Targeting Spike Protein: Modified Antibody for Broad-Spectrum Binding to Coronaviruses: An In Silico Study
An antibody-mediated immune response against the Spike protein is one of the potential ways to target the SARS-CoV-2 virus. The discovery of a highly effective vaccine or an antiviral drug that specifically targets the viral entry or its replication pathway, are commonly pursued as a way to combat COVID-19-like situation. The SARS-CoV-2 virus has RNA as its genetic material, which is known for its high mutation rate. This makes the efficiency and efficacy of the vaccine against it suboptimal later due to the continuous mutations in the epitope region of the virus. This was evident in the case of the spread of the Omicron (B.1.1.529) variant. Here we tested an in-silico approach towards finding an antibody that has the potential to recognise and bind against the Spike protein of the virus across most of the variants. We achieved this by altering the sequences of the VDJ region in the heavy and light chains of the natural human antibody against the Spike protein of the SARS-CoV-2 virus (Wuhan, PDB ID: 7BWJ). This was one of the first antibodies to be studied and published along with a crystal structure during mid- 2020, and known to have the ability to bind and neutralize the Wuhan isolate. Iterative substitutions were induced in the 7BWJ antibody sequence in different combinations. After 28 iterations, a potential antibody was found to bind across the variants with a comparatively higher docking score. This antibody also showed the potential to target the Spike protein of many members of Coronaviruses.
Introduction
Coronavirus disease 2019 (COVID-19) is caused by the virus SARS-CoV-2, which belongs to the family ‘Coronaviridae’ of the order ‘Nidovirales’ and has a single stranded linear RNA (+ssRNA) as its genome [1]. It is closely related to SARS-CoV-1, which caused a similar outbreak in East and Southeast Asia during 2002-2003. The whole genome sequence of SARS-CoV-2 is about 30 kb in length, and codes for 29 proteins of which 10 are the major proteins, including the Surface glycoprotein or S-protein (Spike protein). Spike protein is the largest protein of the virus consisting of 1273 amino acid long genomic sequences [2]. In the viral life cycle, one of the main steps for the virus is to bind with the host cell receptor and inject its genetic material into the host cell. In the case of SARS-CoV-2, the spike protein initiates this step by binding to the hACE2 receptors, which makes it a major target for the neutralizing antibodies (Fig. 1). This puts selection pressure on the virus to mutate the key amino acid residues of the Spike protein targeted by these antibodies [3]. The consequence was the emergence of many variants of the virus across the globe. These variants were able to escape antibody- mediated neutralization which was generated in response to the natural infection or the vaccination [4, 5]. However, there are constraints on the number of possible mutations of the epitope region of any pathogen. Considering this, one can design broad-spectrum antibodies to target several dominant epitopes of the virus [5]. Furthermore, it is possible to earmark the other members of the Coronaviridae by targeting the conserved and dominant epitope regions of the Spike protein either by using antibodies (cocktails) or multi-epitope vaccines. In this regard, multiple methods including natural/synthetic compounds, aptamers, peptidomimetics, affibodies to nanobodies; have been studied to target the Spike protein [6, 7, 8, 9, 10, 11, 12].
Antibodies are a group of proteins that are produced by our body’s immune cells - the plasma cells when they encounter foreign agents. The immune system generates diverse antibodies by changing its variable regions to increase the chances of finding a suitable match for recognizing and binding to different epitopes [13]. These changes are due to the presence of multiple copies of V, D and J segments in the immunoglobulin genes. These segments get reshuffled by the activity of the Recombination-Activating Genes (RAGs) to form different sequence combinations resulting in antibodies that can target innumerable antigens. After the sequence combination of an antibody is created that targets the specific antigen, it is preserved. When the corresponding antigen is detected, it triggers the production of that particular antibody by the respective B cells. This initiates antigen-antibody reactions, ultimately resulting in the elimination of the foreign particle from our body. The antibodies which can act in synergy having superior binding avidity with the escape mutants can be used as a cocktail for therapeutic purposes for diseases like COVID-19. Thus, a proper understanding of the interaction and the role of different amino acid sequences of the Spike protein and its cognate antibody helps in designing the therapeutic strategy. One can target different family members of the Coronavirus or the different variants of a particular virus using the knowledge of interacting amino acids of a receptor and their effect on the binding affinity of cognate antibody [14].
This study primarily focuses on the rearrangement of the sequences of heavy and light chains of the natural human antibody against the Spike protein of the Wuhan isolate for finding similar antibodies within the IgHV*3 family. 7BWJ, one of the initial antibodies against the Spike protein of the Wuhan-isolate with a crystal structure was chosen for the study. The antibody against the C-Terminal Domain of Proprotein convertase subtilisin/kexin type 9 (PCSK9) exhibited structural similarities, as well as partial sequence similarity with 7BWJ. Its sequence helped in deciding the kind of amino acid substitutions which has to be introduced in the 7BWJ variable region to find a broad spectrum binding antibody. We created mutation/substitution in the variable region of the 7BWJ and calculated the binding energies of the interactions with the Spike proteins. We compared the binding energies of 7BWJ with the Spike proteins of different Coronaviruses as well as with those of the variant of concerns of SARS-CoV-2. To introduce variations, amino acid substitutions were iteratively applied to the antibody’s variable region, by trying 28 distinct iterations (hereafter labelled as It1- It28) (Table 2). It28 antibody showed the potential to bind across different variants of the virus with promising docking results as compared with the original antibody which binds specifically only to the Wuhan isolate. The Serine to Cysteine residue substitution increased the docking efficiency more than other changes like Glycine to Isoleucine and Serine to Threonine (Table 2 & Table 3: It20- It28).
Our study offers insights into the impact of mutations on the interaction between the Spike protein of SARS- CoV-2 and the corresponding antibody. We demonstrate how these mutations can influence the binding affinity of the antibody towards the Spike protein. Additionally, we propose specific mutations within the variable region of the antibody that could potentially enable recognition of various family members of the SARS-CoV-2 virus and many of its variants. The engineered antibody holds promise in effectively hindering viral invasion by obstructing the interaction between the Spike protein and its receptor, Angiotensin Converting Enzyme 2 (ACE2) (Fig. 1). Through a comprehensive exploration of mutational possibilities, this study anticipates the development of an antibody with potential effectiveness against Coronavirus family members and many of the SARS-CoV-2 variants.
Methods
All the methods were carried out using various online and offline bioinformatics tools including, Cluspro, RCSB, PDBSum, NCBI BLAST, Bioedit 7.0, SAbPred, etc. for the structural prediction, sequence similarity check, docking, and evaluation of the docked proteins.
Retrieval of PDB Files
The PDB files of the Spike protein of SARS-CoV-2 virus (6VXX) (hereafter addressed as 6VXX), the antibody against the Spike protein of SARS-CoV-2 virus (7BWJ), the antibody against the c-terminal domain of PCSK9 (4K8R), etc. were retrieved from the RCSB Protein data bank (Table 1) [14].
| PDB ID | Name of the Protein | Crystal Structure- Publication Year |
|---|---|---|
| 6VXX | SARS-CoV-2 Spike glycoprotein Wuhan isolate | 2020 |
| 7BWJ | antibody against SARS-CoV-2 (Wuhan isolate) | 2020 |
| 4K8R | Antibody against the C-terminal domain of PCSK9 | 2014 |
| 5O7P | HER3 in complex with Fab of MF3178 | 2018 |
| 5UBY | Fab of anti- HIV-1 gp120 mAb 1A8 | 2018 |
| 6A67 | Crystal structure of influenza A virus H5 hemagglutinin globular head in complex with the Fab of the antibody FLD21.140 | 2018 |
| 6UVO | Structure of antibody 3G12 | 2019 |
| 3QHF | Crystal Structure of Fab del2D1 | 2011 |
| 1NLB | Crystal structure of anti-HCV mAb | 2003 |
| 8DLO | Cryo-EM structure of SARS-CoV-2 Spike protein | 2022 |
| Gamma (P.1) variant | ||
| 7WEV | SARS-COV-2 Beta (B.1.351) variant Spike protein | 2022 |
| 6U7H | Cryo-EM structure of HCoV-229E Spike protein | 2019 |
| 5I08 | Prefusion structure of Spike protein of HCoV-HKU1 isolate | 2016 |
| 7EKF | Spike protein of Alpha (B.1.1.7) variant of SARS-CoV-2 | 2021 |
| 5X5F | Spike protein of MERS-CoV | 2017 |
| 5X5b | Prefusion structure of Spike protein of SARS-CoV-1 | 2017 |
| 7W92 | SARS-CoV-2 Delta variant Spike protein | 2022 |
Table 1: List of the retrieved PDB structures from RCSB.
Retrieving Antibodies with Similar Sequences
The human antibody against the Spike protein of SARS- CoV-2 was derived from the RCSB having PDB ID 7BWJ and the sequence data of its heavy and light chain was retrieved from UniProt [15]. The sequence was then used for similarity search, which led to the discovery of the 4K8R, an antibody against the c-terminal domain of PCSK9 [16, 17].
Sequence Comparison of Antibodies
The sequence data of both antibodies 4K8R and 7BWJ were run on protein BLAST and it was seen that there was a higher number of gaps at the Cysteine and Serine residues.
Alteration of the Amino Acid Residues in Heavy and Light Chain of the Antibody
The alterations of the amino acids were made with the help of an offline bioinformatics tool (BioEdit version 7.0) and the changes are mentioned in Table 2.
| Name of the Antibody | Total Number Of Mutations | Mutations In The Heavy Chain | Mutations In Light Chain |
|---|---|---|---|
| It1 | 3 | S142C, T143A, S144C | - |
| It2 | 4 | E16M, T17A | P8S, S9M |
| It3 | 3 | S19P | A10M, M49A |
| It4 | 2 | E16H | C24A |
| It5 | 1 | S19A | - |
| It6 | 2 | S15C, 15*I | - |
| It7 | 2 | S7I | S2C |
| It8 | 4 | S15I, S19C | S2I, S9C |
| It9 | 4 | 19*I, S19C | S17C, 17*I |
| It10 | 8 | 19*I, S19C, S25I, S28C | S17C, 17*I, I20S, S21I |
| It11 | 7 | 19*I, S19C, S30C | S17C, 17*I, I20S, S21I |
| It12 | 12 | S7I, S15C, 19*I, S19C, S25I, S28C | S2C, S9I, S17C, 17*I, I20S, S21I |
| It13 | 12 | S7I, S15C, 19*I, S19C, S25I, S28C | S2C, S9I, S17C, 17*I, I20S, S21I |
| It14 | 13 | S7I, S15C, 19*I, S19C, S25I, S28C, S125C | S2C, S9I, S17C, 17*I, I20S, S21I |
| It15 | 19 | S7I, S15C, 19*I, S19C, S25I, S28C, S75C, S125C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, C138S |
| It16 | 20 | S7I, S15C, 19*I, S19C, S25I, S28C, S63C, S75C, S125C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, C138S |
| It17 | 23 | S7I, S15C, 19*I, S19C, S25I, S28C, P41F, S63C, S75C, S125C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, P61F, F64P, S91I, S92C, C138S |
| It18 | 18 | S7I, S15C, 19*I, S19C, S25I, S28C, S63C, S75C, S125C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, , C138S |
| It19 | 20 | S7I, S15C, 19*I, S19C,S25I, S28C, S63C, S75C, S125C | S2C, S9I, S17C, 17*I, I20S,S21I, S26I, S27C, S91I, S92C, C138S |
| It20 | 19 | S7I, S15C, 19*I, S19C, S25I, S28C,S75C, S125C | S2C, S9I, S17C, 17*I, I20S,S21I, S26I, S27C, S91I, S92C, C138S |
| It21 | 22 | S7I, S15C, 19*I, S19C, S25I, S28C, S63C, S75C,S125C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, P61F,F64P, S91I, S92C, C138S |
| It22 | 21 | S7I, S15C, 19*I, S19C, S25I, S28C, S75C, S125C,S127C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, S96C, C138S |
| It23 | 22 | S7I, S15C, 19*I, S19C, S25I, S28C, S30C, S75C, S125C, S127C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, S96C, C138S |
| It24 | 23 | S7I, S15C, 19*I, S19C, S25I, S28C, S30C, S75C, S125C, S127C, S192C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, S96C, C138S |
| It25 | 24 | S7I, S15C, 19*I, S19C, S25I, S28C, S30C, S75C, S125C, S127C, S192C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, S96C, C138S, S180C |
| It26 | 23 | S7I, S15C, 19*I, S19C, S25I, S28C, S75C, S125C, S127C, C208S | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, S96C, C138S, C197S |
| It27 | 21 | S7I, S15C, 19*I, S19C, S25I, S28C, S75C, C208S | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, S96C, C138S, C197S |
| It28 | 23 | S7I, S15C, 19*I, S19C,S25I, S28C, S75A, K76T, S125C, S127C | S2C, S9I, S17C, 17*I, I20S, S21I, S26I, S27C, S91I, S92C, S96C, S118C, C138S |
Table 2: List of mutations induced in the variable region of the 7BWJ in each iteration.
* depicts insertion of the amino acid Table 2: List of mutations induced in the variable region of the 7BWJ in each iteration.
Prediction of the Modified Antibody
The light chain and heavy chain sequences of the resulting antibody were submitted to an online antibody prediction tool to develop the antibody with the input sequences and the PDB ID of the same was retrieved [18].
Docking with Spike Protein
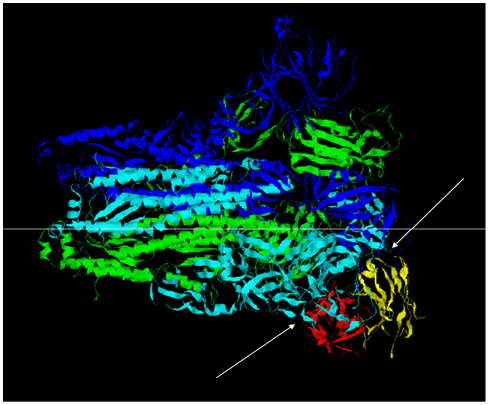
The predicted antibody was docked with the Spike protein of SARS-CoV-2 (6VXX) using the Cluspro protein- protein docking tool [19]. The docking results suggested a better binding to the spike protein. This binding was observed at a slightly different epitope than the docking of the natural antibody with the spike protein of the Wuhan isolate (Figure 2).
Evaluation of the Antibody Docking and Interactions
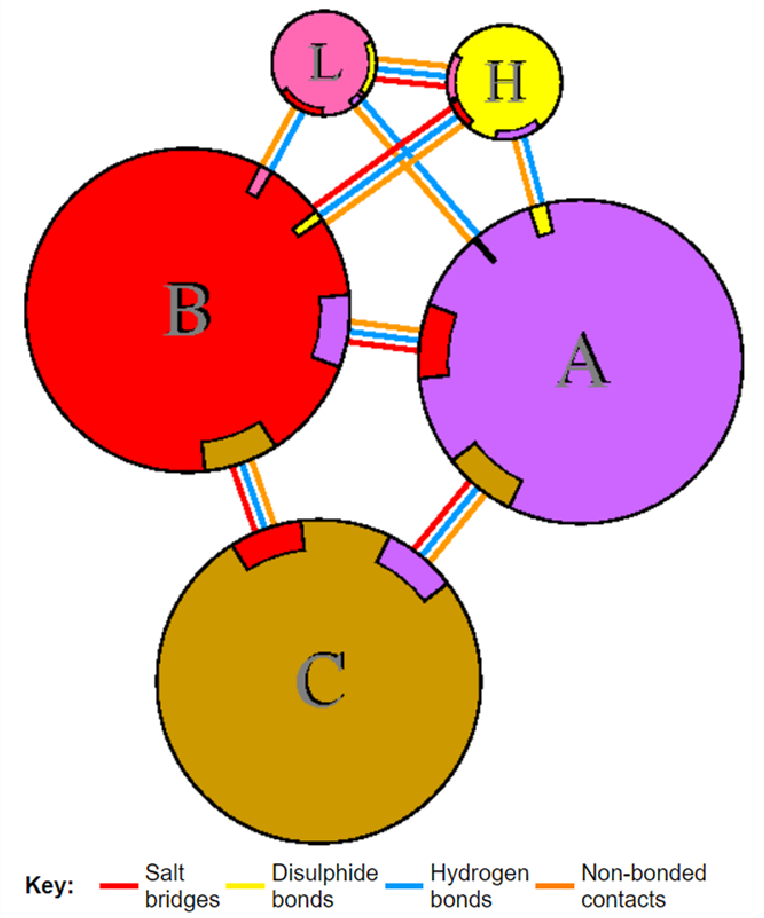
To confirm the binding efficiency, the retrieved PDB files after docking were studied for the various interactions between the molecules. We analysed the presence of different hydrogen bonds, covalent bonds, and disulphide bridges being formed between the docked molecules. The docked files were submitted to an online tool developed by the EMBL group (PDBsum- http://www.ebi.ac.uk/thornton- srv/databases/pdbsum/Generate.html) for the analysis of different interactions.
Results
Comparative Analysis of Docking Score of Different Control Sets
In the initial analysis, we had to find the docking score of the human antibody against the Spike protein of the Wuhan isolate of SARS-CoV-2, with which all the other results were compared. To establish the specificity of the docking, we did a test run of the Spike protein with various antibodies retrieved from the RCSB including 4K8R, 5O7P, 5UBY, 6A67, and 6UVO. Upon docking with the viral Spike proteins, the antibodies 5O7P and 4K8R showed a higher score of docking than the normal antibody whereas all the other antibodies retrieved, showed a less score (Table 3). This included antibodies against Spike protein of SARS-CoV-1 and MERS, Fab of anti-HIV gp120, Fab (FLD21.140) against Influenza A, and monoclonal antibody against HER3. These were considered a set of negative controls because these antibodies were among the top hits in sequence similarity search.
| File name | Number of amino acids participating in docking | Binding energy | Bond formation |
|---|---|---|---|
| 4K8R vs 6VXX | No specific interactions | -1031.1 | No |
| 7BWJ vs6VXX | 37 | -954.6 | yes |
| It1 vs 6VXX | 50 | -773.6 | yes |
| It2 vs 6VXX | 38 | -765.2 | yes |
| It3 vs 6VXX | 47 | -779.2 | yes |
| It4 vs 6VXX | 42 | -806.1 | yes |
| It5 vs 6VXX | 52 | -775.2 | yes |
| It6 vs 6VXX | 94 | -803.3 | yes |
| It7 vs 6VXX | 45 | -772.2 | yes |
| It8 vs 6VXX | 46 | -829.3 | yes |
| It9 vs 6VXX | 98 | -825.7 | yes |
| It10 vs 6VXX | 81 | -829.7 | yes |
| It11 vs 6VXX | 94 | -829.8 | yes |
| It12 vs 6VXX | 68 | -831.4 | yes |
| It13 vs 6VXX | 49 | -830.9 | yes |
| It14 vs 6VXX | 52 | -832 | yes |
| It15 vs 6VXX | 35 | -898.8 | yes |
| It16 vs 6VXX | 40 | -884.6 | yes |
| It17 vs 6VXX | 56 | -897 | yes |
| It18 vs 6VXX | 57 | -877.3 | yes |
| It19 vs 6VXX | 43 | -884 | yes |
| It20 vs 6VXX | 35 | -898.8 | yes |
| It21 vs 6VXX | 38 | -884.6 | yes |
| It22 vs 6VXX | 35 | -934.7 | yes |
| It23 vs 6VXX | 31 | -934.9 | yes |
| It24 vs 6VXX | 31 | -934.9 | yes |
| It25 vs 6VXX | 31 | -934.9 | yes |
| It26 vs 6VXX | 35 | -934.7 | yes |
| It27 vs 6VXX | 32 | -922.9 | yes |
| It28 vs 6VXX | 36 | -982.2 | yes |
Sequence Similarity between 7BWJ and 4K8R Guided the Selection of Amino Acids to Be Substituted
The further task was to find an antibody which can be taken as a reference to induce substitutions in the heavy and light chain region of the natural human antibody - 7BWJ. Hence, the light and heavy chain sequences of the
7BWJ were set for similarity searches in RCSB that resulted in an antibody against the c-terminal domain of PCSK9 (PDB ID- 4K8R) as one of the top hits. Using BLASTp, we found the gaps and similarities between the sequences of heavy and light chains of the 7BWJ and 4K8R antibodies. The mismatch in the amino acid sequences of both these antibodies suggested the kind of substitutions to be made in the 7BWJ for a broader binding ability.
Docking of the Modified Antibody with the Spike Protein
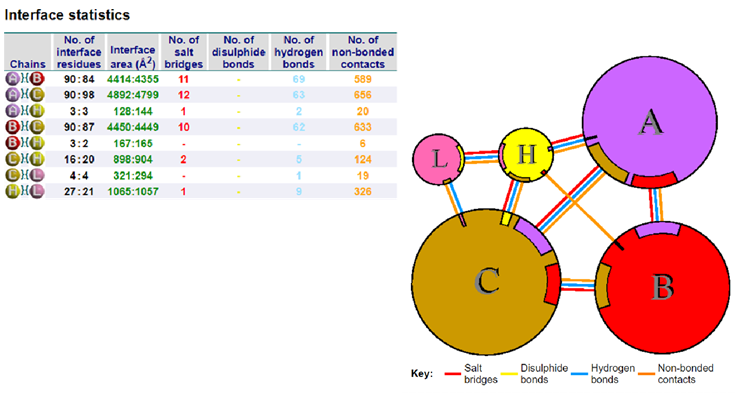
When the modified antibody was docked with the Spike protein of the SARS-CoV- 2, it was found to bind a slightly different epitope region of the Spike protein than the epitopes recognised by the 7BWJ. The higher docking score suggested its better binding ability than the 7BWJ (Table 3) (Figure 2,3, Supp. Figure 1,2). To find the breadth of the efficacy of the modified antibody, it was docked with the Spike proteins of Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2), and Omicron (B.1.1.529) variants. We found slightly better binding in the case of Omicron and Delta variants (Table 3; Figure 4-9). Same epitope was recognized by the It28 antibody of these variants. Likewise, Gamma variant possesses a large number of mutations, yet recognised by the modified antibody (Supp. Fig. 4). This suggests that the antibody is targeting the conserved region or the backbone region of the Spike protein where the mutations don’t seem to interfere with the binding efficiency of the modified antibody (Figure 6,9,12).
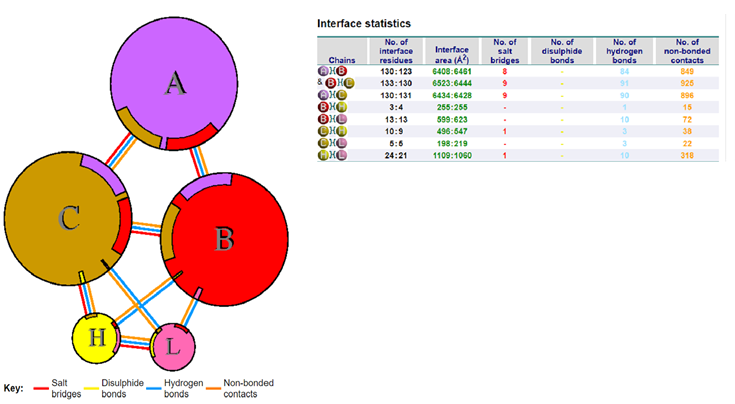
The Docking Comparison of the 7 Family Members of the Coronavirus
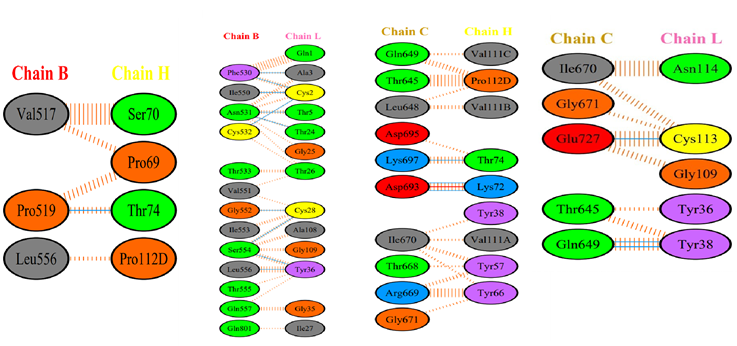
We analysed whether the modified antibody was able to recognise the epitopes of different family members of the Coronavirus, and how specific the docking was when compared with that of the Wuhan isolate. We observed that the antibody was able to bind with 5 out of 7 family members including the SARS-CoV-2 (Wuhan isolate) (Figure 3), HCoV- HKU1 (Figure 10), SARS-CoV-1, HCoV-OC43, HCoV-229E (Supp. Figure 9,10,11).
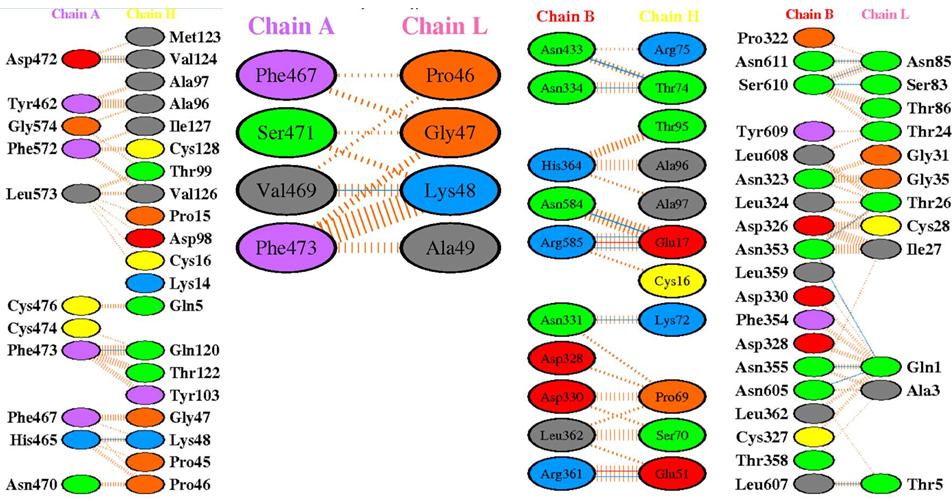
It28 vs. SARS-CoV-1: The docking with the SARS-CoV-1 Spike protein was carried out after retrieving the PDB file from RCSB with a score of -857.5 (Table 2). To further see if the docking was specific, the docked files were subjected to PDBsum for the analysis of participating amino acids and it was seen that chain C of the Spike protein and the light and heavy chains of the antibody were interacting. This suggests that the antibody could recognise the epitopes of the SARS- CoV-1, but with a lower efficiency as compared to the Wuhan isolate.
It28 vs. MERS-CoV: There was no specific interaction seen in the case of docking the modified antibody with the MERS-CoV Spike protein. This is probably because of the high dissimilarity of the sequences in its Spike protein as compared with the Spike protein of the SARS-CoV-2. This indicates the specificity of the antibody towards specific recognition sites or sequences.
It28 vs. HCoV-HKU1: When the Spike protein of HCoV-HKU1 was docked with the modified antibody; we obtained a much higher docking score. It formed 2 salt bridges between the C chain of the Spike protein and the light chain of the antibody. It was also observed that few amino acids were common in bond formation as seen in the case of Omicron’s (B.1.1.529) Spike protein binding with the modified antibody. The binding of the modified antibody with the HCoV-HKU1 Spike protein had interactions involving the amino acids around the 470-490 region embedded in chain A, which was not seen across the other family members or variants during docking (Figure 11).
It28 vs. HCoV-OC43: When the modified antibody was docked with the Spike protein of the HCoV- OC43 virus, there was a slight increase in the docking score as compared with the SARS- CoV-2 Spike protein.
It28 vs. HCoV-229E: There was a significant interaction between the B and C chains of the Spike protein with the heavy chain of the antibody, with a greater number of participating amino acids forming non-bonded contacts. However, the docking score was slightly less when compared to the docking score of the It28 antibody with the Spike protein of the Wuhan isolate.
It28 vs. NL63: As seen in the case with the MERS-CoV, there was no specific docking observed in the case of the NL63 Spike protein with the modified antibody. This might indicate the sequence similarity is not the only factor but the final structure of the Spike protein might have a role in determining the sequences projected and hidden away from the epitope regions, which are targeted and recognized by the antibodies.
The Comparative Study of Docking with Different Variants of the SARS-CoV-2 Virus: The main objective of this study was to find a suitable antibody that can bind and hence probably neutralise SARS-CoV-2 virus and its variants. The Omicron (B.1.1.529) variant possessing the highest number of mutations has a higher docking score. The antibody has also shown efficient docking and the participating amino acids showed sufficient numbers of possible hydrogen bonds with different variants which were not observed in the case of natural antibody. (Figures 4 & ,7, Supp. Figure 3).
It28 vs Beta (B.1.351) Variant: The modified antibody was subjected to dock with the Spike protein of the Beta (B.1.351) variant [20]. It was noted that the docking score of It28 with Beta variant was higher as compared to the 7BWJ antibody. This suggests that the efficiency of the modified antibody to bind with this variant is better than the natural antibody. Additionally, the interactions between the B chain of the Spike protein and the light chain of the antibody had a hydrogen bond between the 256th Serine residue and the 55th Tyrosine residue respectively. This was also observed in a few other variants where the B chain of the Spike protein is being recognized. This suggests that this is one of the main residues in the epitope region when the antibody targets the B chain across the variants (Supp. Figure 6-8).
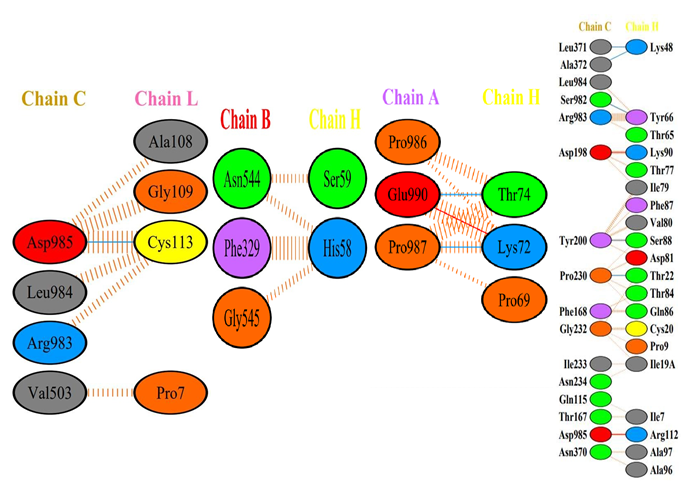
It28 vs Delta Variant: The result of docking between the It28 vs Delta variant reflected the interaction in a better way (Figure 8,9). There was a salt bridge formation within the glutamate 990 residue of the Spike’s A chain and the lysine 72 residue of the heavy chain of the It28 (Figure 9). Similar salt bridge was seen in the interaction between the modified antibody with the HCoV-HKU1 Spike protein which has a high docking score.
It28 vs Omicron (B.1.1.529) Variant: Surprisingly, the Omicron (B.1.1.529) variant which is known to possess the highest number of mutations in the Spike protein, has a high docking score. We observed a double hydrogen bond between the Aspartate 693 and Lysine 72 of the Spike’s C chain and heavy chain of the antibody, and glutamate 649 and tyrosine 38 of the C chain and light chain of the antibody respectively [21]. A salt bridge formation was also observed between the Aspartate 693 and Lysine72, suggesting an efficient recognition by the modified antibody to the epitope (Figure 5,6).
It28 vs Gamma (P.1) Variant: Docking with the Gamma (P.1) variant’s Spike protein showed the highest docking score (Table-2) [22]. Tyrosine 55 of the light chain and Lysine 66 of the heavy chain of the modified antibody formed a hydrogen bond with the chain A of the Spike protein of the Gamma (P.1) variant (Supp. Figure 3,4,5).
It28 vs Alpha (B.1.1.7) Variant: The docking with the Alpha (B.1.1.7) variant showed a null result. This might be due to the E484K mutation seen in this variant, and could be a reason for the inefficient docking. Though in other cases, the epitope recognition is mainly among the B and C chain, the E484K mutation lies in the A chain of the Spike protein and is suspected to have an effect in denying the antibody binding.
Evaluation of Participating Amino Acids and Number of Bonds after Docking
For the evaluation of the type of bond formation between the amino acids that participated in the docking of the antibody with the Spike protein; the docked file was analysed through PDBsum. The hydrogen bond formation, salt bridge formation, and non-bonded contacts between the participating amino acids of the light and heavy chains with the chains of the Spike protein of various variants were analysed. It was observed that the amino acids between the 48th and 73rd residues of heavy and light chains were seen to be more actively forming hydrogen bonds upon substitution made in the It28. The 72nd Lysine and the 51st Glutamic acid residue in the heavy chain of the modified antibody actively participated in multiple bond formation including hydrogen bond and a salt bridge across the variants especially with Omicron, Delta, and Gamma and the HKU1 virus.
| Docking energy | Chains interacting | Presence of hydrogen bond | Presence of a salt bridge | Total number of interactions | |
|---|---|---|---|---|---|
| 7BWJ vs 6VXX | -854.6 | C and L chains, C and H chain | 8 | 0 | 157 |
| 7BWJ vs 7TB4 | Nil | Nil | Nil | Nil | Nil |
| 7BWJ vs 7TB4 | Nil | Nil | Nil | Nil | Nil |
| 7BWJ vs 7WEV | -821.2 | B and H, B and L | 1 | 0 | 37 |
| It28 vs 6VXX | -982.2 | C and L | 9 | 0 | 177 |
| It28 vs 7TB4 | -1021 | A and H, B and H, C and L, C and H | 8 | 3 | 180 |
| It28 vs 7WEV | -880.1 | B and H, B and L | 15 | 0 | 179 |
| It28 vs 7EKF | Nil | Nil | Nil | Nil | Nil |
| It28 vs 8DLO | -1057.5 | A and H, A and L | 13 | 0 | 207 |
| It28 vs 7VHH | -880.5 | C and H, C and L, Band H, B, and L | 17 | 1 | 165 |
Table 4: The docking scores and the bonded and non-bonded contacts between the interacting antibodies and Spike protein.
Discussion
Spike protein of the SARS-CoV-2 virus is primarily targeted by the natural host immune response. This is due to its antigenicity and its major role in the entry of the virus inside the host cell. The humoral immune response is critical in safeguarding the host during a viral infection. The antibodies generated against different antigens of the virus prevent viral entry and its propagation in the host. Antibodies against the Spike protein of the SARS-CoV-2 virus which can bind and neutralize them can also prevent the entry of the virus into the host cell [23]. SARS-CoV-2 virus has a higher mutation rate due to RNA being its genetic material. This could provide better adaptability to the virus by mutating the crucial amino acid residues which are targeted by the host’s adaptive immunity. This was observed in the case of the SARS-CoV-2 lead pandemic, where we saw several variants of the virus emerging and capable of immune escape. The studies based on computational or structural analysis and the affinity and avidity test from patients plasma have revealed the inability of natural antibodies to cross-protect from the virus variants. This was particularly evident in the B.1.1.529 variant possessing a large number of mutations [15, 18, 22, 24].
During the early pandemic, lack of specific treatment for COVID-19 posed serious challenges worldwide. Development of a therapeutic strategy for the emerging variants and to manage future challenges for other such situations is needed. A therapeutic approach based on antibodies is an area of intense research in recent times [25, 26]. Antibodies can recognize a vast range of antigenic repertoires. The Complementarity Determining Regions 1-3 (CDR 1-3) within the Variable Fragment (Fv) of the antibodies play a crucial role in conferring this diversity. Several laboratories have tested single- domain antibodies (nanobodies) that utilize CDR sequences capable of binding to the Receptor-Binding Domain (RBD) and exhibiting virus-neutralizing capacity [27, 28].
By changing the amino acid sequence of the antibody which was naturally generated against the original viral isolate; we conducted this study to determine whether it is possible to predict an antibody capable of binding to a wide range of SARS-CoV-2 variants and the members of the Coronavirus family. After multiple iterative changes in the amino acid sequence, we found such an antibody capable of binding to the Spike protein of most of the viral variants of SARS-CoV-2 as well as of other Coronaviruses. However, there were some exceptions as seen during the docking of It28 with the Spike protein of the Alpha (B.1.1.7) variant. The E484K mutation is most likely the reason for the inability of 7BWJ and It28 to bind with this variant [29]. It was noted that during docking with other variants, most of the target region seems to lie in the B and/or C chain rather than the A chain. There might be a slight chance that the projection of the lysine (E484K) masks the other possible interacting regions hence antibody could not possibly bind with the Alpha variant. We observed a few specific interactions during the docking of this antibody with the Spike proteins of the 7 family members of the Coronavirus. HCoV-HKU1 Spike protein docking with the It28 resulted in the formation of salt bridges and double hydrogen bonds. Similarly, Delta and Omicron variants were also seen to have the presence of a salt bridge or interaction through a double hydrogen bond. The substitutions in the It28 antibody, which are believed to be vital in the interactions include S28C, S127C, and S75A in the heavy chain; and S17C, S91I and Isoleucine insertion at the 17th position in the light chain. Combination of these substitutions probably is important for its broader recognition abilities and interactions.
Conclusion
This work is valuable as the antibody which we have predicted could prevent the infection by the SARS-CoV-2 virus variants, given that the iterated antibody from our study leads to an effective neutralization. We predicted a potent antibody that can target many members of the Coronaviridae as well as SARS-CoV-2 variants. The change in amino acid sequence can lead to altered interaction dynamics between an antibody to its receptor. The approach can be applied as a quick therapeutic method to tackle future challenges posed by a virus with high mutation rate. We need to see whether the antibody harboring the proposed substitutions could be expressed and folded in its host in a natural condition; as some of the mutations may be deleterious [3]. This study has limitations in testing the proposed antibody by the in vitro and in vivo experimental methods and needs validation. However, besides the proposed antibody, our study can guide methodologically in designing antibodies with a broad range of targets to tackle the challenges posed by the continuously evolving SARS-CoV-2 virus.
Conflicts of Interest: Authors declare no conflict of interest.
Acknowledgements: Authors are thankful to the Translational Health Science and Technology Institute, Faridabad for the support.
References
-
Huang Y, Yang C, Xu X, Xu W, Liu S (2020) Structural and functional properties of SARS- CoV-2 Spike protein: potential antivirus drug development for COVID-19. Acta Pharmacol Sinica 41: 1141-1149.
-
Pal M, Berhanu G, Desalegn C, Kandi V (2020) Severe Acute Respiratory Syndrome Coronavirus-2 (SARS- CoV-2): An Update. Cureus 12: e7423.
-
Starr TN, Greaney AJ, Hilton SK, King NP, Veesler D, et al. (2020) Deep Mutational Scanning of SARS- CoV- 2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 182(5): 1295-1310.
-
Greaney AJ, Starr TN, Gilchuk P, Carnahan RH, Crowe Jr JE, et al. (2021) Complete Mapping of Mutations to the SARS-CoV-2 Spike Receptor-Binding Domain that Escape Antibody Recognition. Cell Host & Microbe 29(1): 44-57.
-
Sharma D, Rawat P, Janakiraman V, Gromiha MM (2022) Elucidating important structural features for the binding affinity of Spike - SARS-COV -2 neutralizing antibody complexes. Proteins 90 (3): 824-834.
-
Bharathi M, Sivamaruthi BS, Kesika P, Thangaleela S, Chaiyasut C (2022) In Silico Screening of Bioactive Compounds of Representative Seaweeds to Inhibit SARS- CoV-2 ACE2-Bound Omicron B.1.1.529 Spike Protein Trimer. Marine Drugs 20(2): 148.
-
Du W, Jiang P, Li Q, Wen H, Zheng M, et al. (2023) Novel Affibody Molecules Specifically Bind to SARS-CoV-2 Spike Protein and Efficiently Neutralize Delta and Omicron Variants. Microbiol Spectr 11(1): e03562- e03522.
-
Lin Y-C, Chen W-Y, Hwu E-T, Hu W-P (2022) In-Silico Selection of Aptamer Targeting SARS-CoV-2 Spike Protein. IJMS 23(10): 5810.
-
Sabzian-Molaei F, Khalili MAN, Sabzian-Molaei M, Shahsavarani H, Pour AF, et al. (2022) Urtica dioica Agglutinin: A plant protein candidate for inhibition of SARS-COV-2 receptor- binding domain for control of Covid19 Infection. PLoS ONE 17: e0268156.
-
Singh R, Kumar A, Rane JS, Khan R, Tripathi G, et al. (2022) Arylcoumarin perturbs SARS-CoV-2 pathogenesis by targeting the S-protein/ACE2 interaction. Sci Rep 12: 17038.
-
Ibrahim MT, Tao P (2022) Computational investigation of peptidomimetics as potential inhibitors of SARS- CoV-2 Spike protein. Journal of Biomolecular Structure and Dynamics 41(15): 7144-7157.
-
Verkhivker G (2022) Structural and Computational Studies of the SARS-CoV-2 Spike Protein Binding Mechanisms with Nanobodies: From Structure and Dynamics to Avidity-Driven Nanobody Engineering. IJMS 23(6): 2928.
-
Yang L, Li J, Guo S, Hou C, Liao C, et al. (2021) SARS-CoV-2 Variants, RBD Mutations, Binding Affinity, and Antibody Escape. IJMS 22(22): 12114.
-
Nussenzweig MC, Alt FW (2004) Antibody diversity: one enzyme to rule them all. _Nat Med_ 10: 1304-1305.
-
Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, et al. (2000) The Protein Data Bank. Nucleic Acids Research 28(1): 235-242.
-
Ju B, Zhang Q, Ge J, Ge J, Wang R, Sun J, et al. (2020) Human neutralizing antibodies elicited by SARS- CoV-2 infection. Nature 584: 115-119.
-
Schiele F, Park J, Redemann N, Luippold G, Nar H (2014) An Antibody against the C-Terminal Domain of PCSK9 Lowers LDL Cholesterol Levels In Vivo. Journal of Molecular Biology 426(4): 843-852.
-
Dunbar J, Krawczyk K, Leem J, Marks C, Nowak J, et al. (2016) SAbPred: a structure-based antibody prediction server. Nucleic Acids Res 44(W1): W474-W478.
-
Desta IT, Porter KA, Xia B, Kozakov D, Vajda S (2020) Performance and Its Limits in Rigid Body Protein-Protein Docking. Structure 28(9): 1071-1081.
-
Wang Y, Xu C, Wang Y, Hong Q, Zhang C, et al. (2021) Conformational dynamics of the Beta and Kappa SARS- CoV-2 Spike proteins and their complexes with ACE2 receptor revealed by cryo-EM. Nat Commun 12: 7345.
-
Cerutti G, Guo Y, Liu L, Liu L, Zhang Z, et al. (2022) Cryo- EM structure of the SARS-CoV-2 Omicron Spike. Cell Reports 38(9): 110428.
-
Mannar D, Saville JW, Sun Z, Zhu X, Marti MM, et al. (2022) SARS-CoV-2 variants of concern: Spike protein mutational analysis and epitope for broad neutralization. Nat Commun 13: 4696.
-
Wang C, Li W, Drabek D, Okba NMA, van Haperen R, et al. (2020) A human monoclonal antibody blocking SARS- CoV-2 infection. Nat Commun 11: 2251.
-
Villas-Boas GR, Rescia VC, Paes MM, Lavorato SN, de Magalhães-Filho MF, et al. (2020) The New Coronavirus (SARS- CoV-2): A Comprehensive Review on Immunity and the Application of Bioinformatics and Molecular Modeling to the Discovery of Potential Anti-SARS- CoV-2 Agents. Molecules 25(18): 4086.
-
Griffin MP, Yuan Y, Takas T, Domachowske JB, Madhi SA, et al. (2020) Single-Dose Nirsevimab for Prevention of RSV in Preterm Infants. N Engl J Med 383: 415-425.
-
Mulangu S, Dodd LE, Davey RT, Mbaya OT, Proschan M, et al. (2019) A Randomized, Controlled Trial of Ebola Virus Disease Therapeutics. N Engl J Med 381: 2293-2303.
-
Yang J, Zhang Z, Yang F, Zhang H, Wu H, et al. (2021) Computational design and modeling of nanobodies toward SARS-CoV-2 receptor binding domain. Chem Biol Drug Des 98: 1-18.
-
Wagner TR, Ostertag E, Kaiser PD, Gramlich M, Ruetalo N, et al. (2021) NeutrobodyPlex-monitoring SARS-CoV-2 neutralizing immune responses using nanobodies. EMBO Reports 22(5): e52325.
-
Jangra S, Ye C, Rathnasinghe R, Stadlbauer D, PVI study group, et al. (2021) The E484K mutation in the SARS- CoV-2 Spike protein reduces but does not abolish neutralizing activity of human convalescent and post- vaccination sera. Infectious Diseases (except HIV/AIDS).
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions