How Oncogenic Viruses Exploit p62-Mediated Selective Autophagy for Cancer Development
The 2016 Nobel Prize in Physiology or Medicine was awarded to Dr. Yoshinori Ohsumi for his discoveries of mechanisms for autophagy. Autophagy is a fundamental and conserved cellular program essential for maintaining cellular homeostasis. Unlike non-selective autophagy (including mitophagy) that sorts harmful or surplus cellular contents to the lysosomes for degradation and recycling, selective autophagy is mediated by an increasing pool of receptors, such as p62, NBR1, TAX1BP1, NDP52, OPTN, BCL2L13, FUNDC1, CCDC50, TRIMs, and TOLLIP, and is generally invoked by certain stresses to specifically target functional substrates for lysosomal degradation to modulate various cellular responses [1- 4].
Mini Review
The 2016 Nobel Prize in Physiology or Medicine was awarded to Dr. Yoshinori Ohsumi for his discoveries of mechanisms for autophagy. Autophagy is a fundamental and conserved cellular program essential for maintaining cellular homeostasis. Unlike non-selective autophagy (including mitophagy) that sorts harmful or surplus cellular contents to the lysosomes for degradation and recycling, selective autophagy is mediated by an increasing pool of receptors, such as p62, NBR1, TAX1BP1, NDP52, OPTN, BCL2L13, FUNDC1, CCDC50, TRIMs, and TOLLIP, and is generally invoked by certain stresses to specifically target functional substrates for lysosomal degradation to modulate various cellular responses [1, 2, 3, 4].
The role of autophagy in cancer is still under debate. However, it is now generally accepted that autophagy counteracts tumorigenesis at early stages, whereas promotes metabolism, survival, and progression of already established cancers [5, 6, 7]. As such, oncogenic viruses are known to induce non-selective autophagy and inhibit the final stage of autophagy at their early stage of infection for optimal replication and oncogenic transformation, but induce autophagy in their latency to facilitate their persistence, as well as tumor initiation, metastasis and development [8, 9, 10, 11].
Interestingly, a few recent reports highlight the emerging roles of p62-mediated selective autophagy in the latency maintenance and cancer development in the setting of oncogenic viral infections. We provide some general information in this short overview on how oncogenic viruses promote oncogenesis by exploiting this autophagy mechanism.
Oncogenic Viruses Activate P62-Mediated Selective Autophagy in Their Latency
The discovery of the cellular mechanism of oxygen sensing has awarded the 2019 Nobel Prize in Physiology and Medicine, highlighting its importance in cellular functions. Reactive oxygen/nitrogen species (ROS/RNS, referred to ROS hereafter) are produced by extracellular or intracellular stresses such as cancer chemotherapeutic drugs, and mitochondria produce the majority of ROS [12]. The mitochondria in malignant cells are functionally and structurally deregulated and are able to overproduce ROS [13]. Cancer hypoxia also induces intracellular ROS through the mechanisms involving the hypoxia-inducible factor (HIF) family of transcription factors [14]. Of note, ROS production can be directly induced by signaling pathways downstream of viral oncoproteins in the latency, including EBV LMP1 and EBNA1 [15, 16, 17], HTLV1 Tax [18], KSHV LANA [19, 20], and HPV E6 and E7 [21, 22].
ROS activate various transcription factors such as NFκB, AP1, HIF1α and STAT3 essential for cancer initiation and development, control the expression of various tumor suppressor genes such as p53, Rb, and pTEN [23], and also induce autophagy [24, 25]. ROS are a double-edged sword, depending on their levels; they either promote cell- autonomous apoptosis and anticancer immunosurveillance, or promote chronic inflammation that favors carcinogenesis under certain conditions [26]. ROS represent a crucial component of the tumor microenvironment, in which ROS incites inflammation, and excess inflammation in turn causes oxidative stress, ultimately perturbing genomic instability that promotes malignant transformation [27, 28].
Notably, ROS also trigger the activation of the antioxidative defense, with the Keap1-NRF2 pathway being the master antioxidative defense mechanism that plays a role in cancer (Figure 1) [29, 30, 31]. The transcription factor NRF2, a member of the basic leucine zipper (bZIP) family, is constitutively ubiquitinated for proteasomal degradation under normoxia by the Ub E3 ligase complex Keap1/ Cul3/RBX1. ROS trigger autophagic degradation of Keap1, resulting in the accumulation and activation of NRF2. As the master regulator of oxidative and inflammatory Stresses, NRF2 transactivates about 250 genes, including p62, Keap1, Cox-2, iNOS, PRDX1, HIF1, NQO1, HMOX1, GSTs, and NRF2 itself [32, 33]. Due to its importance in redox homeostasis, quick activation of NRF2 is a hallmark in response to ROS under various stresses [34, 35, 36].
The ubiquitin sensor and selective autophagy receptor p62 is one of the most important transcriptional targets of NRF2. In turn, phosphorylated p62 (S349) binds to Keap1 and disrupts Keap1-NRF2 interaction, promoting NRF2 stabilization and activation in a positive regulatory circuit [37, 38]. Induction of p62 expression is a prerequisite for consequent activation of p62- mediated selective autophagy, which also requires autophagosome biogenesis and site- specific PTMs of p62, including phosphorylation of S403 and ubiquitination of K7/K420 [39].
Importantly, recent studies have indicated that oncogenic viruses induce p62 expression in their latency, through different mechanisms, including the Keap1-NRF2 pathway [40, 41]. We have shown that EBV and HTLV1 activate p62- mediate selective autophagy in their latency [42]. To further explore the mechanisms, we have accumulated evidence showing that EBV and HTLV1 induce p62, partially by NFκB that is activated downstream of LMP1 and Tax signaling pathways, respectively, and also likely by the Keap1-NRF2 pathway (to be published).
p62-mediated Selective Autophagy Serves as an Alternative DNA Damage Response Mechanism
ROS are the major cause of endogenous DNA damage, including double-strand DNA breaks (DSBs), which is directly linked to many human diseases including cancer [43, 44, 45]. Eukaryotic organisms have developed complicated mechanisms to repair DNA damage to ensure genomic integrity, and thus the DNA repair machinery represents a highly complex defense mechanism [46]. Homologous recombination (HR) and non-homologous end-joining (NHEJ) are two major mechanisms responsible for the repair of DSBs [47].
ROS production and DNA damage are the most common features of cancers and chronic viral infections [48, 49]. Most cancers, if not all, harbor deficient traditional DNA repair mechanisms [50]. As such, viral infections can hijack the traditional DNA repair mechanisms. As a part of DNA damage response (DDR), p62-mediated selective autophagy serves as an alternative DNA repair mechanism in these settings, allowing these cancer or infected cells to escape DNA damage- induced cell death, by selectively targeting degradation of p62 itself and other DDR-related proteins such as RAD51 (Figure 1) [51, 52].
We have shown that a well-controlled autophagy-p62 interplay renders EBV-infected cells with the ability to balance pro-survival DNA damage resistance and pro-mutagenic genomic perturbation under oxidative stress [42, 53], and have defined two distinct roles for the autophagy-p62 interplay in this setting: cytoplasmic p62 mediates selective autophagy, and nuclear p62 upon autophagy inhibition represses DNA repair at least by promoting proteasome- mediated degradation of CHK1 and RAD51 [42]. These original findings disclose novel mechanisms underneath LMP1-mediated oxidative DDR. Elucidating how p62- mediated autophagy regulates DDR is necessary for improving our understanding of its mechanistic role in viral latency and oncogenesis.
p62-mediated Selective Autophagy Suppresses the Antitumor cGAS-STING Innate Immune Pathway
DNA fragments derived from DNA damage, if left unrepaired, can accumulate in the cytoplasm and serves as a major category of damage-associated molecular patterns (DAMPs), which are recognized by host germline-encoded pathogen recognition receptors (PRRs) [54, 55, 56, 57], leading to chronic inflammation that promotes immunosenescence, inflammaging, and the development of tumor microenvironment under certain conditions [27, 28, 58, 59].
Among these PRR pathways, the cGAS-STING pathway plays the most important role in sensing endogenous damaged DNA fragments (Figure 1) [60, 61, 62]. p62-mediated selective autophagy inhibits cGAS-STING signaling through different mechanisms. In turn, the cGAS-STING pathway promotes p62-mediated selective autophagy. Mechanistically, there are multiple points of crosstalk between these two processes, including that TBK1 targets both IRF3 (also STING) and p62 for activation [39], that STING can directly induce autophagy [63, 64, 65, 66], and that p62-mediated selective autophagy can selectively target cGAS and STING for autophagic degradation (Figure 1) [67].
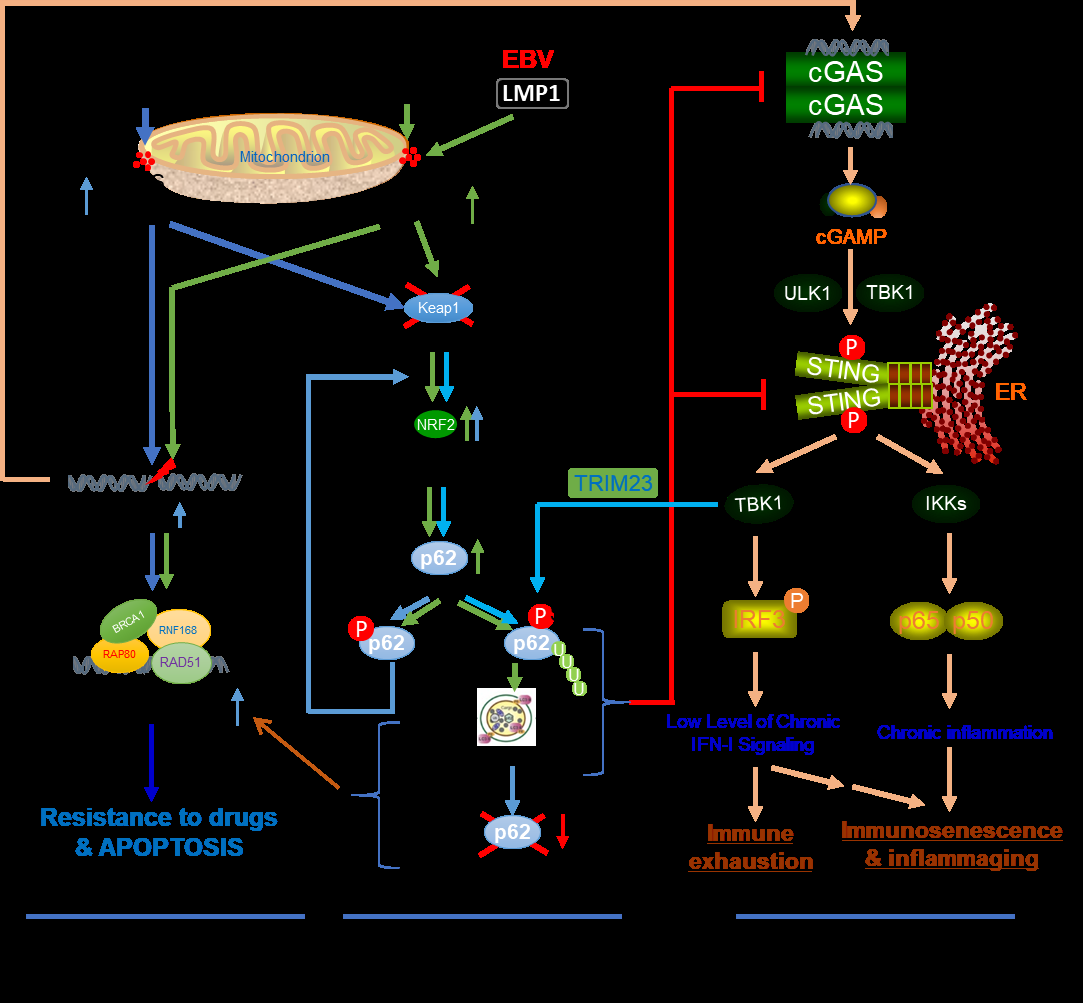
Figure 1: Autophagy mediates the crosstalk of oxidative stress with the Keap1-NRF2 antioxidative response, the DNA damage response, and the cGAS-STING-mediated antitumor immune response. ROS are produced by various stresses, such as viral infections, cancer chemotherapeutic drug treatments, and cancer hypoxia, as well as by signaling pathways triggered by oncogenic viral proteins. Different levels of ROS result in distinct outcomes, through the activation of: 1) DDR. p62-mediated selective autophagy is induced by ROS and promotes DNA repair in cancer cells, conferring to drug resistance. massive DNA damage that cannot be repaired causes cell death; 2) antioxidative response. The master antioxidant transcription factor NRF2 remains at low protein levels. ROS promote the accumulation and activation of NRF2 in an autophagy-dependent manner. Activated NRF2 induces the expression of a pool of target genes, including p62 that is required for the induction of p62-mediated selective autophagy; and 3) DNA sensing pathways, with the cGAS-STING pathway being the major one. p62-mediated selective autophagy suppresses the cGAS-STING- mediated signaling, and in turn, the cGAS-STING pathway promotes p62-mediated selective autophagy.
Perspectives on Cancer Therapy
Aberrant redox homeostasis, deficient DDR machinery, chronic inflammation, genomic instability, evasion of the immune response, and resistance to cell death are among the hallmarks of cancer [68, 69, 70]. Autophagy plays a crucial role in the crosstalk of these hallmarks, and frequently contributes to cancer chemoresistance [71, 72]. The indispensable requirement of autophagy for the survival of many cancer cells suggests targeting autophagy may serve as a viable therapeutic strategy [73, 74, 75, 76, 77, 78, 79]. All anticancer chemotherapeutic agents (such as etoposide and doxorubicin) produce ROS, which are able to induce autophagy as well as DNA damage to mount the cGAS-STING immune response that triggers cell death and tumor rejection in cancer cells [80, 81]. Inhibition of physiological autophagy (both selective and non-selective) can lead to cell death in cancers [24]. Lysosome inhibitors, such as hydroxychloroquine, inhibit autophagy and are clinically important for treating drug-resistant cancers [82].
However, inhibition of autophagy activity alone is not specific for cancer therapy although several autophagy inhibitors are in clinical trials. Combined targeting a cellular mechanism, especially the cGAS-STING pathway that is specifically coupled with autophagy, in a given cancer context may improve clinical efficacy and specificity.
Competing Interests
The authors declare that they have no competing interests.
Acknowledgment
This work was supported by NIH (CA252986 and DE027314) and ASH, and in part by the NIH grant C06RR0306551. This publication is the result of work supported with resources and the use of facilities at the James H. Quillen Veterans Affairs Medical Center. The contents in this publication do not represent the views of the Department of Veterans Affairs or the United States Government.
References
-
Farre JC, Subramani S (2016) Mechanistic insights into selective autophagy pathways: lessons from yeast. Nat Rev Mol Cell Biol 17(9): 537-552.
-
Zaffagnini G, Martens S (2016) Mechanisms of Selective Autophagy. Journal of Molecular Biology 428(9Part A): 1714-1724.
-
Sparrer KMJ, Gack MU (2018) TRIM proteins: New players in virus-induced autophagy. PLOS Pathogens 14(2).
-
Li W, He P, Huang Y, Li YF, Lu J, et al (2021) Selective autophagy of intracellular organelles: recent research advances. Theranostics 11(1): 222-256.
-
Lorin S, Hamai A, Mehrpour M, Codogno P (2013) Autophagy regulation and its role in cancer. Semin Cancer Biol 23(5): 361-379.
-
White E (2015) The role for autophagy in cancer. J Clin Invest 125(1): 42-46.
-
Santana-Codina N, Mancias JD, Kimmelman AC (2017) The Role of Autophagy in Cancer. Annual Review of Cancer Biology 1(1): 19-39.
-
Vanzo R, Bartkova J, Merchut-Maya JM, Hall A, Bouchal J, et al (2019) Autophagy role(s) in response to oncogenes and DNA replication stress. Cell Death Differ 27(3): 1134-1153.
-
Silva LM, Jung JU (2013) Modulation of the autophagy pathway by human tumor viruses. Semin Cancer Biol 23(5): 323-328.
-
Pratt ZL, Sugden B (2012) How human tumor viruses make use of autophagy. Cells 1(3): 617-630.
-
Mattoscio D, Medda A, Chiocca S (2018) Human Papilloma Virus and Autophagy. Int J Mol Sci 19(6): 1775.
-
Perillo B, Di Donato M, Pezone A, Di Zazzo E, Giovannelli P, et al. (2020) ROS in cancer therapy: the bright side of the moon. Exp Mol Med 52(2): 192-203.
-
Fulda S, Galluzzi L, Kroemer G (2010) Targeting mitochondria for cancer therapy. Nat Rev Drug Discov 9(6): 447-464.
-
Kawanishi S, Ohnishi S, Ma N, Hiraku Y, Murata M (2017) Crosstalk between DNA Damage and Inflammation in the Multiple Steps of Carcinogenesis. Int J Mol Sci 18(8): 1808.
-
Cerimele F, Battle T, Lynch R, Frank DA, Murad E, et al (2005) Reactive oxygen signaling and MAPK activation distinguish Epstein–Barr Virus (EBV)-positive versus EBV-negative Burkitt’s lymphoma. Proc Natl Acad Sci USA 102(1): 175-179.
-
Sun J, Hu C, Zhu Y, Sun R, Fang Y, et al. (2015) LMP1 Increases Expression of NADPH Oxidase (NOX) and Its Regulatory Subunit p22 in NP69 Nasopharyngeal Cells and Makes Them Sensitive to a Treatment by a NOX Inhibitor. PLoS One 10(8): e0134896.
-
Gruhne B, Sompallae R, Marescotti D, Kamranvar SA, Gastaldello S, et al. (2009) The Epstein-Barr virus nuclear antigen-1 promotes genomic instability via induction of reactive oxygen species. Proc Natl Acad Sci USA 106(7): 2313-2318.
-
Mohanty S, Harhaj EW (2020) Mechanisms of Oncogenesis by HTLV-1 Tax. Pathogens 9(7): 543.
-
Ye F, Zhou F, Bedolla RG, Jones T, Lei X, et al. (2011) Reactive Oxygen Species Hydrogen Peroxide Mediates Kaposi’s Sarcoma-Associated Herpesvirus Reactivation from Latency. PLoS Pathog 7(5): e1002054.
-
Li X, Feng J, Sun R (2011) Oxidative Stress Induces Reactivation of Kaposi’s Sarcoma- Associated Herpesvirus and Death of Primary Effusion Lymphoma Cells. J Virol 85(2): 715-724.
-
Marullo R, Werner E, Zhang H, Chen GZ, Shin DM, et al. (2015) HPV16 E6 and E7 proteins induce a chronic oxidative stress response via NOX2 that causes genomic instability and increased susceptibility to DNA damage in head and neck cancer cells. Carcinogenesis 36(11): 1397-1406.
-
Williams VM, Filippova M, Filippov V, Payne KJ, Duerksen- Hughes P (2014) Human papillomavirus type 16 E6* induces oxidative stress and DNA damage. J Virol 88(12): 6751-6761.
-
Gupta SC, Hevia D, Patchva S, Park B, Koh W, et al. (2012) Upsides and downsides of reactive oxygen species for cancer: the roles of reactive oxygen species in tumorigenesis, prevention, and therapy. Antioxid Redox Signal 16(11): 1295-1322.
-
Poillet-Perez L, Despouy G, Delage-Mourroux R, Boyer- Guittaut M (2015) Interplay between ROS and autophagy in cancer cells, from tumor initiation to cancer therapy. Redox Biol 4: 184-192.
-
Filomeni G, De Zio D, Cecconi F (2015) Oxidative stress and autophagy: the clash between damage and metabolic needs. Cell Death Differ 22(3): 377-388.
-
Zitvogel L, Kepp O, Galluzzi L, Kroemer G (2012) Inflammasomes in carcinogenesis and anticancer immune responses. Nat Immunol 13(4): 343-351.
-
Liao Z, Chua D, Tan NS (2019) Reactive oxygen species: a volatile driver of field cancerization and metastasis. Molecular Cancer 18(1): 65.
-
Weinberg F, Ramnath N, Nagrath D (2019) Reactive Oxygen Species in the Tumor Microenvironment: An Overview. Cancers 11(8): 1191.
-
Jaramillo MC, Zhang DD (2013) The emerging role of the Nrf2–Keap1 signaling pathway in cancer. Genes Dev 27(20): 2179-2191.
-
Mitsuishi Y, Motohashi H, Yamamoto M (2012) The Keap1–Nrf2 system in cancers: stress response and anabolic metabolism. Front Oncol 2: 200.
-
Kansanen E, Kuosmanen SM, Leinonen H, Levonen AL (2013) The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biology 1(1): 45-49.
-
Tonelli C, Chio IIC, Tuveson DA (2018) Transcriptional Regulation by Nrf2. Antioxid Redox Signal 29(17): 1727- 1745.
-
Ahmed SMU, Luo L, Namani A, Wang XJ, Tang X (2017) Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim Biophys Acta Mol Basis Dis 1863(2): 585-597.
-
Nguyen T, Nioi P, Pickett CB (2009) The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J Biol Chem 284(20): 13291-13295.
-
Lee C (2018) Therapeutic Modulation of Virus-Induced Oxidative Stress via the Nrf2- Dependent Antioxidative Pathway. Oxid Med Cell Longev.
-
Ramezani A, Nahad MP, Faghihloo E (2018) The role of Nrf2 transcription factor in viral infection. J Cell Biochem 119(8): 6366-6382.
-
Komatsu M, Ichimura Y (2010) Physiological significance of selective degradation of p62 by autophagy. FEBS Lett 584(7): 1374-1378.
-
Ichimura Y, Komatsu M (2018) Activation of p62/ SQSTM1-Keap1-NRF2- Related Factor 2 Pathway in Cancer. Frontiers in Oncology.
-
Ning S, Wang L (2019) The Multifunctional Protein p62 and Its Mechanistic Roles in Cancers. Curr Cancer Drug Targets 19(6): 468-478.
-
Schwob A, Teruel E, Dubuisson L, Lormières F, Verlhac P, et al. (2019) SQSTM-1/p62 potentiates HTLV-1 Tax- mediated NF-κB activation through its ubiquitin binding function. Scientific Reports 9(1): 16014.
-
Hong S, Li Y, Kaminski PJ, Andrade J, Laimins LA (2020) Pathogenesis of Human Papillomaviruses Requires the ATR/p62 Autophagy-Related Pathway. mBio 11(4): e01628-01620.
-
Wang L, Howell MEA, Sparks-Wallace A, Hawkins C, Nicksic C, et al. (2019) p62-mediated Selective Autophagy Endows Virus-transformed Cells with Insusceptibility to DNA Damage under Oxidative Stress. PLoS Pathogens 15(4): e1007541.
-
Srinivas US, Tan BWQ, Vellayappan BA, Jeyasekharan AD (2019) ROS and the DNA damage response in cancer. Redox Biol 25: 101084.
-
Tubbs A, Nussenzweig A (2017) Endogenous DNA Damage as a Source of Genomic Instability in Cancer. Cell 168(4): 644-656.
-
Vilenchik MM, Knudson AG (2003) Endogenous DNA double-strand breaks: Production, fidelity of repair, and induction of cancer. Proceedings of the National Academy of Sciences 100(22): 12871-12876.
-
Bouwman P, Jonkers J (2012) The effects of deregulated DNA damage signalling on cancer chemotherapy response and resistance. Nat Rev Cancer 12(9): 587-598.
-
Chang HHY, Pannunzio NR, Adachi N, Lieber MR (2017) Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nat Rev Mol Cell Biol 18(8): 495-506.
-
Kgatle MM, Spearman CW, Kalla AA, Hairwadzi HN (2017) DNA Oncogenic Virus-Induced Oxidative Stress, Genomic Damage, and Aberrant Epigenetic Alterations. Oxid Med Cell Longev.
-
Ivanov AV, Bartosch B, Isaguliants MG (2017) Oxidative Stress in Infection and Consequent Disease. Oxidative Medicine and Cellular Longevity.
-
O’Connor Mark J (2015) Targeting the DNA Damage Response in Cancer. Molecular Cell 60(4): 547-560.
-
Gomes LR, Menck CFM, Leandro GS (2017) Autophagy Roles in the Modulation of DNA Repair Pathways. Int J Mol Sci 18(11): 2351.
-
Hewitt G, Carroll B, Sarallah R, Correia-Melo C, Ogrodnik M, et al. (2016) SQSTM1/p62 mediates crosstalk between autophagy and the UPS in DNA repair. Autophagy 12(10): 1917-1930.
-
Wang L, Riggs K, Kohne C, Yohanon JU, Foxler DE, et al. (2018) LIMD1 Is Induced by and Required for LMP1 Signaling, and Protects EBV- Transformed Cells from DNA Damage-Induced Cell Death. Oncotarget 9(5): 6282-6297.
-
Hernandez C, Huebener P, Schwabe RF (2016) Damage- associated molecular patterns in cancer: a double-edged sword. Oncogene 35(46): 5931-5941.
-
Wang L, Ning S (2017) “Toll-free” pathways for production of type I interferons. AIMS Allergy Immunol 1(3): 143-163.
-
Patidar A, Selvaraj S, Sarode A, Chauhan P, Chattopadhyay D, et al. (2018) DAMP-TLR- cytokine axis dictates the fate of tumor. Cytokine 104: 114-123.
-
De Galarreta MR, Lujambio A (2017) DNA sensing in senescence. Nat Cell Biol 19(9): 1008-1009.
-
Finger EC, Giaccia AJ (2010) Hypoxia, inflammation, and the tumor microenvironment in metastatic disease. Cancer Metastasis Rev 29(2): 285-293.
-
Fulop T, Larbi A, Dupuis G, Le Page A, Frost EH, et al. (2018) Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: Friends or Foes?. Front Immunol 8: 1960.
-
Li T, Chen ZJ (2018) The cGAS-cGAMP-STING pathway connects DNA damage to inflammation, senescence, and cancer. J Exp Med 215(5): 1287-1299.
-
Ng KW, Marshall EA, Bell JC, Lam WL (2018) cGAS–STING and Cancer: Dichotomous Roles in Tumor Immunity and Development. Trends Immunol 39(1): 44-54.
-
Wan D, Jiang W, Hao J (2020) Research Advances in how the cGAS-STING Pathway Controls the Cellular Inflammatory Response. Frontiers in Immunology 11: 615.
-
Moretti J, Roy S, Bozec D, Martinez J, Chapman JR, et al. (2017) STING Senses Microbial Viability to Orchestrate Stress- Mediated Autophagy of the Endoplasmic Reticulum. Cell 171(4): 809-823.
-
Gui X, Yang H, Li T, Tan X, Shi P, et al. (2019) Autophagy induction via STING trafficking is a primordial function of the cGAS pathway. Nature 567(7747): 262-266.
-
Yang J, Tang X, Nandakumar KS, Cheng K (2019) Autophagy induced by STING, an unnoticed and primordial function of cGAS. Cell Mol Immunol 16(8): 683-684.
-
Liu D, Wu H, Wang C, Li Y, Tian H, et al. (2019) STING directly activates autophagy to tune the innate immune response. Cell Death & Differentiation 26(9): 1735-1749.
-
Prabakaran T, Bodda C, Krapp C, Zhang BC, Christensen MH, et al. (2018) Attenuation of cGAS‐STING signaling is mediated by a p62/SQSTM1‐dependent autophagy pathway activated by TBK1. EMBO J 37(8): e97858.
-
Hanahan D, Weinberg Robert A (2011) Hallmarks of Cancer: The Next Generation. Cell 144(5): 646-674.
-
Panieri E, Santoro MM (2016) ROS homeostasis and metabolism: a dangerous liason in cancer cells. Cell Death Dis 7(6): e2253.
-
Mantovani A (2009) Cancer: Inflaming metastasis. Nature 457(7225): 36-37.
-
Holohan C, Van Schaeybroeck S, Longley DB, Johnston PG (2013) Cancer drug resistance: an evolving paradigm. Nature Reviews Cancer 13: 714-716.
-
Housman G, Byler S, Heerboth S, Lapinska K, Longacre M, et al. (2014) Drug Resistance in Cancer: An Overview. Cancers 6(3): 1769-1792.
-
Fulda S (2017) Autophagy in Cancer Therapy. Front Oncol 7: 128.
-
Islam MA, Sooro MA, Zhang P (2018) Autophagic Regulation of p62 is Critical for Cancer Therapy. Int J Molecular Sci 19(5): 1405.
-
Marinkovic M, Sprung M, BuljubasicM, Novak I (2018) Autophagy Modulation in Cancer: Current Knowledge on Action and Therapy. Oxidative Medicine and Cellular Longevity.
-
Nigussie A, Mishore MK, Degaga Abdi A (2018) Autophagy as a Possible Target for Cancer Therapy. J Orthop Oncol 4: 1000124.
-
Sui X, Chen R, Wang Z, Huang Z, Kong N, et al. (2013) Autophagy and chemotherapy resistance: a promising therapeutic target for cancer treatment. Cell Death Dis 4(10): e838.
-
Zheng K, He Z, Kitazato K, Wang Y (2019) Selective Autophagy Regulates Cell Cycle in Cancer Therapy. Theranostics 9(1): 104-125.
-
Liu H, He Z, Simon HU (2016) Chapter 14-The Role of Autophagy in Cancer and Chemotherapy. Autophagy: Cancer, Other Pathologies, Inflammation, Immunity, Infection, and Aging. Academic Press 8: 253-265.
-
Luthra P, Aguirre S, Yen BC, Pietzsch CA, Sanchez- Aparicio MT, et al. (2017) Topoisomerase II Inhibitors Induce DNA Damage-Dependent Interferon Responses Circumventing Ebola Virus Immune Evasion. mBio 8(2): e00368.
-
Härtlova A, Erttmann SF, Raffi FAM, Schmalz AM, Resch U, et al. (2015) DNA Damage Primes the Type I Interferon System via the Cytosolic DNA Sensor STING to Promote Anti-Microbial Innate Immunity. Immunity 42(2): 332- 343.
-
Galluzzi L, Bravo-San Pedro JM, Levine B, Green DR, Kroemer G (2017) Pharmacological modulation of autophagy: therapeutic potential and persisting obstacles. Nature Reviews Drug Discovery 16: 487.
- Are the Vaccines the Only Solution to Prevent the COVID-19 Pandemic? Part Two
- Clinical Characteristics of Women in this New Global Immunodeficiency
- Cell Dynamics in HIV Pathogenesis: Insights and Implications
- Determination of the CDR (CDR1, CDR2) « Complementary- Determining Region Invertebrate Primitive Antibody from Sea Star »
- Prioritizing Care for High-Risk COVID-19 Patients in the EU: 10 Civic Recommendations to the Institutions
- Comprehensive Insights into ModRNA Vaccines: Persistent PP-Spike Recombinant Protein, Hyperimmune/Inflammatory Reactions, Thrombotic Vasculopathy, Chronic Organ Complications and Excess Deaths