphate O-unit composed of common sugar. This is only Proteus OPS that includes GlcNAc 1- phosphate and the only bacterial O antigen that contains choline phosphate [25]. The result about P. mirabilis O20 was agree with the result of Kondakova [26] who showed that the neutral of P. mirabilis O20 has a branched pentasaccharide O-unit consisting of common sugars only.
Genotyping of UreR gene of P. Mirabilis
Results in the figure 1 indicated that P. mirabilis isolates, which contain UreR gene, exhibited positive PCR products on gel which all of the isolates contained that gene. This result was similar to previous reports by Poore and Mobley [27]; they performed PCR to confirm the presence of urease gene in 10 P. mirabilis. Their results revealed that all isolates contain UreR. Two regulators of urease transcription have been characterized, UreR and H-NS (histone-like nucleoid structuring protein). UreR is a member of the AraC family of transcriptional regulators and contains both DNA- and urea-binding domains [28]. The UreR gene is transcribed in the opposite direction of UreDABCEFG; UreR binds the promoters of UreR and UreD [28]. Transcription of the structural genes of urease is urea-inducible [27]. UreR acts as a positive regulator of urease activity and stimulates expression of the urease genes in the presence of urea; H-NS is a negative regulator that represses UreR transcription. H-NS binds to the poly (A) tracts located in the intergenic region between UreR and UreD and inhibits transcription of UreR [27]. Dattelbaum concluded that the UreR is required for basal urease activity in the absence of urea, for induction of urease by urea, and for virulence of P. mirabilis in the urinary tract [15]. The results of UreR sequencing showed that the P. mirabilis local isolates were had a common ancestor with P. mirabilis standard strains as shown in the figure 2 and table 7, 8 and 9. These results were in agreement with Nicholson who found that the UreR gene of P. mirabilis had a cluster with P. mirabilis BB2000 and P. mirabilis HI4320 [29]. Also, he showed that identity percentage was 99% and 97% with P. mirabilis HI4320 and P. mirabilis BB2000, respectively [29]. This result was convergence with the current result, which obtained by NCBI blast where the identity percentage was ranged from (97.55%-99.44%) and (97.28%-98.60%) with P. mirabilis BB2000 and P. mirabilis HI4320, respectively. Nicholson demonstrated that the transcription of the urease operon is regulated by UreR. This gene lies upstream from the seven accessory or structural genes and is transcribed from its own promoter in the direction opposite that of the rest of the operon. Secondary regulation by a nitrogen regulatory system or catabolite repression does not appear to come into play. These data suggest that UreR governs the inducibility of P. mirabilis urease [29]. Despite the fact that Proteus and Providencia species produce inducible ureases, only P. mirabilis gene sequences were recognized on colony blots by the UreR probe hybridized under stringent conditions. This was surprising, because the structural genes of P. mirabilis and P. vulgaris ureases (UreABC) are highly conserved at the nucleotide level and ureases from both species are urea inducible [29].
Genotyping of O-antigen of P. Mirabilis
The results of O-antigen detection of P. mirabilis local isolates using PCR technique indicated that all local isolates were possessed the O-antigen gene and according to product size of the primer used for the detection of O- antigen, the location of O-antigen was between Wzx and Wzy genes. These results were similar to Wang who described the identification and molecular characterization of the putative Proteus O antigen synthesis gene cluster from 5 distinct serogroups and the development of a novel PCR-based method for the identification of respective Proteus isolates based on the amplification of unique Wzx and Wzy gene sequences. They suggested that the process for synthesis and translocation of O-antigens in these five Proteus serogroups was a Wzx/Wzy dependent process [10]. The NCBI blast sequencing results of the present study as shown in the tables [10] were in parallel with the results of Wang [10]. They showed that the sequencing alignment of putative Proteus O-antigen have the same O- antigen sequencing (100%) of P. mirabilis HI4320 and P.
mirabilis Prk 20/57, while the identity percentage was (99%) with P. mirabilis BB2000 and P. mirabilis Prk 50/57. Among the closely related genera Providencia, Proteus and Morganella, the O-antigen GC rich (OGC) has been characterized only in Proteus where it maps between the cpxA and secB genes [10].
Conclusions
NMR spectroscopy was considered a suitable method for determine the serogroup of P. mirabilis based on the determination of chemical structure of O-antigen repeating units. The UreR and O-antigen gene sequence comparison seems to be an appropriate method for inferring genetic relationships within the P. mirabilis isolates on a molecular basis. The UreR gene and O- antigen gene cluster sequences of P. mirabilis isolates allow rapidly a specific PCR assays to be designed.
References
-
Aquilini E (2012) Lipopolysaccharide (LPS) core biosynthesis in Proteus mirabilis. Ph.D. Environmental Microbiology and Biotechnology. Thesis. University College Barcelona. 285(15): 1-242.
-
Li X, Zhao H, Lockatell CV, Drachenberg CB, Johnson DE, et al. (2002) Visualization of Proteus mirabilis within the matrix of urease induced bladder stones during experimental urinary tract infection. J Infec Immun 70(1): 389-394.
-
Gonzalez G (2003) Proteus infections. Article by Gus Gonzalez, e Medicine.
-
Burall LS, Harro JM, Li X, Lockatell CV, Himpsl SD, et al. (2004) Proteus mirabilis genes that contribute to pathogenesis of urinary tract infection: Identification of 25 signature-tagged mutants attenuated at least 100-fold. J Infect and Immun 72(5): 2922-2938.
-
Heimer SR, Mobley TL (2001) Interaction of Proteus mirabilis urease apoenzyme and accessory proteins identified with yeast two-hybrid technology. J Bacteriol 183(4): 1423-1433.
-
Anna NK, Rafal F, Sofya NS, Alexander SS, Yuriy AK, et al (2005) Structure of a lactic acid ether containing and glycerol phosphate containing O-polysaccharide from _Proteus mirabilis_ O40. J Carbohyd Res 340: 1612-1617.
-
Perepelov A, Zabłotni A, Zych K, Senchenkova SN, Shashkov AS, et al (2004) Structure of the O- polysaccharide of Proteus mirabilis CCUG 10701 (OB) classified into a new Proteus serogroup, O74. J Carbohyd Res 339: 1395-1398.
-
Nazarenko EL, Crawford RJ, Ivanova EP (2011) The Structural Diversity of Carbohydrate Antigens of Selected Gram-Negative Marine Bacteria. J Mar Drugs 9: 1914-1954.
-
Raetz CRH, Whitfield C (2002) Lipopolysaccharide endotoxins. Annual Review of Biochemistry 71: 635- 700.
-
Wang Q, Torzewska A, Ruan X, Wang X, Rozalski A, et al. (2010) Molecular and Genetic Analyses of the Putative Proteus O Antigen Gene Locus. J Appl Environ Microbiol 76(16): 5471-5478.
-
Pearson MM, Sebaihia M, Churcher C, Quail MA, Seshasayee AS, et al. (2008) Complete genome sequence of uropathogenic Proteus mirabilis, a master of both adherence and motility. J Bacteriol 190(11): 4027-4037.
-
Al-Dawah MJ, Al-Hamadany AH, Al-Jarallah EM (2015) Study of some virulence factors of Proteus mirabilis isolated from urinary stone patients. J Bio Agriculture and Health 23(5): 85-95.
-
Al-Dawah MJ, Al-Hamadany AH, Al-Jarallah EM (2015) The role of lipopolysaccharide and O-antigen of Proteus mirabilis in urinary stones formation, in- vitro study. J Natural Sci Research 23(5): 47-52.
-
Micoli F, Rondini S, Gavini M, Pisoni I, Lanzilao L, et al. (2013) A scalable method for O-antigen purification applied to various Salmonella serovars. J Anal Biochem 434(1): 136-145.
-
Dattelbaum JD, Lockatell CV, Johnson DE, Mobley HLT (2003) UreR, the transcriptional activator of the Proteus mirabilis urease gene cluster, is required for urease activity and virulence in experimental urinary tract infection. J Infect Immun 71(2): 1026-1030.
-
Sanger F, Coulson AR (1975) A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J Mol Biol 94(3): 441-448.
-
Sambrook J, Rusell DW (2001) Molecular cloning: A laboratory manual. 3rd (Edn.) Cold Spring Harbor Laboratory Press, N.Y.
-
Adiguzel A, Ozkan H, Baris O, Bektas KI, Gulluce M, et al. (2009) Identification and characterization of thermophilic bacteria isolated from hot springs in Turkey. J Microbiol 79: 321-328.
-
Lee YK, Kwon KK, Cho KH, Kim HW, Park JH, et al. (2003) 16SrRNA. Culture and identification of bacteria from marine biofilms. J Microbiol 41(3): 183- 188.
-
Felsenstein J (2006) PHYLIP (phylogeny inference package). University of Washington, Seattle, USA.
-
Sidorczyk Z, Zych K, Toukach FV, Arbatsky NP, Zablotni A, et al. (2002) Structure of the O- polysaccharide and classification of Proteus mirabilis strain G1 in Proteus serogroup O3. Eur J Biochem 269(5): 1406-1412.
-
Toukach P, Arbatsky NP, Shashkov AS, Knirel Y, Zych K, et al. (2001) Structure of the O-specific polysaccharide of Proteus mirabilis O16 containing ethanolamine phosphate and ribitol phosphate. J Carbohydr Res 331: 213-218.
-
Toukach FV, Kondakova AN, Arbatsky NP, Senchenkova SN, Shashkov AS, et al. (2002) New structures of the O-specific polysaccharides of bacteria of the genus Proteus. Part 1. Phosphate- containing polysaccharides. Biochemistry J Carbohydr Res 67(2): 265-276.
-
Fudala R, Kondakova AN, Bednarska K, Shashkov AS, Knirel YA, et al. (2003) Structure and serological characterization of the O-antigen of Proteus mirabilis O18 with a phosphocholine-containing oligosaccharide phosphate-repeating unit. J Carbohydr Res 338(18): 1835-1842.
-
Kondakova AN, Fudala R, Bednarska K, Senchenkova S N, Knirel YA, et al. (2004) Structure of the neutral O- polysaccharide and biological activities of the lipopolysaccharide of Proteus mirabilis O20. J Carbohydr Res 339(3): 623-628.
-
Poore CA, Mobley HLT (2003) Differential regulation of the Proteus mirabilis urease gene cluster by UreR and H-NS. J Microbiol 149(12): 3383-3394.
-
D'Orazio SEF, Thomas V, Collins CM (1996) Activation of transcription at divergent urea-dependent promoters by the urease gene regulator UreR. J Mol Microbiol 21(3): 643-655.
-
Nicholson EB, Concaugh EA, Foxall PA, Island MD, Mobley HLT (1993) Proteus mirabilis urease: transcriptional regulation by UreR. J Bacteriol 175(2): 465-473.
-
Morsdorf G, Kaltwasser H (1990) Cloning of the genes encoding urease from Proteus vulgaris and sequencing of the structural genes. FEMS Microbiol Lett 66(1-3): 67-73.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions