Comparative Investigation of In Vitro Genotoxicity of Pro-Mutagens Using Human-Derived Hepatoma HepG2 Cells with Human and Rat Liver S9
The genotoxicity of pro-mutagens has been traditionally detected by test procedures with metabolically incompetent cells requiring addition of exogenous activation mixtures, such as rat liver S9. HepG2 cells established from huma hepatoma in 1979 have metabolic activation ability. For this characteristic, HepG2 cells were tried to be used in the genotoxicity test. Since metabolic activation ability in HepG2 cells is not the same as that in exogenous activation mixtures, it is necessary to ascertain whether HepG2 cells could be used to detect the genotoxicity of various kinds of pro-mutagens and to compare the detecting ability between HepG2 cells and metabolically incompetent cells under exogenous activation mixtures. The ability of HepG2 cells to detect the genotoxicity of 38 pro-mutagens was studied to compare that of metabolically incompetent human lymphoblastoid WTK1 cells under human and rat S9’s. Present results showed that the ability of HepG2 cells to detect the genotoxicity of pro-mutagens parallels qualitatively to that of WTK1 cells under human liver S9 and cytochrome (CYP) induced rat liver S9 even if the existence of species difference in CYP isoforms. Therefore, HepG2 cells could be usefully used to detect the gentoxicity of pro-mutagens for the purpose of the evaluation of the genotoxic risk for humans.
Abbreviations
CYP: Cytochrome; BF: Benzoflavone.
Introduction
A main purpose of a genotoxicity test includes to detect a mutagen and to predict a risk in humans. Up to date, the prediction of carcinogenicity from mutagenicity is done broadly, which plays an important role in the development of new medicines. For this purpose, data obtained using organisms other than human are usually used. An S9 fraction prepared from the liver of the rat pre-treated with phenobarbital/5,6-benzoflavone (PB/BF) or Aroclor 1254 is generally used as a model of metabolic activation system in the liver of humans [1, 2]. However, species difference between humans and rats cannot be ignored because of difference in P450 molecules. Furthermore, the possibility cannot be denied that metabolic activation by cytochrome (CYP)
induced rat liver S9 overestimates metabolic activation in the human liver. Our previous study was therefore undertaken to determine the genotoxicity of 38 pro-mutagens which are preferentially activated by different P450 isozymes using the comet assay with human lymphoblastoid WTK1 cells under human and rat liver S9’s [3]. It was shown that there were no appreciable qualitative differences in the ability to detect pro-mutagen between human liver S9 and CYP-induced rat liver S9 and that conventionally used CYP-induced rat liver S9 fraction is useful for the evaluation of the genotoxic risk for humans in spite of the species difference in CYP isoforms. HepG2 cells having metabolic activation ability were established from human hepatoma in 1979 by Aden, et al. [4, 5]. For this characteristic, HepG2 cells were tried to be used in the genotoxicity test [5]. Since metabolic activation ability in HepG2 cells is not the same as that in human liver, it is necessary to ascertain whether HepG2 cells could be used to detect the genotoxicity of various kinds of pro-mutagens and to compare the detecting ability between metabolically incompetent cells under exogenous activation mixtures (human and rat S9’s) and HepG2 cells. Therefore, we studied the detecting ability of HepG2 cells for the genotoxicity of 38 pr-omutagens same to those have been studied under human and rat liver S9’s [3].
Materials and Methods
Chemicals, Cells and Medium
Table 1 lists the pro-mutagens tested, their abbreviation, and CAS number. Human hepatoma (HepG2) cells, obtained from Riken Cell Bank, Tokyo, were grown in 1:1 mixture medium of Dulbecco MEM and Ham’s F12 supplemented with 10% fetal bovine serum (HyClone Laboratories, Inc., U.S.A.). All media were purchased from Nissui Pharmaceutical Inc., Tokyo. Cells were grown at 37°C in a humidified atmosphere containing 5% CO2.
| Promutagens | Abbreviation | CAS number | Sourcea | Vehicleb |
|---|---|---|---|---|
| Aromatic hydrocarbons | ||||
| Benzene | 71-43-2 | W | D | |
| Benz[a]anthrathene | B[a]A | 56-55-3 | T | D |
| Benzo[a]pyrene | B[a]P | 50-32-8 | S | D |
| Phenanthrene | 85-01-8 | T | D | |
| Pyrene | 129-00-0 | T | D | |
| Aromatic amines | ||||
| 2-Aminoanthracene | 2AA | 613-13-8 | W | D |
| Aniline | 62-53-3 | W | D | |
| 2,4-Diaminotoluene | 2,4DAT | 95-80-7 | W | D |
| 1-Naphthylamine | 1NA | 134-32-7 | T | D |
| 2-Naphthylamine | 2NA | 91-59-8 | * | D |
| Phenacetin | 62-44-2 | W | D | |
| 1-Aminopyrene | 1AP | 1606-67-3 | W | D |
| Benzidine | 92-87-5 | * | D | |
| 3,3’-Dichlorobenzidine | DCB | 91-94-1 | S | D |
| o-Tolidine | 119-93-7 | T | D | |
| Heterocyclic amincs | ||||
| Trp-P-1 acetate | 68808-54-8 | W | S | |
| Trp-P-2 acetate | 62450-10-3 | W | S | |
| AaC | 26148-68-5 | W | D | |
| IQ | 76180-96-6 | W | D | |
| MeIQ | 77094-11-2 | W | D | |
| PhIP hydrochloride | 105650-23-5 | W | S | |
| Dialkyl N-nitrosoamines | ||||
| N-Nitrosodimethylamine | DMN | 62-75-9 | W | S |
| N-Nitrosoethylmethylamine | EMN | 10595-95-6 | W | S |
| N-Nitrosodiethyamine | DEN | 55-18-5 | W | S |
| N-Nitrosodiethanolamine | DEolN | 1116-54-7 | S | S |
| N-Nitrosodipropylamine | DPN | 621-64-7 | N | D |
| N-Nitrosodibutylamine | DBN | 924-16-3 | S | D |
| N-Nitrosomorpholine | NMOR | 59-89-2 | S | S |
| N-Nitrosopiperidine | NPIP | 100-75-4 | Ka | S |
| N-Nitrosopyrrolidine | NPYR | 930-55-2 | Ka | D |
| Azo dyes | ||||
| Azobenzene | 103-33-3 | T | D | |
| p-Aminoazobenzene | pAAB | 60-09-3 | W | D |
| p-Dimethylaminoazobenzene | DAB | 60-11-7 | W | D |
| Nitro compounds | ||||
| 1-Nitronaphtherene | 1NN | 86-57-7 | T | D |
| 1-Nitropyrene | 1NP | 5522-43-0 | T | D |
| Others | ||||
| Aflatoxin B1 | AFB | 1162-65-8 | W | S |
| Cyclophosphamide | CYP | 50-18-0 | W | S |
Table 1: Pro-mutagens tested in this study.
aK: Kanto Chemical Co., Inc., Tokyo (Japan); Ka: Katayama Chemical; S: Sigma Chemical Co., St. Louis, MO (U.S.A.). *2-Naphthylamine was synthethized from 2-naphththol and ammonium sulfite in Laboratory of Genotoxicity, National Institute of Technology, Hachinohe College. Benzidine was synthethized from hydrazobenzene in Laboratory of Genotoxicity, National Institute of Technology, Hachinohe College. bD, DMSO; S, Saline.
Comet Assay
Since the aim of the present study was to compare the ability to detect pro-mutagens of endogenous system of human-derived hepatoma HepG2 cells to that of WTK1 cells under liver S9, HepG2 cells were treated for 4 h like as WTK1 cells were treated for 4 h with S9’s [3]. At the end of a 4 h treatment period, cells were trypsinized, isolated by centrifugation for 5 min and re-suspended in 200 µL medium.
One hundred microliters of the suspensions were used to determine viability using the trypan blue method to obtain relative cell viability to control. The highest treatment concentration was 5000 µg/mL or the concentration at which relative cell viability was about 70 %, or gross precipitation was not observed in the treatment medium. One-hundred- microliter portions of the remaining suspensions were used for the comet assay. Seventy five µL of 1% agarose GP-42 (Nakalai Tesque, Inc., Kyoto, Japan) was quickly layered on a slide (Matsunami Glass Ind., Ltd., Osaka, Japan) and covered with another slide glass. One-hundred-microliter of treated cell suspensions were mixed with in 100 µL of 2% agarose- LGT (Nakalai Tesque, Inc.) and 75 µL of cell suspension (5 x 105 cells/75µL) was quickly layered at the same manner after removing the slide glass. Finally, 75 µL of 1% agarose GP-42 was quickly laid on again. The slide were lysed immediately in a solution (pH 10) of 2.5 M NaCl, 100 mM Na2EDTA, 10 mM Trizma, 1% sarkosyl, 10% DMSO, and 1% Triton X-100 at 4oC for 60 min [3].
The slides were placed on a horizontal gel electrophoresis platform, and covered with an alkaline solution made up of 300 mM NaOH and 1 mM Na2EDTA (pH>13). The slides were left in the solution for 20 min to allow the unwinding of the DNA and expression of alkali-labile sites. The power supply was set at 25 V and 250 mA. The DNA was electrophoresed for 20 min and the slides were rinsed with 400 mM Trizma (pH 7.5) to neutralize the excess alkali. Each slide was stained with 50 µL of 20 µg/mL ethidium bromide (Wako Pure Chemical Industries, Ltd.) and covered with a coverslip. Cells on one slide were photographed (black and white 400 ASA Fuji film) as seen with the aid of fluorescence microscope (Olympus at 200x magnification) equipped with an excitation filter of 515 - 560 nm and a barrier filter of 590 nm. The length of the comet tail was measured for 50 cells per treatment group. The relationship between chemical treatment and tail length was analyzed by ANOVA and the Dunnett test.
Results
Figures 1A-G shows the length of the comet tail for each pro-mutagen. Relative survival less than 70% was considered to show cytotoxicity. All the studied pro-mutagens, except
1AP, IQ, MeIQ, and DEolN, yielded significant increase in tail length. DEolN showed neither the increase in tail length nor cytotoxicity at 5000 µg/mL or lower. 1AP showed remarkable cytotoxicity at 200 µg/mL but did not increase tail length at the concentration range where remarkable cytotoxicity was not observed (100 µg/mL or lower). IQ and MeIQ showed cytotoxicity at 2500 µg/mL or higher, but they did not increase tail length at the concentration range where cytotoxicity was not observed (1250 µg/mL or lower).
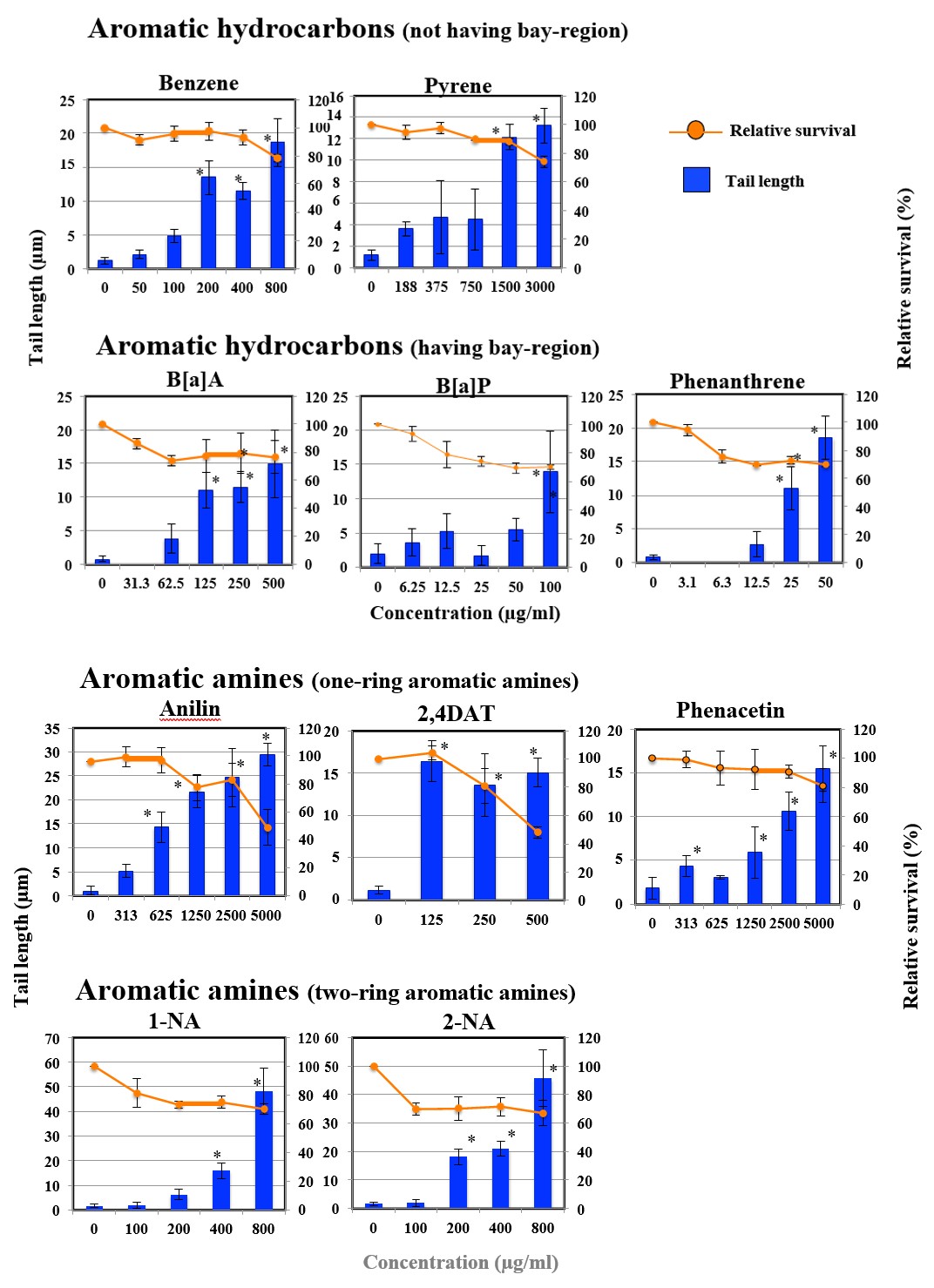
Figure 1A: The results of comet assay for aromatic hydrocarbons in HepG2 cells. Each graph shows Mean ± SD for 3 independent experiments. *Significant difference from untreated control: p<0.05.
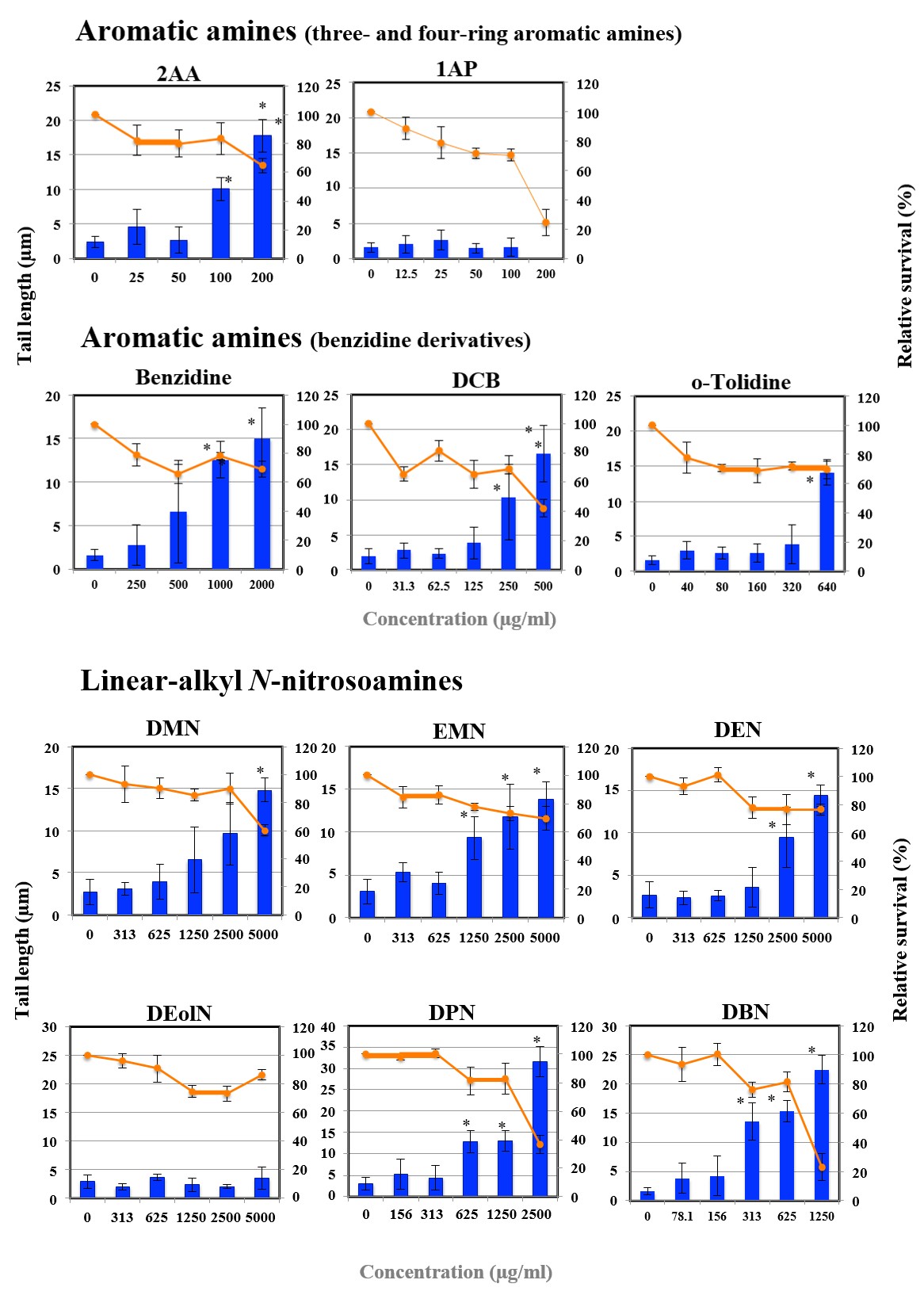
Figure 1B: The results of comet assay for aromatic amines in HepG2 cells. Each graph shows Mean±SD for 3 independent experiments. *Significant difference from untreated control: p<0.05.
Figure 1C: The results of comet assay for linear-alkyl N-nitrosoamines in HepG2 cells. Each graph shows Mean±SD for 3 independent experiments. *Significant difference from untreated control: p<0.05.
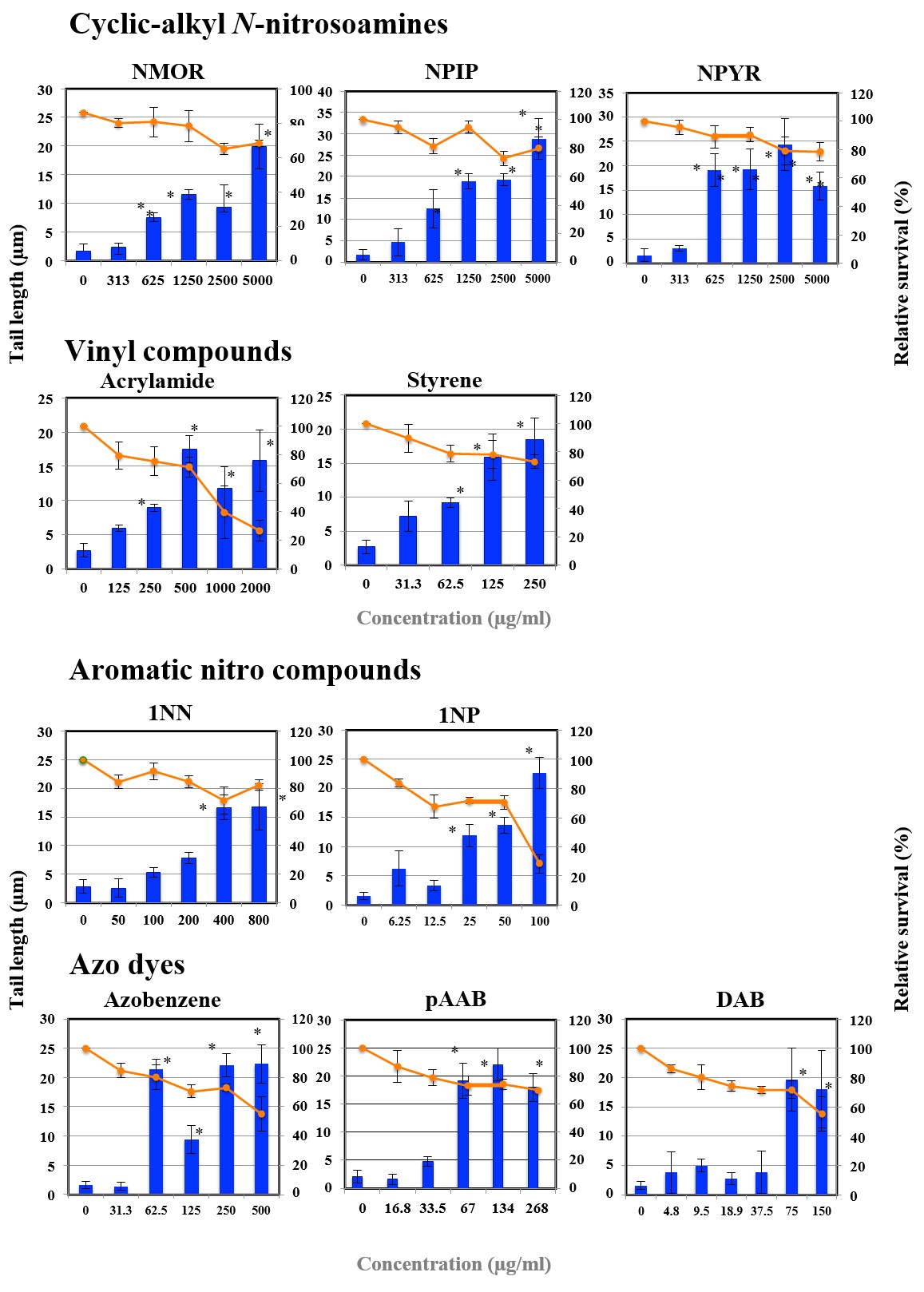
Figure 1D: The results of comet assay for cyclic-alkyl N-nitrosoamines and vinyl compounds in HepG2 cells. Each graph shows Mean±SD for 3 independent experiments. *Significant difference from untreated control: p<0.05.
Figure 1E: The results of comet assay for aromatic nitro compounds and azo dyes in HepG2 cells. Each graph shows Mean±SD for 3 independent experiments. *Significant difference from untreated control: p<0.05.
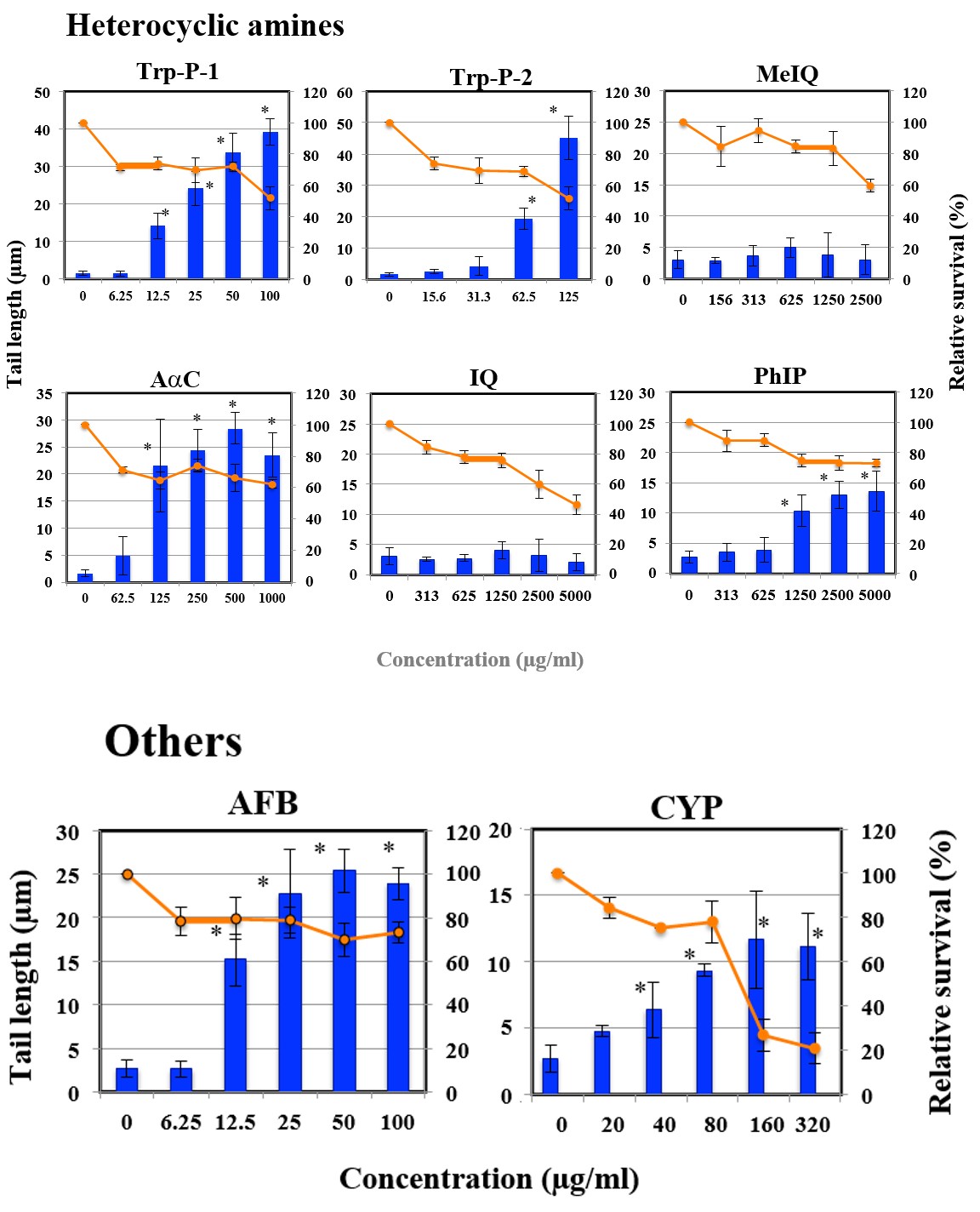
Figure 1F: The results of comet assay for heterocyclic amines in HepG2 cells. Each graph shows Mean±SD for 3 independent experiments. *Significant difference from untreated control: p<0.05.
Figure 1G: The results of comet assay for other compounds in HepG2 cells. Each graph shows Mean±SD for 3 independent experiments. *Significant difference from untreated control: p<0.05.
Discussion
It is shown that HepG2 cells retain the activities of certain phase I and phase II enzymes acting in the activation and detoxification of mutagens. Table2 shows drug metabolizing enzymes in HepG2 cells [5]. Table 3 shows enzymes primarily involved in metabolic activation of specific pro-mutagens. As shown in Tables 2 & 3, HepG2 cells have various drug metabolizing enzymes, it is reasonable that HepG2 cells can detect the genotoxicity of pro-mutagens which are preferentially activated by different P450 isozymes.
| Enzymes | Substrate metabolized | Remarks |
|---|---|---|
| Phase Ienzymes Total cytochrome P450 | 2.5 pmol/mg | |
| CYP1A1/1A2 | B[a]P and other Polycyclic hydrocarbons, DBN, Acrylamines, Aromatic amines, Heterocyclic amines | lower activity than human hepatocytes |
| CYP2B | 2AA, 2NA, Phenacetin | |
| CYP2C | Poly cyclic hydrocarbons | |
| CYP3A | 1NP | |
| CYP2E1 | Aniline, Benzene, Styrene, DMN, DEolN | |
| Arylhydrocarbon-hydroxylase | B[a]P, Aniline | |
| N-dimethylase | ||
| Catarase | ||
| Peroxidase | ||
| FMO | ||
| NAD(P)H: Cytchrome P450 reductase | Azo dyes, Nitro compounds | Similar activity as in human hepatocytes |
| NAD(P)H: Quinone oxidoreductase | Azo dyes, Nitro compounds, Quinone | twoisoenzymes expressed |
| Phase II enzymes Epoxide hydrolase | B[a]P-4,5-oxide | |
| Sulfo transferase ST | DMBA | very low sulfotransferase activity |
| GST | 2,4-Dinitrophenol | |
| UDGTP | 1-Naphthol | |
| N-acetyltransferase | p-Aminobenzoic acid, Anisidine |
Table 2: Drug metabolizing enzymes in HepG2 cells.
Drug metabolizing enzymes in HepG2 cells are cited from reference [5, 11] Table 2: Drug metabolizing enzymes in HepG2 cells.
| Enzymes | Pro-mutagens |
| CYP1A1 | B[a]P and other polycyclic hydrocarbons, DBN |
| CYP1A2 | Aromatic amines, Aromatic amides,1NPa |
| CYP2A6 | Phenacetin, DEN, DPNb, NMORb, NPIPb, NPYRb |
| CYP2C9 | Poly cyclichydro carbons |
| CYP2E1 | Benzene, Aniline, DMN, EMNa, DEolNc, Styrene, Acrylonitrile |
| CYP3A4 | Polycyclic hydrocarbon dihydrodiols, 6-Aminochrysene, 1NP, AFB1 |
| NADPH-P450reductase | Azo compounds, Nitro compounds |
Table 3: Enzymes primarily involved in metabolic activation of specific pro-mutagens.
References for enzymes primarily involved in metabolic activation:[11] a[12];b [13];c [14] Table 3: Enzymes primarily involved in metabolic activation of specific pro-mutagens.
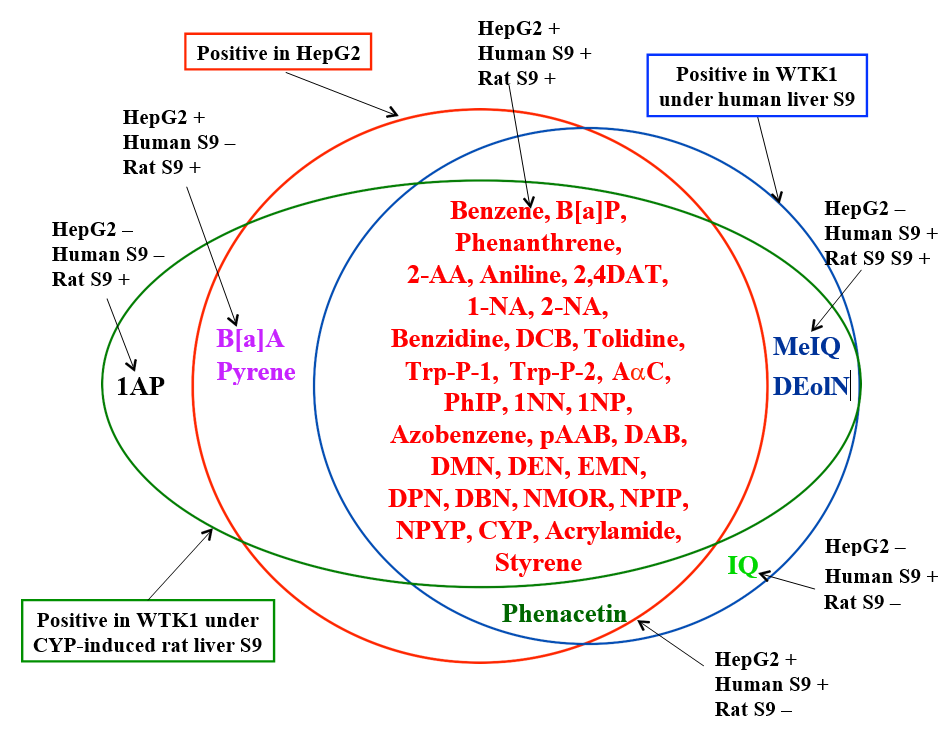
The genotoxicity of various mutagens, such as aflatoxins, heterocyclic aromatic amines, polycyclic aromatic hydrocarbons, and nitrosoamines, are detected in HepG2 cells without exogenous metabolic activation system [4, 5, 6, 7, 8]. Using HepG2 cells to detect pro-mutagens, it is important to know whether some pro-mutagens are missed by the use of HepG2 cells. We believe that the importance is whether detecting ability of HepG2 cells qualitatively parallels to that of metabolically incompetent cells under conventionally used CYP-induced rat liver S9.
To compare present results with HepG2 cells to previous results with metabolically incompetent WTK1 cells under human and rat S9’s, present results are summarized with previous results [3] and shown in Figure 2. Out of 38 pro- mutagens studied, 34 were positive in HepG2 cells.
Only 4 (1AP, IQ, MeIQ, and DEolN) were negative in HepG2 cells. 1AP was negative in both WTK1 cells under human liver S9 and HepG2 cells. 1AP and IQ were positive only in WTK1 cells under CYP-induced rat liver S9 and human liver S9, respectively. Phenacetin was positive in both HepG2 cells and WTK1 cells under human but not rat liver S9. There were 5 cases where the positive and negative results of WTK1cells under human S9 and HepG2 cells were inconsistent, i.e., MeIQ, IQ, and DEolN were negative in HepG2 cells but positive in WTK1 cells under human liver S9. B[a]A and pyrene were positive in HepG2 cells but negative in WTK1 cells under human liver S9. There were 4 cases where the positive and negative results of WTK1cells under rat liver S9 and HepG2 cells were inconsistent, i.e., 1AP, MeIQ, IQ, and DEolN were negative in HepG2 cells but positive in WTK1 cells under rat liver S9. Out of 38 pro-mutagens studied, the positive and negative results of 32 pro-mutagens were consistent between WTK1 cells under human liver S9 and HepG2 cells. The consistent rate between HepG2 cells and WTK1 cells under human liver S9 was 82.2%. Out of 38 promutagens studied, the positive and negative results of 33 pro-mutagens were consistent between WTK1 cells under rat liver S9 and HepG2 cells. The consistent rate between WTK1 cells under rat liver S9 and HepG2 cells was 86.8%.
These results show high concordance between the results by HepG2 cells and those by WTK1 cells under human and rat S9’s. In our previous study, it is shown that the exposure period to mutagens is an important factor to affect the outcomes of the comet assay and that long exposure period should not be employed for direct acting mutagens that are unstable in culture media [9]. HepG2 cells are assumed to have drug metabolizing enzymes that metabolize 4 pro-mutagens (IQ, MeIQ, DEolN, and 1AP) that were negative in this study: i.e., heterocyclic amines (IQ and MeIQ), DEolN, and 1AP could become substrates for CYP1A2, CYP2E1, and CYP1A2, respectively (Tables 2 & 3). Therefore, the genotoxicity of these 4 pro-mutagens could be detected by HepG2 cells, if optimal exposure period (exposure period in HepG2 cells means not only exposure period of mutagens to DNA but also metabolizing period) is selected. As discussed by Uhl, et al. [7], the long exposure period might be necessary to detect the genotoxicity of pro-mutagens by HepG2 cells.
The most frequently used methods to detect genotoxic effects of pro-mutagens in HepG2 cells are the alkaline elution technique, detection of the induction of sister chromatid exchanges, micronuclei, or 6TGr mutations [5]. Among them, the alkaline elution technique detects the induction of DNA single strand breaks and alkali labile sites like as the comet assay does. It was reported that 7 N-nitroso compounds (DMN, DEN, DPN, DBN, NMOR, NPIP, and NPYR) led to positive responses in the alkaline elution experiments [6]. In the comet assay, the positive results have been reported earlier with B[a]P, CYP, DMN, IQ, dimethylbenz[a]anthracene (DMBA), 2-acetylaminofluorene (2AAF), and hexamethylphosphoramide. With 150 µM B[a]P, 2 mM IQ, and 90 mM DMN, it was reported that tail lengths increased gradually as a function of the exposure periods, with the significant increased values reached after 20-24 h [7]. In this study, the treatment with IQ for 4 h did not induce DNA damage, which might coincide with their findings. Furthermore, optimal induction of DNA damage by 2AAF, B[a]P, CYP, and DMBA is observed after 72 h incubation [7]. Since the aim of the present study was to compare the ability of endogenous system in human-derived hepatoma HepG2 cells to that of WTK1 cells under liver S9, HepG2 cells were treated for 4 h like as WTK1 cells were treated for 4 h with S9’s [3]. In this study, 4 out of 38 pro-mutagens did not showed positive responses in HepG2 cells. Considering those earlier results, using extended incubation times at lower concentrations of these 4 pro-mutagens may allow HepG2 cells to be continuously exposed to metabolites within the cells without toxicity to the cells and to increase tail length.
The major advantage of the using metabolically competent human-derived cells is its low cost and directly measuring genotoxic effects in the activating cells [10]. In the present approach, we ascertained that the genotoxicity of pro-mutagens, which are traditionally detected by test procedures with metabolically incompetent cells requiring addition of exogenous activation mixtures, can be detected by the comet assay with HepG2 cells. We demonstrated in the present study that positive results are obtained in the comet assay using HepG2 cells for representatives of different classes of pro-mutagens and anticipate that this system might become a promising alternative to in vitro assays with metabolically incompetent indicator cells which are currently used in routine testing [11, 12, 13, 14].
Acknowledgements
This research was conducted in Advanced Material and Biological Engineering Course, National Institute of Technology, Hachinohe College as part of a graduation research project by Saeko Okutani under the research guidance of course staffs based on allocation of school educational expenses. The authors acknowledge the Advanced Material and Biological Engineering Course, National Institute of Technology, Hachinohe College.
Ethical Approval
It is not applicable.
References
-
Guengerich FP, Shimada T (1991) Oxidation of toxic and carcinogenic chemicals by human cytochrome P-450 enzymes. Chem Res Toxicol 4: 391-407.
-
Maron DM, Ames BN (1983) Revised methods for the Salmonella mutagenicity test. Mutation Res 113(3-4): 173-215.
-
Kawaguchi S, Nakamura T, Tsuda S, Murashige R, Sasaki YF (2018) Detection of in vitro genotoxicity of pro- mutagens using the comet assay under human and rat liver S9 fractions. MOJ Toxicol 4(4): 255-261.
-
Aden DP, Vogel A, Plotkin S, Damjanov I, Knowles BB (1979) Controlled synthesis of HBs Ag in a differentiated human liver carcinoma-derived cell line. Nature 282(5739): 615-616.
-
Knasmuller S, Parzefall W, Sanyal R, Ecker S, Schwab C, et al. (1998) Use of metabolically competent human hepatoma cells for the detection of mutagens and antimutagens. Mutation Res 402(1-2): 185-202.
-
Brambilla-Campart G, Canonero R, Mereto E, Ferro M (1989) Cytotoxic and genotoxic effects of 10 N-nitroso- compounds in a human hepatoma cell line HepG2: comparison with human hepatocyte Primary Cultures. ATLA 17: 22-27.
-
Uhl M, Helma C, Knasmuller S (2000) Evaluation of the single cell gel electrophoresis assay with human hepatoma (Hep G2) cells. Mutation Res 468(2): 213-225.
-
Doostdar H, Duthie SJ, Burke MD, Melvin WT, Grant MH (1988) The influence of culture medium composition on drug metabolizing enzyme activities of the human liver- derived Hep G2 cell line. FEBS Lett 241(1-2): 15-18.
-
Sasaki YF, Nakamura T, Kawaguchi S (2007) What is better experimental design for in vitro comet assay to detect chemical genotoxicity?. Proc. 6th World Congress on Alternatives & Animal Use in the Life Sciences: 499- 504.
-
Natarajan AT, Darroudi F (1991) Use of human hepatoma cells for in vitro metabolic activation of chemical mutagens/carcinogens. Mutagenesis 6(5): 399-403.
-
Parkinson A (1996) Biotransformation of xenobiotics. In: Klaassen CD, (Ed.), Casarett and Doull’s Toxicology: The Basic Science of Poisons. 5th (Edn.), New York: McGraw-Hill, PP: 113-186.
-
Wang M, Gao Y, Luo N, Chen G Niu X, et al. (2023) 1-Nitropyrene Metabolism and Health Risk: Identification of Key Enzymes, Pathways, and Metabolites Using a Multimethod Approach Environ. Health 1(6): 383-393.
-
Fujita K, Kamataki T (2001) Predicting the mutagenicity of tobacco-related N-nitrosamines in humans using 11 strains of Salmonella typhimurium YG7108, each coexpressing a form of human cytochrome P450 along with NADPH-cytochrome P450 reductase. Environ Mol Mutagen 38(4): 339-346.
-
Loeppky RN, Fuchs A, Janzowski C, Humberd C, Goelzer P, et al. (1998) Probing the mechanism of the carcinogenic activation of N-nitrosodiethanolamine with deuterium isotope effects: in vivo induction of DNA single-strand breaks and related in vitro assays. Chem Res Toxicol 11(12): 1556-1566.
- Evaluation of Skin Aging Preventive Effects of Cherry Blossom Petal Extracts Through Antioxidant and Anti-Glycation Activities
- Is Cell Death Responsible for False Positive Results of In Vivo Comet Assay?
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism