Uterine Rat Telocyte Structure and Organization: An Immunohistochemical and Ultrastructural Study
Telocytes are a kind of interstitial cell found in a variety of organs, including the uterus. They regulate uterine contractions, maintain pregnancy, and prevent premature labour. The purpose of this research was to look at the organisation and distribution of telocytes in different uterine layers in adult and senile rats at various stages of reproduction. Materials & Methods: Twenty-four female rats were evenly divided between four groups: GI: adult non-pregnant, GII pregnant, GIII postpartum, and GIV senile. The uterine samples from middle one-third of the right horns were processed for light microscopic examination were treated using C-kit stains for immunohistochemical detection of telocytes, and TEM examination. Results: Telocytes were discovered in the endometrium and myometrium of adult non-pregnant uteri as tiny cells with numerous lengthy telopodes. Endometrial telocytes increased significantly, but myometrial telocytes decreased significantly in (GII). (GIII) and (GIV) had the largest count of myometrial telocytes. Conclusion: Telocytes were found in the endometrium and myometrium of the rat uterus. Telocytes serve as a uterine peacekeeper, initiating and coordinating myometrial contraction.
Introduction
Telocytes (TCs) are a distinct group of cells identified in 1911 in the mammalian stomach by the pioneering Spanish neurologist and pathologist Santiago Ramón y Cajal. He dubbed them ‘interstitial neurons’ because to their projections and their locations between nerve terminals and smooth muscle cells [1], independently established that ‘interstitial neurons’ were not truly neurons and renamed them ‘interstitial cells of Cajal’ (ICCs) [2]. After establishing that these cells differ from interstitial cells of Cajal (ICC) and all other interstitial cells, their designation was changed to telocytes (TCs) in 2010 [3].
Telocytes have been detected in a variety of vertebrates, including humans, mice, rats, guinea pigs, and chickens, and in a variety of organs, including the pancreas, the esophagus, the small intestine, and the colon. Telocytes are also found in all of the heart’s layers, including the epicardium, the endocardium and the myocardium. Telocytes are found throughout the reproductive system, including the prostate, testesuterus, myometrium and placenta [4, 5, 6, 7, 8, 9, 10, 11, 12]. The word “telocyte” refers to a cell with lengthy protrusions on [13]. TCs feature a triangular or ovoid somatic body and many (two to five) moniliform cytoplasmic projections (telopodes) with thin segments (podomeres) and dilated parts (podoms). TCSs have an oval nucleus and a little amount of cytoplasm containing mitochondria. 5-10% of the cytoplasmic volume; 1-2% of the cytoplasmic volume is endoplasmic reticulum, which may be smooth or rough. Additionally, intermediate filaments, thin filaments, and microtubules exist [14]. Telopodes make interaction with a variety of cell types in their environment, including immune cells, muscle fibres, blood vessels, and epithelial cells [4].
TCs vary from Cajal interstitial cells and other interstitial cells (e.g. fibroblasts, fibrocytes, and fibroblast-like cells) in terms of their expression of cell surface antigens and microRNA profiles [15].
Telocytes operate as a pacemaker, generating the bioelectrical slow wave potential required for smooth muscle contraction [16]. They are involved in the regulation of smooth muscle cells’ (SMCs) contractility and excitability [17]. They may participate in the myogenic contractile process that occurs during sperm transit prior to fertilisation, embryo implantation, and delivery [18].
Although the role of uterine TCSs is unknown, experimental data suggests that telocytes may operate as modulators of spontaneous uterine contraction. This may be accomplished hormonally, since uterine telocytes express oestrogen and progesterone receptors numerous studies have suggested that telocytes may operate as sensors for sex hormone levels associated with pregnancy maintenance [11].
Due to the fact that the form and quantity of those cells alter in pathological diseases such as pre-eclampsia, endometriosis, and ovarian failure, there is a possibility that they might help to the treatment of such disorders [18]. Telocytes account for around 7% of total cells in non- pregnant myometrial cell culture and approximately 3% of total cells in the myometrium of adult non-pregnant people [3]. The purpose of this research was to determine the structure, organisation, and distribution of telocytes in the uterus of adult and senile female rats at various stages of reproduction.
Materials and Methods
Animals
Twenty-four female albino rats were housed in ordinary stainless steel crushed cages (5/cage) at Al-Azhar University’s college of medicine for females. They were maintained under careful care and sanitary conditions of temperature, relative humidity, and a 12-hour light/dark cycle. They were given food pellets from Cairo’s Factory of Oil and Soap Company, as well as certain vegetables for vitamins, which were accessible ad libitum along with drinking tap water. For one week, the rats were acclimated to the laboratory setting. They were divided into four groups, each with six rats.
Adult non-pregnant rats weighing 170–190 gm were classified as: Group I (GI) Adult pregnant rats in Group II (GII) on days 16–18 of gestation. They were around three to four months old and weighed between 230 and 270 grammes. The presence of a vaginal plug on day 1 of gestation was used to determine the gestational day. The average time of delivery on day 23 was in the AM. Adult postpartum rats in Group III (GIII) on day 3 postpartum. They were around three to four months old and weighed between 180 and 200 grammes. Group IV (GIV): Senile group, aged around 18-24 months and weighing between 280-330 gm.
All rats in each group were killed while sedated with ether. The abdomens were opened midline, the uteri were dissected, and the middle one-third of the right uterine horns (or the implantation sites of the middle one-third if pregnant rats) were removed.
Immunohistochemistry Using c-kit (CD117)
The sections were routinely immunohistochemically prepared and incubated overnight at 4 °C with primary antibodies against c-kit (CD117) diluted in 1% bovine serum albumin in phosphate-buffered saline (PBS; pH 7.4) (DAKO corporation laboratories, Carpinteria, CA 93013, USA supplied the primary antibody and antibody diluent). They were kept between 2 and 8 degrees Celsius). Negative controls were handled in the same way as positive controls, except that the main antibody was replaced with PBS. The cells that were positive for the c-kit had brown cytoplasmic deposits and blue nuclei [19].
Electron Microscopic Examination (TEM)
Tiny pieces (about 0,5 mm3) of the central one-third of the right uterine horns were removed and promptly treated with 3 percent glutaraldehyde and stored at 0 -4 degrees Celsius for 24 hours [20]. The sections were cut with a diamond knife on a LKB ultramicrotome. To begin, 0.5 m semithin sections were cut, picked up on a glass slide, and stained with toluidine blue for light microscopic examination; ultrathin sections (80nm) were cut and picked up on 200 mesh copper grids [21], stained with uranyl acetate [22], and lead citrate stain [20]. Sections were inspected and photographed using a transmission electron microscope (JEOL 100S Tokyo, Japan) at electron microscopic facilities at AL- Azhar University’s college of medicine for females.
Morphometric and Statistical Studies
Morphometric measurements were made using a computerised image system that included a Leica Quin 500 image analyzer coupled to a Leica microscope. Telocytes (mean number of CD117 immunopositive cells) were counted in the endometrium and myometrium of each group tested. Ten non-overlapping fields from each group were inspected at a magnification of x100 using light microscopy relayed to the screen.
The whole statistical study was carried out using the statistical programme “Statistica for Windows” Version 5. The statistical analysis was conducted using the mean (M) and standard deviation (S.D) values as specified in [23]. When the probability (p) was 0.05 or more, the results were judged significant; when the probability (p) was 0.01 or greater, the results were regarded very significant [24].
Results
Immunohistochemical Results
The uterus’s cross-sections consisted of three layers from the inside outward: endometrium, myometrium, and perimetrium. Historically, the endometrium was composed of a single layer of simple columnar epithelium that was overlaid by a thick layer of lamina propria containing endometrial glands. Three layers comprised the myometrium: the inner circular (IC), the middle stratum vascular (SV), and the outer longitudinal layers (OL). Immunohistochemical staining demonstrated a high number of c-kit-positive cells in all levels of the uterine wall of all groups investigated (Figure 1).
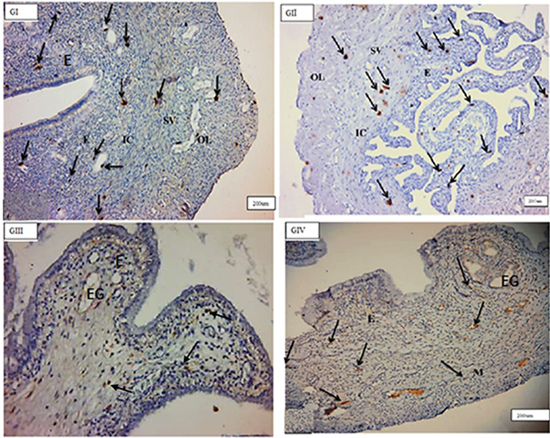
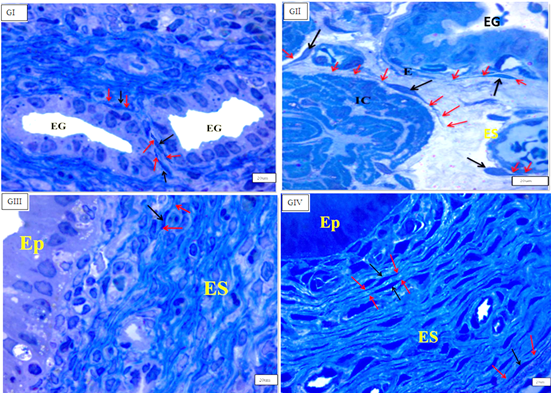
Figure 1: Photomicrographs of cross section in the rat uterus showing many c-kit-positive cells (arrows) in the endometrium (E) in–between endometrial stromal cells, near to blood vessels and around endometrial gland (EG), inner circular(IC), middle stratum vasculare(SV) and outer longitudinal(OL) smooth muscle layers in(GI), (GII),(GIII), (GIV). (C-kit immunohistochemistry counterstained with Hx, (GI, GII, GIV) & (GIII), O.M, X100&200 respectively).
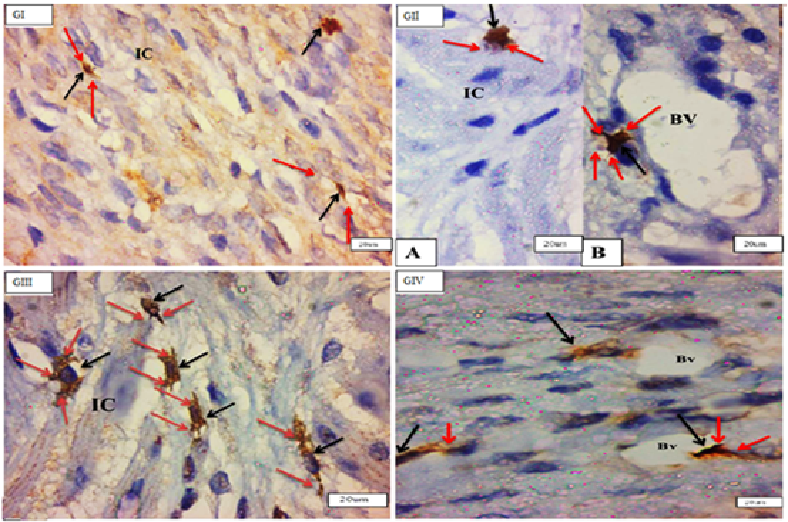
Endometrial telocytes exhibited as tiny, spindle- or pyriform-shaped cells with sparse cytoplasm and big oval nuclei with one or two long thin processes when stained with C-kit (CD117) (telopodes). Telocytes from several c-kit- positive bodies were often seen in close proximity to blood arteries, surrounding endometrial glands, and between stromal cells in all investigated groups, but primarily in (GII) endometrium (Figure 2).
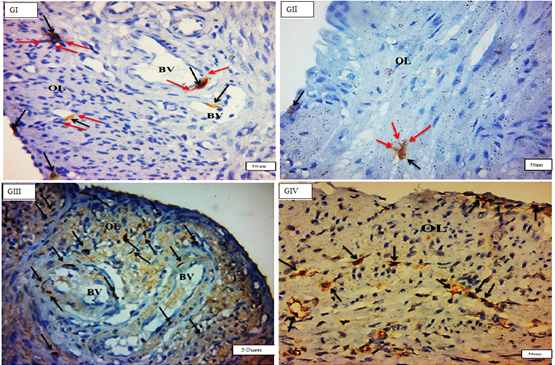
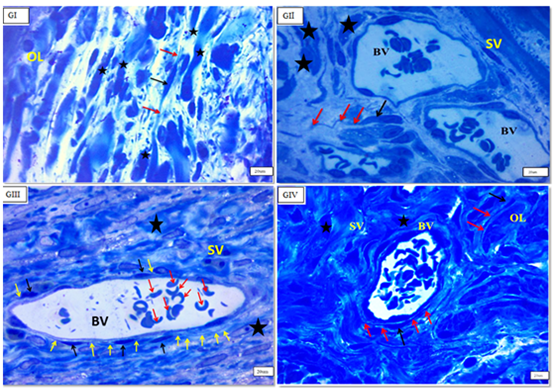
TCSs were discovered mostly in the myometrium’s (IC) layer, where c-kit-positive telocytes were organised parallel to smooth muscle fibres. Telocytes were also found in close proximity to blood vessels in the myometrium’s (SV) layer and between smooth muscle fibres in the myometrium’s (OL) layer. There was a significant rise in the quantity of c-kit-positive telocytes in the myometrium, particularly in the (OL) layer, in (GIII). They ran parallel to the (IC) smooth muscle and bordered and interspersed the (OL) muscles. Immunostained slices of the uterus of senile rats (GIV) exhibited many c-kit-positive cells oriented parallel to one another in the (IC) layer of the myometrium. More prevalent in (GIII) than in (GI) (Figures 3 & 4).
Electron Microscopic Results
Semi-thin uterine sections stained with toluidine blue revealed the typical form of interstitial cells telocytes with tiny oval bodies and long thin processes (telepodes) in the endometrium, particularly surrounding endometrial glands, and in the endometrial stroma (Figure 5). TCs in the myometrium exhibited a tiny body and distinctive telopodes located between myocytes of all groups, but primarily in (GIII) (Figure 6).
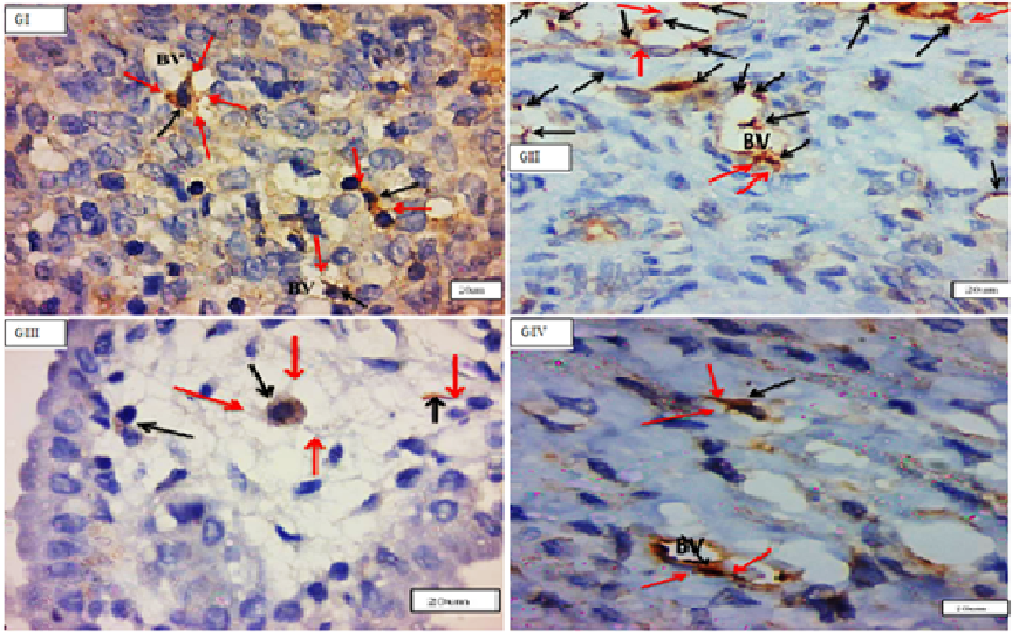
By TEM, the TCs had a tiny body surrounded by a small amount of cytoplasm including a central nucleus, organelles such as mitochondria and rough endoplasmic reticulum, polyribosome aggregations or glycogen granules, and other electron-dense entities. Telopodes (1–4) are very long cellular extensions with alternating thin segment podomers and thick sections podoms that stretch from the body. TCSs were discovered in the endometrium lamina propria, just under the simple columnar epithelium, and surrounding the endometrial gland. TCs were not present in the basal lamina of (GII). There was a direct relationship between telopodes and longitudinal and transverse sections of collagen fibres along their course. Numerous telocytes were discovered in the myometrium, mostly in the vicinity of blood vessels and between smooth muscle cells (Figures 7-9).
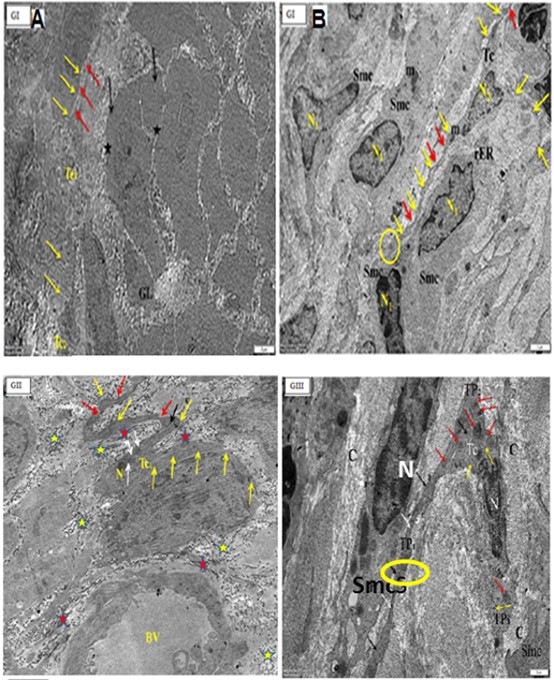
Figure 7: Electron micrographs of cross sections in the uterus of adults non-pregnant rat (GI) (A) showing endometrial stroma containing endometrial gland with lumen (GL) with epithelium (black arrows) and nuclei (star). There are two podomers (yellow arrows) and podoms (red arrows) along the telopodes. (B): showing parallel arranged smooth muscle cells (SMCs) with nuclus (N1), Telocyte (TC) is present inbetween smooth muscles with a small body containing the nucleus (N2), and three apparent telopodes; (red arrows), podoms (yellow arrows) and the encircled area showing the close connection of telopodes with the adjacent Smc. Note, mitochondria (m), rough endoplasmic reticulum (Eer). (O.M, x4000).
Figure 8a: Electron micrographs of cross sections in: in the uterus of A pregnant rat (GII) showing telocyte near to blood vessels (BV). Tc body with nucleus (N), mitochondria (white arrows), rER(black arrow) and twisted, long telopodes with podoms (yellow arrows) and podomers (red arrows). Note, collagen fibers, longitudinal(red star) and transverse (yellow stars) sections.
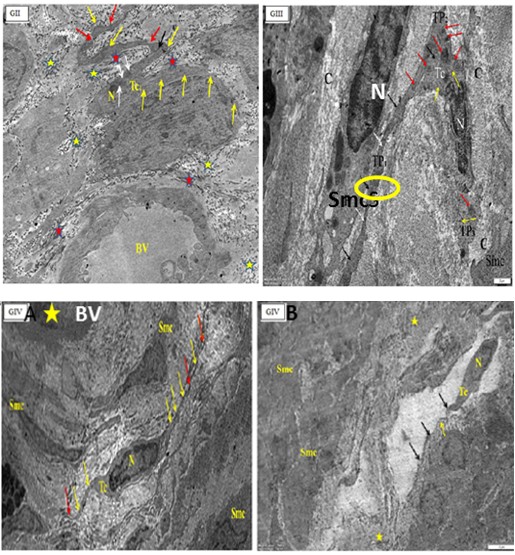
Figure 8b: Electron micrographs of cross sections in: in the uterus of A pregnant rat (GII) showing telocyte near to blood vessels (BV). Tc body with nucleus (N), mitochondria (white arrows), rER(black arrow) and twisted, long telopodes with podoms (yellow arrows) and podomers (red arrows). Note, collagen fibers, longitudinal(red star) and transverse (yellow stars) sections.
B postpartum rat (GIII) showing a part of the myometrium containing (TC) with body and nucleus (N), three telopodes (TP1,2&3), podomes (black arrows), podomers (white arrows). Note mitochondria (red arrows), aggregations of polyribosomes or glycogen granules (yellow arrows). Smooth muscles cells (SMCs) with nucleus (N), collagen fibers (C). The yellow circle showing close connection of telopode with SMC (O.M, x X10000-8000 respectively).
Figure 9: Electron micrographs of cross sections in the uterus of senile rats (GIV) (A) showing (TC) with nucleus (N) and long telopodes, Note, podoms (yellow arrows), podomers (red arrows), blood vessel (BV) with its endothelian linning (star). (B) showing (Tc) with its cell body, nucleus (N) and telopode (arrows) in the myometrium. Note, smooth muscles cells (Smcs), collagen fibers(star), and podoms (black arrows). (O.M X3000).
Morphometric and Statistical Results
The mean number of TCSs per ten HPF is determined. Image analysis of the current data indicated that the mean number of TCSs in the endometrium of (GI) was 8.291.11, (GII) was 131.16, (GIII) was 8.291.26, and (GIV) was 8.571.27. These data indicated a rise in a statistically meaningful manner (GII). As indicated in table (1) and histogram (1), the P-value was 0.05. While the mean number of TCSs in the (GI) myometrium was 10.29.76, (GII) myometrium was 5.43.54, (GIII) myometrium was 15.14.69, and (GIV) myometrium was 12.291.98. These data indicated that (GIII) & (GIV) increased statistically significantly, whereas (GIV) decreased statistically significantly (GII). Table 1 and histogram (1) illustrate the P values (P 0.05).
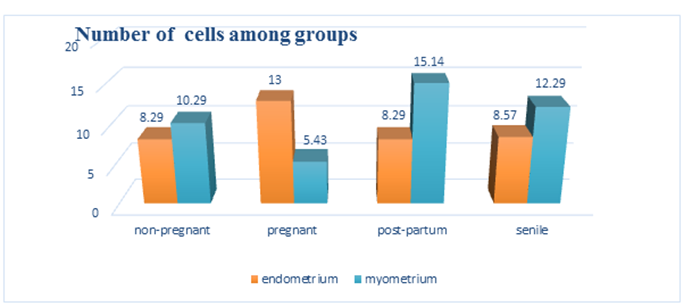
Histogram1: Comparison between the four studied groups as regard number of telocytes/1 in the endometrium and myometrium of adult and senile female albino rats.
| Non pregnant (A) M±SD | Pregnant (B) | Postpartum (C) M±SD | Senile (D) | P-value | Post hoc | |
|---|---|---|---|---|---|---|
| (GI) | M±SD | (GIII) | M ± SD | |||
| (GII) | (GIV) | |||||
| Endometrium | 8.29±1.113 | 13 ± 1.155 | 8.29 ± 1.254 | 8.57 ± 1.27 | 0 | B-A, B-C |
| B-D | ||||||
| Myometrium | 10.29 ± .756 | 5.43 ± .535 | 15.14 ±.690 | 12.29±1.976 | 0 | A-B,A-C |
| A-D |
Table 1: Statistical analysis of the number of telocytes / 10 HPF in the endometrium and myometrium of adult and senile female al
Discussion
Telocytes TCs, previously known as interstitial Cajal- like cells (ICLCs), have been described in practically every organ of the human body in recent years [25]. The present study demonstrated the presence of TCs in the endometrium and myometrium of rat uterus in various reproductive states (adult non-pregnant (GI), pregnant (GII), postpartum (GIII), and senile (GIV)) using toluidine blue staining, immunohistochemistry with anti-tic-kit antibodies, and transmission electron microscopic analysis.
Telocytes are tiny, pyriform, or spindle-shaped cells with long, thin, and sparse telopodes (2–5). This is consistent with Cristian, et al. [26] findings about TCs in the subepicardial habitat. According to Przemysław, et al. [18]. Telopods may reach a length of 1000 m, making them one of the longest structures in the body, save for certain axons. Furthermore, unlike CD 34, c-kit was shown to stain primarily the cell body of TCs with a lower affinity for staining the cell processes. Salama, et al. [27] and Ivan, et al. [17] concur on this point. With their long and thin cell bodies, semithin slices of uterine fragments stained with toluidine blue were suitable for assessing the distribution of TCs throughout the uterus (Tps). This is consistent with the findings of Zheng, et al. [28] who investigates uterine telocytes.
Telocytes were discovered using TEM in the endometriums of all examined groups, mostly near endometrial glands, endometrial blood vessels, and between endometrial stromal cells. This is consistent with the findings of Przemysaw, et al. [18] who investigated TCs in the female reproductive system and established that uterine stromal cells. It is possible that TCs act as a scaffold for endometrial maintenance, glandular support, and stromal cell communication. Additionally, TCs are abundant around blood vessels, where they are involved in tissue homeostasis, remodelling, assisting in the development of new blood vessels (angiogenesis), suppressing oxidative stress and cellular ageing, and protecting against inflammation and oncogenesis [29].
There was no significant difference in the number of TCs in the endometrium of (GI) and (GIII) patients in this investigation, although the greatest count of endometrial TCs was observed in (GI) (GII). These findings corroborated those of Przemysaw, et al. [18]. Additionally, there was no significant difference in the quantity of TCs in the endometrium of (GIV) patients vs controls (GI). The morphometric analysis corroborated these findings, and Hatta, et al. [30], postulated that TCs are typically found in tissues with a low cell density and large space between surrounding cells. This hypothesis may be supported by the present finding that the pregnant group had a significantly higher number of endometrial TCs than the other groups, as the endometrium, unlike the myometrium, becomes looser and loses cellularity during pregnancy, necessitating the presence of more TCs to facilitate cell-to-cell contact over long distances [27, 18].
Additionally, the present study established the existence of c-kit-positive TCs in the myometrium of all groups investigated. This data corroborates Veronika, et al. [31]’s discovery that TCs comprised around 7% of the overall cell population in non-pregnant myometrial cell culture and approximately 3% of the total cell population in the myometrium of adult non-pregnant individuals.
Myometrial TCs were indicated to play a critical role in the production and coordination of myometrial contractility in a c-kit-dependent manner [32]. Additionally, TCs possess excitatory and inhibitory neurotransmitter receptors and are capable of transmitting nerve impulses to smooth muscle cells, where they participate in mechanoreception [29]. Experiments indicated that TCs may be involved in the spontaneous contraction of the uterus [11]. This may have happened as a result of the hormonal influence, since uterine TCs have been shown to express oestrogen and progesterone receptors, operate as steroid sensors, and contribute to the coordination of human myometrial contractions and pregnancy maintenance [26, 32].
The current morphometric analysis demonstrated that the number of myometrial TCs was substantially lower in (GII) and significantly greater in (GIII) and senile uteri (GIV) compared to adult non-pregnant uteri (GI). This observation might be explained by the fact that the number of myometrial TCs is decreased during pregnancy to avoid early uterine contractility and preterm birth, but the number of myometrial telocytes is raised postpartum to promote myometrial contraction during uterine involution [18, 27]. There are no published data on the number of TCs in the senile uterus, but it may have risen to adjust to the wide muscle separation caused by extra collagen fibers or may have reacted to any low levels of steroid hormones due to its estrogen and progesterone receptors.
Telocytes make extensive connections with neighboring cells, forming a unique three-dimensional network inside interstitial tissues. These structural characteristics underpin the suggested numerous roles of TCs [32]. Additionally, uterine TCs form linkages with other extracellular matrix components (for example, collagen fibers) [18].
Conclusion
Telocytes increased in the endometrium of pregnant uteri and the myometrium of postpartum and senile uteri, but decreased in the pregnant uterus myometrium.
Recommendation
Additional research is required to describe uterine TCs during parturition. Gaining a better knowledge of uterine TCs may aid in the development of therapeutic options for dysmenorrhea, recurrent pregnancy loss, and preterm delivery.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
-
Cajal RS (1911) Histologie du systeme nerveux de l’Homme et de Vertebres. Paris: Maloine.
-
Popescu LM, Ciontea SM, Cretoiu D, Hinescu ME, Radu E, et al. (2005) Novel type of interstitial cell (Cajal-like) in human fallopian tube. J Cell Mol Med 9(2): 479-523.
-
Popescu LM, Manole CG, Gherghiceanu M, Ardelean A, Nicolescu MI, et al. (2010) Telocytes in human epicardium. J Cell Mol Med 14(8): 2085-2093.
-
Kondo A, Kaestner KH (2019) Emerging diverse roles of telocytes. Development 146(14): dev175018.
-
Chen X, Zheng Y, Manole CG, Wang X, Wang Q, et al. (2013) Telocytes in human oesophagus. J Cell Mol Med 17(11): 1506-1512.
-
Hennig GW, Spencer NJ, Jokela-Willis S, Bayguinov PO, Lee HT, et al. (2010) ICC-MY coordinate smooth muscle electrical and mechanical activity in the murine small intestine. Neurogastroenterol Motil 22(5): 138-151.
-
Klein M, Csöbönyeiová M, Žiaran S, Danišovič Ľ, Varga I, et al. (2021) Cardiac Telocytes 16 Years on—What Have We Learned So Far, and How Close Are We to Routine Application of the Knowledge in Cardiovascular Regenerative Medicine. Int J Mol Sci 22(20): 10942.
-
Kostin S (2010) Myocardial telocytes: a specific new cellular entity. J Cell Mol Med 14(7): 1917-1921.
-
Corradi LS, Jesus MM, Fochi RA, Vilamaior PS, Justulin LA Jr, et al. (2013) Structural and ultrastructural evidence for telocytes in the prostate stroma. J Cell Mol Med 17(3): 398-406.
-
Gomes VLA, Braz JKFDS, Martins GM, Clebis NK, Oliveira MF, et al. (2021) Identification of telocytes in dystrophic mice testis. Einstein (Sao Paulo) 19: eAI5737.
-
Creţoiu SM, Creţoiu D, Popescu LM (2012) Human myometrium – the ultrastructural 3D network of telocytes. J Cell Mol Med 16(11): 2844-2849.
-
Suciu L, Popescu LM, Gherghiceanu M, Regalia T, Nicolescu MI, et al. (2007) Telocytes in human term placenta: morphology and phenotype. Cells Tissues Organs 192(5): 325-339.
-
Wang J, Jin M, Ma WH, Zhu Z, Wang X, et al. (2016) The History of Telocyte Discovery and Understanding. Adv Exp Med Biol 913: 1-21.
-
Vannucchi MG (2020) TheTelocytes: Ten Years after Their Introduction in the Scientific Literature. An Update on Their Morphology, Distribution, and Potential Roles in the Gut. Int J Mol Sci 21(12): 4478.
-
Wang L, Xiao L, Zhang R, Huiling Jin, Haixia Shi, et al. (2020) Ultrastructural and immunohistochemical characteristics of telocytes in human scalp tissue. Scientific Reports 10: 1693.
-
Díaz-Flores L, Gutiérrez R, García MP, Gayoso S, Gutiérrez E, et al. (2020) Telocytes in the Normal and Pathological PeripheralNervous System. Int J Mol Sci 21(12): 4320.
-
Varga I, Polák Š, Kyselovič J, Kachlík D, Danišovič Ľ, et al. (2019) Recently Discovered Interstitial Cell Population of Telocytes: Distinguishing Facts from Fiction Regarding Their Role in the Pathogenesis of Diverse Diseases Called “Telocytopathies”. Medicina (Kaunas) 55(2): 56.
-
Janas P, Kucybała I, Radoń Pokracka M, Huras H (2018) Telocytes in the female reproductive system. Adv Clin Exp Med 27(4): 559-565.
-
Bancroft J, Gamble M (2015) Theory and Practice of Histological Techniques (6th Edn) Churchill Livingstone Edinburugh London Melbourne pp: 201-217.
-
Robinson D, Ehlers U, Herken U, Herrmann B, Mayer F, et al. (1987) Method for preparation for electron microscopy. An introduction to the biochemical sciences. Muhlethaler K. Springer-verlag. Berlin, Heidelberg, New York-London-Paris-Tokyo.
-
William W, Carter C (1996) Transmission electron microscopy. A Text Book For Material Science, (2nd Edn). Springer Verlag. Heidelberg pp: 721-756.
-
Johanneseen J (1978) Electron microscopy in humn medicine. Instruments and Techniques. McGraw Holl international book company 1: 20-57.
-
Mould R (1989) Introductory medical statistics. 2nd (Edn.), Adam Hilger, Bristol, Philadelphia 17: 22.
-
Armitage P, Berry G (1994) Statistical methods in medical research. 3rd Oxford Blackwell Scientific Publication pp: 1-10.
-
Soliman SA (2021) Telocytes are major constituents of the angiogenic apparatus. Sci Rep 11(1): 5775.
-
Cristian B, Mugure C, Laurenţiu M, Sorin H, Oana D, et al. (2019) The Telocytes in the Subepicardial Niche. Appl Sci 9(8).
-
Salama N (2013) Immunohistochemical characterization of telocytes in rat uterus in different reproductive states. Egyptian Journal of Histology 36 (1): 185-194.
-
Zheng Y, Bai C, Wang X (2012) Telocyte morphologies and potential roles in diseases. J Cell Physiol 227(6): 2311-2317.
-
Abdel Maksoud F, Abd Elhafeez H, Soliman A (2019) Morphological changes of telocytes in camel efferent ductules in response to seasonal variations during the reproductive cycle. Sci Rep 9: 4507.
-
Hatta K, Huang ML, Weisel RD, Li RK (2012) Culture of rat endometrial telocytes. J Cell Mol Med 16(17): 1392- 1396.
-
Aleksandrovych V, Walocha JA, Gil K (2016) Telocytes in female reproductive system (human and animal). J Cell Mol Med 20(6): 994-1000.
-
Tang XL, Zhang FL, Jiang XJ, Yang XJ (2019) Telocytes enhanced the proliferation, adhesion, and motility of endometrial stromal cells as mediated by the ERK pathway in vitro. Am J Transl Res 11(2): 572-585.
- Huddled and Preternatural- Atypical Lobular Hyperplasia
- Cytologic Surveillance Versus Immediate Referral to Colposcopy for Women with a Cervical Cytology Diagnosis of ASC-US and LSIL in the Absence of HPV DNA Test
- Callow and Vernal-Hepatoblastoma
- Black Plum Fruit Extract as a Substitute for Haematoxylin in Haematoxylin and Eosin Staining Technique
- Critical Analysis of World Status of Research on the Development of High Yielding Strains of Jatropha Curcas L
- Non-Invasive Follicular Thyroid Neoplasm with Papillary-Like Nuclear Features: Pitfalls of Cytological and Histopathologic Diagnosis