Effects of Oxidative Stress Prevention Using DNA Aptamer (Aptamin C®) in Keratinocyte
Oxidative stress is one of the leading causes of various diseases. Antioxidants are effective against reactive oxygen species (ROS), a major factor in oxidative stress. Antioxidants play important roles in our body by scavenging free radicals, thereby preventing them from damaging biological molecules. Vitamin C is essential for various physiological functions such as the synthesis of collagen and neurotransmitters, and it is a major antioxidant. Although vitamin C is widely used in cosmetic and therapeutic products, it easily undergoes oxidation by air, pH, temperature, and UV light, thereby decreasing its potency as an antioxidant and reducing the shelf-life of products containing vitamin C. To overcome this drawback, we developed Aptamin C®, an innovative single-strand DNA aptamer that maximizes the antioxidant efficacy of vitamin C by binding to its reduced form and delaying its oxidation. To investigate the effect of Aptamin C® and vitamin C complex on human skin, we performed both in vitro and clinical tests. We observed that the Aptamin C® and vitamin C complex had effective ROS-scavenging and anti-inflammatory effects.
Sooho Choi*, Yoonjeong Hwang, Taejun Kim and Jeong Hoon Kim
Nexmos Inc., Republic of Korea *Corresponding author: Sooho Choi, Nexmos, Inc., 2207 U-Tower, 767, Sinsu-ro, Suji-gu, Yongin-si, Gyeonggi-do, Republic of Korea, Tel: 82-70-5138-0962; Fax: 82-70-8260-0300; Email: joon@nexmos.com
Introduction
A variety of biochemical reactions such as photosynthesis and respiration in organelles generate reactive oxygen species (ROS) [1, 2, 3, 4]. Oxidative stress conditions occur when ROS levels exceed cell defense mechanisms [5]. Oxidative stress directly impacts various diseases, especially skin diseases [6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16]. Besides, oxidative stress and inflammation are closely related. Several studies have shown that inflammation around the upper third of hair follicles suggests that inflammation has a pathogenic role in androgenic alopecia [17, 18, 19, 20, 21].
Biological antioxidants are present in low concentrations compared with an oxidizable substrate and substantially delay or prevent substrate oxidation [22]. The human body contains various antioxidants that prevent oxidative stress. Antioxidants such as vitamin C, vitamin E, coenzyme Q10, and polyphenolic compounds protect against oxidative stress- related diseases [23, 24, 25]. Vitamin C, also known as ascorbic acid (AA), is one of the many antioxidants that plays an important role in maintaining skin health. Normal skin cells contain higher concentrations of vitamin C than the blood. Vitamin C has roughly four roles in the skin. First, vitamin C is an extremely potent antioxidant that effectively counters skin damage-generated ROS. Second, vitamin C stimulates collagen synthesis by skin fibroblasts and stabilizes it. Third, vitamin C promotes keratinocyte differentiation, enhancing the barrier function. Fourth, vitamin C decreases melanin synthesis, which prevents hyperpigmentation of the skin [26]. However, vitamin C quickly loses its function due to its antioxidant properties.
To slow down the oxidation of vitamin C, we developed Aptamin C®, a DNA aptamer that binds to vitamin C and inhibits its oxidation, through SELEX [27]. Here, we report that Aptamin C® increases cell proliferation and confirm its positive effect on inflammation in vitro.
Materials and methods
Chemicals
2′,7′-dichlorofluorescin diacetate (DCFH-DA), AA, lipopolysaccharide (LPS), dexamethasone, and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich. Hydrogen peroxide (H2O2) was purchased from Wako. Aptamin C® was synthesized at IDT. Dulbecco’s modified eagle medium (DMEM), fetal bovine serum (FBS), penicillin– streptomycin, and distilled water were purchased from Gibco. 20X PBS was purchased from biosesang and diluted in gibco distilled water to obtain 1X PBS.
The nitric oxide (NO) detection kit was purchased from Intron. The Tumor Necrosis Factor alpha (TNF-α) ELISA kit was purchased from R&D system.
HaCaT and RAW264.7 Cell Culture
HaCaT and RAW264.7 cells were grown in a growth medium (DMEM supplemented with 5% FBS and 1% penicillin–streptomycin) and maintained at 37°C and 5% CO2. Cells between passages 1 and 10 were used in this study.
Cellular Antioxidant Assay (CAA) for ROS Production Detection
Human epithelial cells, HaCaT, were seeded at a density of 2.5 × 104 /well in a 96-well microplate with 100 µL of growth medium/well. We used only the inner wells of the plates because the results from the outer wells showed greater variation. 24 hours after seeding, the growth medium was removed and the wells were washed with PBS. All wells were treated with 100 µL of 10 µM DCFH-DA dissolved in PBS, wrapped in foil, and incubated for 1 h at 37°C under CO2 atmosphere. 10 mM DCFH-DA stock was dissolved in DMSO. Then, all wells were washed with PBS and treated three times with 50 µL of 100 µM H2O2 plus 50 µL of AA with or without Aptamin C® for 1 h at 37°C under CO2 atmosphere. After incubation, the 96-well microplate was read with a Synergy H1 Hybrid Multi-Mode Reader (BioTek Instruments, Inc.). Wavelengths: excitation 485 nm, emission 535 nm. The plate included triplicate control and blank wells: blank wells contained cells treated with DCFH-DA dye and 10% FBS without H2O2; control wells contained cells treated with DCFH-DA dye and 10% FBS with H2O2.
Oxidative conditions: 10 µM AA was mixed with Aptamin C® (100:1, mass ratio) at room temperature for 10 minutes, then 30 µM H2O2 was added and the reaction mixture was placed at 37°C for 1 h.
Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements (2007)
High-Performance Liquid Chromatography (HPLC) Analysis
To quantify AA by HPLC, we followed the general test method of the Korean Pharmacopoeia. The AA solution was filtered with a 0.2 mm PVDF filter and 10 µL was injected in a reverse phase C-18 HPLC column (Agilent, USA) (4. 6 × 250 mm, 5 μm diameter). The mobile phase consisted of sodium- 1-hexanesulfonate, glacial acetic acid, and methanol with a flow rate of 1 ml/min. The sample was detected at 254 nm on an Agilent 1260 Infinity II multiple wavelength detector. The retention time for standard AA was recorded as 3.41 min.
Oxidative conditions: 10 µM AA was mixed with Aptamin C® (100:1, mass ratio) at room temperature for 10 minutes, then 30 µM H2O2 was added and the reaction mixture was placed at 37°C for 1 h.
Antioxidant phytochemicals in cabbage (Brassica oleracea L. var. capitata) (2006).
Statistical analyses
All results are presented as mean ± SD. Pairs of means were compared using unpaired Student’s t-test. Multiple means were compared by ANOVA followed by Fisher’s least significant difference test. Differences were considered significant when p < 0.05 and p < 0.001.
Measurement of NO Production and TNF-α Production
The RAW264.7 cells were seeded into 24-well plates (1.6 × 105 cells/well) and incubated for 16 h. The cells were then treated with various concentrations of Aptamin C® and vitamin C in the presence or absence of 1 μg/mL LPS. After 24 h, 100 μL of the supernatants and 100 μL of the NO detection reagent were added to a 96-well plate and reacted for 10 min in the absence of light. The absorbance at 540 nm was measured with a microplate reader. The value was corrected by total protein value. Dexamethasone (7.84 μg/ mL) was used as a positive control.
Results
In vitro ROS inhibition ability using the CAA assay
The CAA assay enables quantifying ROS via the production of DCF. DCFDA penetrates the cell membrane and cellular esterases convert it to non-fluorescent DCF. ROS then oxidizes non-fluorescent DCF, which becomes fluorescent
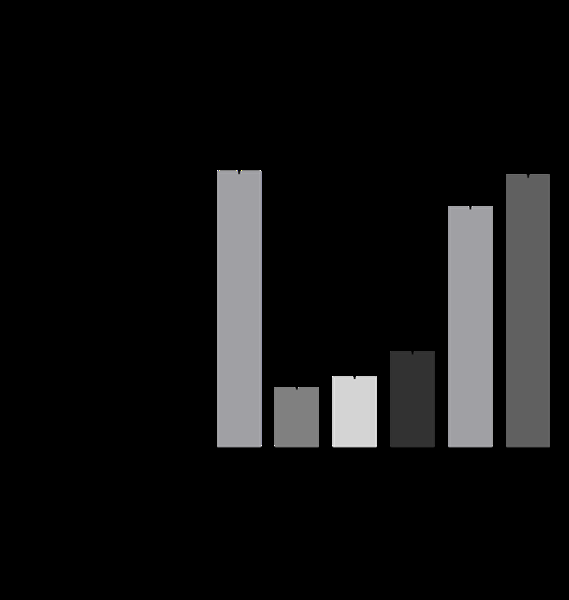
DCF.
To determine whether AA inhibits ROS production, we performed the CAA assay on HaCaT cells. Under oxidative conditions, the control cells generated ROS (Figure 1A). AA inhibited ROS production in a dose-dependent manner. Moreover, we confirmed that low concentrations of the oxidized form of AA, dehydroascorbic acid, did not inhibit ROS production (Figure 1B).
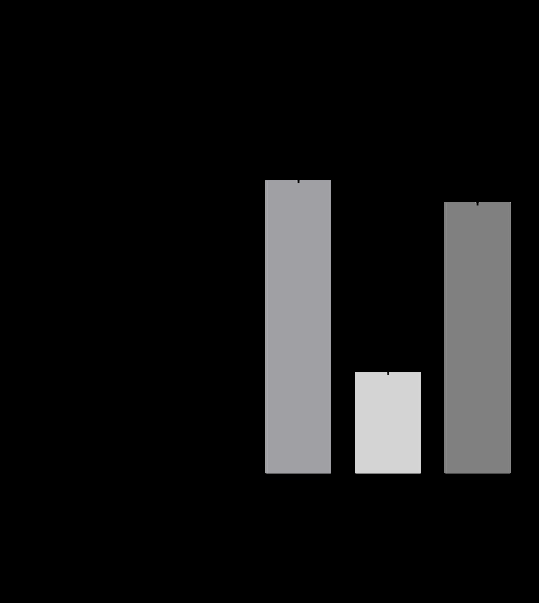
Antioxidant Effect of AA with/without Aptamin C® under Oxidative Conditions
To evaluate the stabilization effect of AA by Aptamin C®, we treated AA with an oxidant (H2O2) with or without Aptamin C® and quantified it by HPLC. Aptamin C® did not change the retention time of AA (Figure 2A). With Aptamin C®, the stabilization effect of AA was about 36% higher than
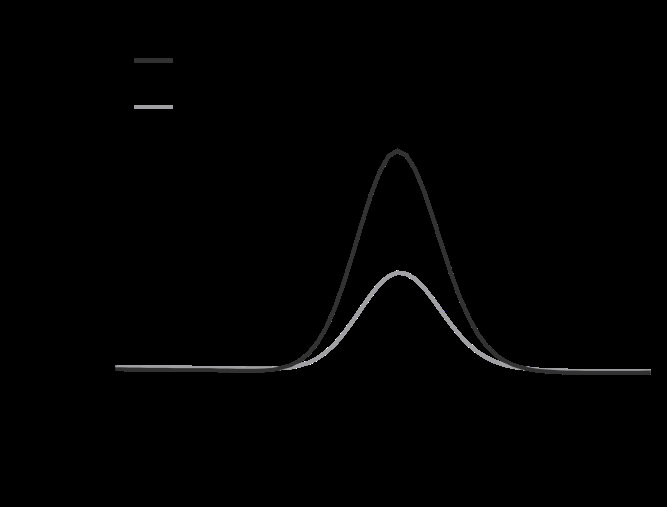
Figure 2: Stability of ascorbic acid with and without Aptamin C® under oxidative conditions. (A) HPLC chromatograms of ascorbic acid with or without Aptamin C®. (B) Ascorbic acid amount with or without Aptamin C®, as determined by HPLC. (C) ROS production by HaCaT cells in the presence of ascorbic acid, with or without Aptamin C®, using the CAA assay. The values are presented as mean ± SD (n = 3). ***, P-value < 0.001 (compared to other groups and as indicated).
without Aptamin C® (Figure 2B).
We also conducted CAA assay in HaCaT cell line to evaluate the stabilization effect of AA in cells with or without Aptamin C®. Under non-oxidative conditions, Aptamin C® did not affect the ROS inhibitory ability of AA. Whereas, under oxidative conditions, Aptamin C® enhanced the inhibition of ROS production (Figure 2C).
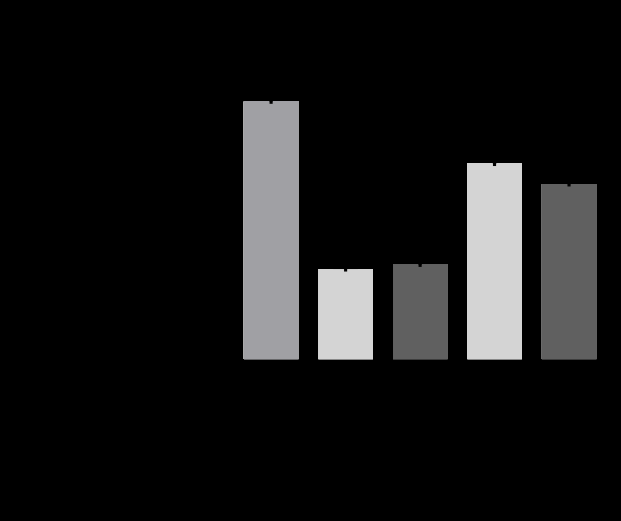
Effect on the Inflammatory Response Identified through Analysis of NO Production and TNF-α Production
NO production was significantly reduced in a dose- dependent manner, with 4.37%, 11.12%, 15.05%, 23.47%, and 24.19% at concentrations of 0.01 μg/mL + 0.5 μg/mL, 0.1 μg/mL + 5 μg/mL, 0.25 μg/mL + 12.5 μg/mL, 0.5 μg/mL +
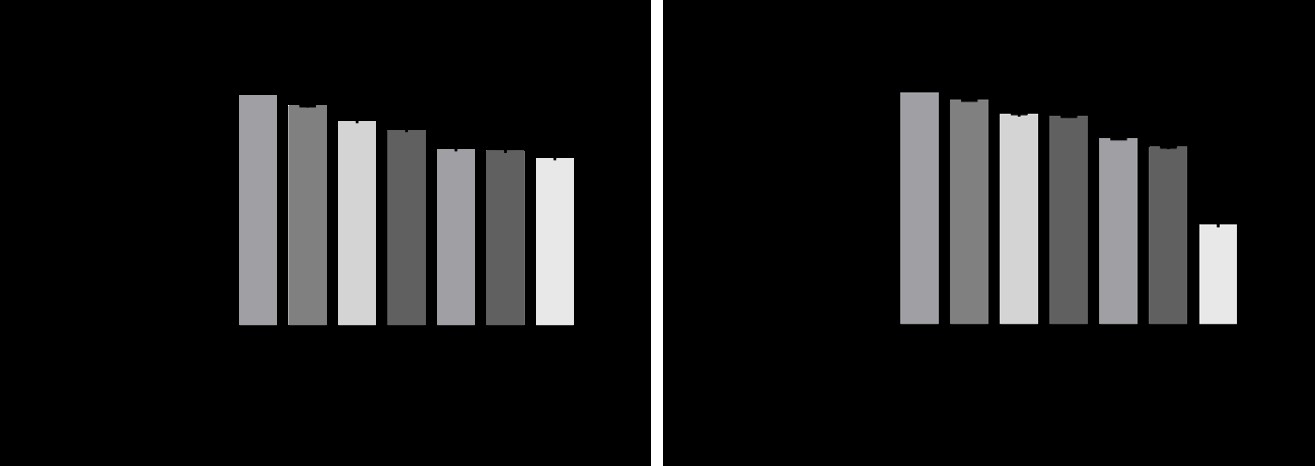
Figure 3: Anti-inflammatory effect of ascorbic acid with and without Aptamin C®. (A) NO production in RAW 264.7 cells after treatment. (B) TNF-α production in RAW 264.7 cells after treatment. The data are expressed as % of control. The values are presented as mean ± SD (n = 3). † P-value < 0.05 (Ascorbic acid with Aptamin C® vs control without LPS). * P-value < 0.05 (Ascorbic acid with Aptamin C® vs control with LPS).
25 μg/mL, and 1 μg/mL + 50 μg/mL (p < 0.05, Table 1, Figure 3A).
TNF-α production was significantly reduced in a dose- dependent manner, with 2.99%, 9.41%, 10.04%, 19.79%, and 23.54% at concentrations of 0.01 μg/mL + 0.5 μg/mL, 0.1 μg/mL + 5 μg/mL, 0.25 μg/mL + 12.5 μg/mL, 0.5 μg/mL + 25 μg/mL, and 1 μg/mL + 50 μg/mL (p < 0.05, Table 2, Figure 3B).
- Material
- LPS
- Dose (µg/mL)
- Mean
- (%)
- SD p-value2
- Change from solvent control (%)
- Aptamin C®
- Ascorbic acid
- -
- 0
- 0
- 3.64
- 0
- -
- -
- 0
- 0
- 100
- 0
- 0.000†
- -
- 0.01
- 0.5
- 95.63
- 1
- 0.000*
- 4.37▼
- Ascorbic acid with Aptamin
- C®
- 0.1
- 5
- 88.88
- 4
- 0.008*
- 11.12▼
- +
- 0.25
- 12.5
- 84.95
- 5
- 0.005*
- 15.05▼
- 0.5
- 25
- 76.53
- 2
- 0.000*
- 23.47▼
- 1
- 50
- 75.81
- 2
- 0.000*
- 24.19▼
- Dexamethasone1
- 7.84
- 72.68
- 4
- 0.000*
- 27.32▼
- 1 Positive control
- 2 Significant difference at p<0.05 compared to solvent control without LPS(†) and with LPS(*)
- Material
- LPS
- Dose (µg/mL)
- Mean
- (%)
- SD p-value2
- Change from solvent control (%)
- Aptamin C®
- Ascorbic acid
- -
- 0
- 0
- -
- 0
- -
- -
- 0
- 0
- 100
- 0
- 0.000†
- -
- 0.01
- 0.5
- 97.01
- 1
- 0.001*
- 2.99▼
- Ascorbic acid with Aptamin C®
- 0.1
- 5
- 90.59
- 1
- 0.000*
- 9.41▼
- +
- 0.25
- 12.5
- 89.96
- 1
- 0.000*
- 10.04▼
- 0.5
- 25
- 80.21
- 1
- 0.000*
- 19.79▼
- 1
- 50
- 76.46
- 1
- 0.000*
- 23.54▼
- Dexamethasone1
- 7.84
- 42.56
- 3
- 0.000*
- 57.44▼
- 1 Positive control
- 2 Significant difference at p<0.05 compared to solvent control without LPS(†) and with LPS(*)
Table 1: NO production in RAW 264.7 cells after treatment (n=3).
Conclusion
Aptamin C® delays the oxidation of vitamin C and maximizes its antioxidant properties, suggesting that the Aptamin C® and vitamin C complex is effective against ROS and ROS-associated diseases. In this study, we demonstrated that the Aptamin C® and vitamin C complex removes intracellular ROS. Besides, based on previous studies showing that ROS causes inflammation, we used HaCaT cells to confirm that Aptamin C® and vitamin C have anti-inflammatory effects. Overall, our results suggest that Aptamin C® could be used as a treatment for oxidative stress-associated diseases, particularly in skin and scalp care.
References
-
Edreva A (2005) Generation and scavenging of reactive oxygen species in chloroplasts: A submolecular approach. Agriculture, Ecosystems & Environment 106(2-3): 119- 133.
-
Dietz KJ ((2016) Thiol-based peroxidases and ascorbate peroxidases: why plants rely on multiple peroxidase systems in the photosynthesizing chloroplast. Molecules and cells 39(1): 20.
-
Muller F (2000) The nature and mechanism of superoxide production by the electron transport chain: its relevance to aging. Journal of the American Aging Association 23(4): 227-253.
-
Han D, Williams E, Cadenas E. (2001) Mitochondrial respiratory chain-dependent generation of superoxide anion and its release into the intermembrane space. Biochemical Journal 353(2): 411-416.
-
Sharma P, Jha AB, Dubey RS, Pessarakli M, et al. (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of botany 2012: 217037.
-
Hollis F, Alexandros KK, Claudia B (2017) Mitochondrial dysfunction in Autism Spectrum Disorder: clinical features and perspectives. Current opinion in neurobiology 45: 178-187.
-
Haider L, Fischer MT, Frischer JM, Bauer J, Höftberger R, et al. (2011) Oxidative damage in multiple sclerosis lesions. Brain 134(7): 1914-1924.
-
Patel VP, Chu CT (2011) Nuclear transport, oxidative stress, and neurodegeneration. Int J Clin Exp Pathol 4(3): 215-229.
-
Nunomura A, Castellani RJ, Zhu X, Moreira PI, Perry G, et al. (2006) Involvement of oxidative stress in Alzheimer disease. J Neuropathol Exp Neurol 65(7): 631-641.
-
Bošković M, Vovk T, Kores Plesničar B, Grabnar I (2011) Oxidative stress in schizophrenia. Current neuropharmacology 9(2): 301-312.
-
Ramalingam M, Kim SJ (2012) Reactive oxygen/ nitrogen species and their functional correlations in neurodegenerative diseases. J Neural Transm 119(8): 891-910.
-
Nijs J, Meeus M, De Meirleir K (2006) Chronic musculoskeletal pain in chronic fatigue syndrome: recent developments and therapeutic implications. Manual therapy 11(3): 187-191.
-
Handa O, Naito Y, Yoshikawa T (2011) Redox biology and gastric carcinogenesis: the role of Helicobacter pylori. Redox Report 16(1): 1-7.
-
Trouba KJ, Hamadeh HK, Amin RP (2002) Oxidative stress and its role in skin disease. Antioxidants and Redox Signaling 4(4): 665-673.
-
David RB, Mohammad A (2006) Oxidative stress in the pathogenesis of skin disease.” Journal of Investigative Dermatology 126(12): 2565-2575.
-
Rinnerthaler M, Bischof J, Streubel MK, Trost A, Richter K (2015) Oxidative stress in aging human skin. Biomolecules 5(2): 545-589.
-
Aslani FS, Dastgheib L, Banihashemi BM (2009) Hair counts in scalp biopsy of males and females with androgenetic alopecia compared with normal subjects.” Journal of cutaneous pathology 36(7): 734-739.
-
Magro CM, Rossi A, Poe J, Manhas-Bhutani S, Sadick N (2011) The role of inflammation and immunity in the pathogenesis of androgenetic alopecia. J Drugs Dermatol 10(12): 1404-1411.
-
Wadley AJ, Veldhuijzen van Zanten JJ, Aldred S (2013) The interactions of oxidative stress and inflammation with vascular dysfunction in ageing: the vascular health triad. Age 35(3): 705-718.
-
Prie BE, Iosif L, Tivig I, Stoian I, Giurcaneanu C (2016) Oxidative stress in androgenetic alopecia. J Med Life 9(1): 79-83.
-
Jadkauskaite L, Coulombe PA, Schäfer M, Dinkova-Kostova AT, Paus R, et al. (2017) Oxidative stress management in the hair follicle: Could targeting NRF2 counter age‐ related hair disorders and beyond? Bioessays 39(8): 1700029.
-
Halliwell B, John MC G (1985) Free radicals in biology and medicine. pp: 331-332.
-
Birben E, Sahiner UM, Sackesen C, Erzurum S, Kalayci O (2012) Oxidative stress and antioxidant defense. World Allergy Organ J 5(1): 9-19.
-
Tamari Y, Nawata H, Inoue E, Yoshimura A, Yoshii H, et al. (2013) Protective roles of ascorbic acid in oxidative stress induced by depletion of superoxide dismutase in vertebrate cells. Free radical research 47(1): 1-7.
-
Poljsak B, Šuput D, Milisav I (2013) Achieving the balance between ROS and antioxidants: when to use the synthetic antioxidants. Oxid Med Cell Longev 2013: 956792.
-
Pullar JM, Carr AC, Vissers MCM (2017) The roles of vitamin C in skin health. Nutrients 9(8): 866.
-
Choi S, Han J, Kim JH, Kim AR, Kim SH, et al. (2020) Advances in dermatology using DNA aptamer “Aptamin C” innovation: Oxidative stress prevention and effect maximization of vitamin C through antioxidation. J Cosmet Dermatol 19(4): 970-976.
- Epithelioid Granuloma; 3cases with Different Clinical Features
- Advancing Representation in Dermatology Clinical Trials: Ethical, Scientific, and Regulatory Imperatives for Inclusion Across all Fitzpatrick Skin Types
- A Case of Atopic Dermatitis with Concurrent Psoriasis Vulgaris: Successful Treatment with Upadacitinib
- Innovation Lifting Eyeshadow: A Synthesis of Makeup and Optical Illusion
- Distinguishing Superficial Actinic Porokeratosis from Actinic Keratosis with UVF Dermoscopy: A Case Report
- High Mobility Group Box 1 (HMGB1) in Cutaneous Inflammation: An Immune Modulator Bridging Cellular Stress, Ferroptosis and Danger Signaling