Coconut Oil, Soybean Oil, Lard: Good or Bad? The link to Obesity
Obesity is a worldwide public health problem. Obesity and adiposity are caused by different factors but many cases are as a result of overconsumption of high-fat diets. Fat and oil in the diets are made available to the body in form of fatty acids. Considering the degree of saturation, fatty acids can be classified into saturated fatty acids (SFA), monounsaturated fatty acids (MUFA) and polyunsaturated fatty acids (PUFA). There have been controversies as regards the consumption of different fats and oils, which is essential for the establishment of public health policies to control the prevalence of metabolic disorders in the global population. The purpose of this review is to examine data from different studies and dietary intervention performed in animal models, as well as humans with the consumption of coconut oil, soybean oil, lard and their link to obesity. In addition, it sheds light on the intracellular metabolism and the changes in gene expression caused by the different dietary fat, elucidating the sex differences in obesity. It also emphasizes the public health policies and the different nutritional recommendations associated to the consumption of dietary fats. Taken together, the different studies showed evidence that high intake of coconut oil, soybean oil or lard promotes obesity, adiposity and insulin resistance through changes in the expression of inflammatory genes in a sex-dependent manner. However, it was revealed that soybean oil is more obesogenic than coconut oil and lard.
Introduction
The global increase in the rate of obesity and its associated co-morbidities (diabetes and cardiovascular diseases) has been alarming overtime. In the year 2011, the rate of obesity reported was 1.46 billion overweight people (body mass index, BMI>25) [1]. Later on, the World Health Organization (WHO) reported approximately 1.9 billion individuals with overweight in 2017 in the world [2]. Given this scenario, obesity has been reported as a major global health issue [3], with approximately 3.4 million of deaths worldwide reported as a result of obesity [2]. Body mass index is generally used to classify the different classes of obesity.
Factors contributing to obesity include genetics and environmental factors. However, the most relevant contributing factor is diet, which results in obesity from the number of calories intake and the origin of those calories. There is increasing evidence of the link between dietary fats and oils and obesity, with the first reports far back to four decades ago [4, 5]. As a result of this link, nutritional guidelines were developed to encourage people to limit the intake of dietary fat especially saturated fatty acids but in recent times, some studies have been questioning these nutritional guidelines and these studies are encouraging the reconsideration of the nutritional guidelines [6, 7]. Poudyal and Brown examined the effects of different dietary fatty acids on cardiometabolic risk factors and reported that there are contradiction between the chemical and the biological responses. In the review, dietary fatty acids was classified into five groups based on their effects on cardiometabolic risk, the group includes neutral, reduce one or more cardiometabolic risk factors, increase on or more cardiometablic risk factors [8]. Despite the high rate of production of coconut oil in Brazil, Brazilian domestic demand for coconut oil is met by additional imports from Philippines and Indonesia [9]. Brazilian domestic consumption of soybean oil in 2019 amounted to approximately 7.3 million metric tons. Comparing this to year 2010, there is an increase of over 40 percent in consumption [10]. Lard is included in this review basically because of the consumption rate in Brazil. Brazil is the third largest producer of Lard with 481 thousand tonnes produced annually [11]. The rationale for focusing on these three fat sources (coconut oil, soybean oil and lard) is that they are highly consumed in Brazil due to social, economic and cultural reason. Therefore, this review considers various studies, emphasizing coconut oil, soybean oil and lard consumption and its relationship to obesity and other metabolic disorders. Studies that discussed any of the three fat sources or compare two or the three fat sources and its link to obesity and other related metabolic disorders were reviewed. The studies were obtained from electronic databases Pubmed and Google scholar and we focus on studies involving animal models and human clinical trials. The intracellular metabolism of the different dietary fat, some approved drugs that inhibit or trigger the metabolic process and the changes in the gene expression after the intake of dietary fat were also discussed. Finally, an overview of the sex differences in obesity was also discussed.
An overview of Coconut oil, Soybean oil and Lard
Coconut oil The coconut (Cocos nucifera) tree, which is often referred to as ‘tree of life’, is an important food in some tropical countries. Coconut and its product (oil and milk) are used in such places to serve different purposes which include cooking, skin and hair treatment and medicine [12]. The cultivation of coconut is predominant in Asia representing about 84% of the global production in 2016. In the year 2020, Indonesia has been the largest coconut producing country with 17.72 million tons, and then Philippines (13.83 million tons), India (11.13 million tons), Brazil (2.65 million tons) and Sri Lanka (2.52 million tons) [13]. Coconut oil, which is the main product of coconut, has attracted recognition worldwide. Coconut oil is composed of the fatty acids, caprylic acid, C-8:0 (8%); capric acid, C-10:0 (7%); lauric acid, C-12:0 (49%); myristic acid, C-14:0 (8%); palmitic acid, C-16:0 (8%); stearic acid, C-18:0 (2%); oleic acid, C-18:1 (6%); and 2% of C-18:2 linoleic acid. In total, coconut oil has 92% saturated fat. Some research and review articles as well as non-scientific blogs are encouraging the consumption of coconut oil due to claims that it has some health benefits which include cholesterol lowering effect, cardiovascular diseases risk reduction, weight reduction, action as an antimicrobial, antifungal and antiviral agent, immune system enhancement [12, 14, 15, 16]. In an attempt to market coconut oil based on the supposed health benefits, many producers and marketers put in their labels, allegations such as ‘good for cooking’ [17, 18], ‘healthiest cooking oil’ [19]. In addition, coconut oil contain phytosterols which are reported to have some health benefits such as decrease in cholesterol absorption, reduction in low-density lipoproteins (LDL) and also its anti-inflammatory effects [20]. Martins and colleagues reported the presence of two phytosterols (stigmasterol and campesterol) in coconut oil [21].
With this recent development, the sales of coconut oil increased worldwide, as did so the consumption of coconut oil in the USA (34% from 2004 to 2014) [18]. However, with all this beneficial effects of coconut oil purported, high intake of coconut oil should be discouraged considering its saturated nature. The studies performed in the last 25 years describing the main findings on induction of obesity and other related metabolic disorders through coconut oil, soybean oil and lard intake are summarized in Table 1.
| Publications | Experimental model | Sex | Duration | Intervention | Reported Effect |
|---|---|---|---|---|---|
| Coconut oil vs Soybean oil | |||||
| Assunção et al. [30] | Humans | F | 12 weeks | Daily dosage of 30 ml of either soybean oil or coconut oil. | The group that received soybean oil presented an increase in total cholesterol, LDL, and LDL:HDl ratio when compared to the group that received coconut oil. The group that received Coconut oil exhibited a reduction in waist circumference (abdominal obesity) when compared with the group that received soybean oil. |
| Ippagunta et al. [150] | Mice (ICR ) | M | 52-54 days | Determination of the effects of conjugated linoleic acid in mice fed with coconut oil or soybean oil diets. Mice were fed diets containing soybean oil, coconut oil or fat free for 6 weeks followed by 10-12 days of conjugated linoleic acid supplementation. | It was observed that dietary conjugated linoleic acid induces lipolysis in adipose tissue of coconut oil-fed mice but not in soybean oil-fed mice. |
| Diaz- Villasenor et al. [151] | Rats (Zucker (fa/fa)). | M | 8 weeks | Differential effects of the interaction of the type (coconut oil or soybean oil) and amount (5% or 10%) of fat with the type of dietary protein (casein or soy protein) on the functionality of white adipose tissue of Zucker (fa/fa) rats. | It was observed from the result that soybean oil (10% fat) reduced adipocyte size and decreased esterified saturated fatty acids in white adipose tissue when compared with coconut oil fed mice. The results also showed that the type and amount of dietary fat modify the activity of white adipose tissue even in genetic model of obesity. |
| Ippagunta et al. [152] | Mice (ICR) | M | 52-56 days | Determination of the effects of conjugated linoleic acid (CLA) on soybean oil or coconut fed mice. | It was observed that coconut oil fed mice had greater fatty acid synthase and stearyl CoA desaturase 1 mRNA expression and less acetyl CoA carboxylase mRNA expression. It was also observed that sterol regulatory binding protein 1c was decreased by conjugated linoleic acid (CLA) in coconut oil (CO) fed mice and increased in soybean oil (SO) fed mice. It was reported that CLA- induced lipolysis occurs more rapidly in CO vs SO-fed mice and lipogenesis is decreased in CO-fed mice. |
| Deol et al. [38] | Mice (C57BL/6) | M | 32 weeks | Administration of series of isocaloric diets which include, soybean oil, coconut oil and fructose (40 kcal% fat) to investigate the effects of saturated versus unsaturated fat, as well as fructose, on obesity and diabetes | It was observed that soybean oil is more obesogenic and diabetogenic than coconut oil and fructose in mouse. |
| Patrone et al. [153] | Mice (C57BL/6N) | M | 8 weeks | Examination of the diversity and metabolic capacity of the cecal bacterial community in mice after the administration of either coconut oil or soy oil | The study shows significant differences in cecal predictive functions of mice. These differences can be linked to host lipid metabolism and energy storage and this give an insight to the role of gut microbiota contributing to metabolic disorders |
| Olivares- Garcia, et al. [154] | Rats (Sprague- Dawley) | M | 21 days | Assessment of the postprandial regulation of adipokine secretion in nonobese rats that consumed high-fat diet (HFD) composed of different types of fat (coconut oil, safflower oil or soybean oil). The serum concentrations of adiponectin, leptin, retinol- binding protein-4 (RBP-4), vistatin and resistin were determined at fasting and after refeeding. Adiponectin multimerization, intracellular localization, expression of endoplasmic reticulum (ER) chaperons and transcriptional regulators were evaluated in epididymal white adipose tissue | It was observed that; with coconut oil, all the three multimeric forms were reduced; with safflower oil, only the high-molecular- weight (HMW) and medium- molecular-weight (MMW) forms were reduced; and with soybean oil, only the HMW form was decreased. As a result of all these reduction noticed, it was reported that consumption of any of these oils differentially affect the abundance of ER resident protein 44 kDa (ERp44), sirtuin 1 (SIRT 1) and peroxisome proliferator-activated receptor-γ (PPARγ) all of which has been linked to obesity and are involved in post-translational processing of adiponectin required for its secretion. |
| Feranil et al. [155] | Humans | F | 22 years | Examination of the association between coconut oil consumption and lipid profiles in 1,839 Filipino women | It was reported that dietary coconut oil intake was positively associated with HDL-c levels |
| Kinsella et al. [156] | Humans | F = 18 | Administration of 25g of either coconut oil or medium chain triglycerides oil to 24 persons | It was observed that coconut oil promoted a smaller satiety compared with the medium chain triglycerides oil. The MCT oil group had a reduced food intake throughout the day compared with the group that received coconut oil. | |
| M = 6 | |||||
| T = 24 | |||||
| Deol et al. [157] | Mice (C57BL/6N) | M | 24 weeks | Assessing the linked of soybean oil and coconut oil to obesity and investigating the dysregulation of hypothalamic gene expression and the oxytocinergic system. Male mice were fed with one of coconut oil (40% fat, HFD), soybean oil high in linoleic acid, LA (40% fat, HFD), low LA soybean oil. | It was observed that the soybean oil diets (both high and low linoleic acids, LA) causes alterations in hypothalamic Oxt and plasma oxytocin, however these alterations were not observed in the group fed with coconut oil. It was postulated that LA is not responsible for the effects of soybean oil diets on oxytocin. It was noted that soybean oil is more obesogenic than coconut oil. It was also reported that given the presence of soybean oil in the American diet, its observed effects on hypothalamic gene expression could have important public health ramifications. |
| Soybean oil vs Lard | |||||
| Ikemoto et al. [60] | Mice (C57BL/6J) | F | 19 weeks | Comparing the differential effects of palm oil, lard, rapeseed oil, soybean oil, safflower oil and tuna fish oil in mice. Mice receive each of these as high-fat- diet (60%). | The body weight induced by the different diet in the following order; soybean oil > palm oil ≥ lard ≥ rapeseed oil ≥ safflower oil ≥ perilla ˃ fish oil. It was confirmed from this study that high intake of soybean oil leads to obesity and also a risk factor for dysregulation of glucose tolerance. |
| Wang et al. [66] | Rats (Sprague- Dawley) | M | 8 weeks | Differential effects of high-fat-diet rich in lard oil or soybean oil on osteopontin expression and inflammation of adipose tissue in diet induced obese rats. Rats were fed with either lard or soybean oil and later switched to low- fat diet. | It was observed that reducing lard oil with soybean oil in high- fat diets or reducing it alleviates obesity-related inflammation and insulin resistance by attenuating the upregualtion of osteopontin and macrophage infiltration into adipose tissue induced by high-fat diet. |
| Zhao et al. [158] | Rats (Sprague- Dawley) | M | 18 weeks | Investigation of the effects of high-fat-diet enriched with lard oil or soybean oil on liver endoplasmic reticulum (ER) stress and inflammation markers in diet-induced obese (DIO) rats and estimation of the influence of following low-fat diet feeding. Lard oil and soybean oil are used to induce obesity and their deleterious effects considered. | It was observed that the markers of ER stress were significantly increased in rats fed with high lard compared to high soybean oil and low-fat diets. It was reported that switching to low-fat diet reduced liver fat deposition. From the results, it was reported that high intake of lard oil, soybean oil and low-fat diet induced obesity. However, lard oil was more deleterious in insulin resistance and hepatic steatosis via promoting ER stress and inflammation responses in DIO rats. It was also reported that low-fat diet was confirmed to be useful in recovering from impaired insulin sensitivity and liver fat deposition. |
| Wang et al. [159] | Mice (C57BL/6J) | M | 12 weeks | Evaluating the anti-obesity effect of a traditional Chinese dietary habit. Mice were repeatedly fed with 3.8% or 6.5% of either soybean oil, lard or the oil blended by both. | It was observed in this study that oil mixture containing lard and soybean oil had a remarkable anti- obesity effect which suggests that the Chinese dietary habits of using oils blended with lard and soybean oil might be responsible for the lower percentages of overweight and obesity in China. |
| Henkel et al. [62] | Mice (C57BL/6) | M | 20 weeks | Examining the effect of soybean oil and lard on nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH) which are hepatic manifestations of the metabolic syndrome. Western diet containing either soybean oil (25g/100g n-6-PUFA) or lard (21g/100g). | It was reported that soybean oil induced steatosis, inflammation and fibrosis accompanied by hepatic lipid peroxidation and oxidative stress in livers of the mice, which in addition showed increased weight gain and insulin resistance. Also, lard and supplemented fructose in drinking water resulted in more prominent weight gain, insulin resistance and hepatic steatosis. |
| Bezan et al. [50] | Rats (Wistar) | M | 30 days | Evaluating the consequences of a high dose of conjugated linoleic acid in rats fed with a normal low fat or a high fat diet. 4 different group of mice were fed with either soybean oil (7% fat), mixture of soybean oil (4% fat) and conjugated linoleic acid (3% fat), lard (45% fat), mixture of lard and conjugated linoleic acid (3% fat). | It was observed that both conjugated linoleic acid group developed insulin resistance after 30 days with an increase in glucose in the fasting state and in an insulin tolerance test. The groups that received the conjugated linoleic acid had increased liver weight and percentage of saturated fatty acids in the liver and adipose tissue. It was concluded that high dose of linoleic acid mixture increases insulin resistance and exacerbates hepatic steatosis when combined with high fat diet. |
|---|---|---|---|---|---|
| Gondim et al. [63] | Rats (Wistar) | M | 16 weeks | Evaluating the effect of the consumption of different levels and sources of lipids on metabolic parameters of Wistar rats. Animals were fed with high-fat diet containing 20% of lard to cause metabolic obesity. Animals were then divided into groups and were fed diets with lipid concentration of 5% or 20% of either lard, soybean oil or fish oil. | It was confirmed that the groups that consumed fish oil showed less weight gain and lower serum levels of triacylglycerol, total cholesterol, aspartate aminotransferase activity, atherogenic index, less amount of fat in the carcass. It was concluded that fish oil intake was able to modulate positively the metabolic changes resulting from high fat diets |
| Tung et al. [160]. | Mice (ICR) | F | 12 weeks | Menopause is associated with changes in body composition (decline in lean body mass and an increase in total fat mass), leading to obesity, metabolic syndrome, nonalcoholic fatty liver disease and heart disease. This study investigate the effect of long-term feeding of edible oils (soybean oil (SO), tea seed oil (TO), and lard oil (LO)) on female ovariectomized (OVX) mice. | It was observed that the body weight and relative tissues of UFP (uterus fatty peripheral) and total fat (TF) were significantly decreased in the OVX+TO group compared with the OVX+SO and OVX+LO groups. Also, mice that were fed with HFD rich in TO showed decreased accumulation of liver lipid droplets and adipocyte sizes of UFP compared with those fed with HFD rich in SO or LO. This reveals that SO and LO induce obesity and increase accumulation of liver lipid droplets. |
| Gao et al. [161] | Mice (ICR) | M | 3 weeks | Evaluating the effects of common oils on carbon tetrachloride induced liver fibrosis. Healthy male mice were administered with carbon tetrachloride intraperitoneally at 2.5 ml/ kg twice a week and the mice were pre-treated with olive oil, soybean oil, corn oil or lard oil. | It was discovered that soybean oil or olive oil significantly reduced alanine aminotransferase and aspartate aminotransferase levels in the serum and malondialdehyde, hydroxyproline and triglycerides in the liver when compared with corn oil or lard oil. It was concluded that soybean oil but not corn oil or lard oil exerts protective effects against carbon tetrachloride-induced liver fibrosis in mice, probably because of its antioxidant activity. |
|---|---|---|---|---|---|
| Yan et al. [162] | Mice (C57BL/6) | M | 12 weeks | Assessing the effects of a mixture of lard and vegetable oil on lipid metabolism. Mice were divided into 5 groups and each group were fed with either lard, sunflower oil, soybean oil, lard blended with sunflower oil or lard blended with soybean oil. | It was observed that the final body weights of mice in the lard group were significantly higher. Body fat and volume of fat cell were significantly higher in the lard group compared to others. It was concluded that lard diet induced accumulation of body fat, live, serum lipids which can increase the risk of obesity. Differential oil and fats have an impact on differential aspects in mouse lipid metabolism. |
| Soybean oil vs coconut oil vs lard | |||||
| Bueno et al. [163] | Mice (C57BL/6) | M | 2 days (acute) 60 days (chronic) | Evaluation of the effects of four different high-fat diets (enriched with soybean oil, fish oil, coconut oil, or lard) on adiponectin gene expression and secretion by the white adipose tissue (WAT) of mice fed on a selected diet for either 2 (acute treatment) or 60 days (chronic treatment). 3T3-L1 adipocytes were treated for 48 h with six different fatty acids: palmitic, linoleic, eicosapentaenoic (EPA), docosahexaenoic (DHA), lauric or oleic acids. | It was reported that the same reduction in levels of adiponectin gene expression was observed in epididymal adipose tissue of animals chronically fed soybean oil and coconut diets and 3T3- L1 cells treated with palmitic, linoleic, EPA and DHA acids. It was also reported that the effects appear to be time dependent as it was only observed in chronically fed (60 days) soybean oil and coconut oil. It is postulated in this study that the downregulation of adiponectin expression by dietary enrichment with soybean oil or coconut oil may contribute to the development of obesity, insulin resistance and atherosclerosis. |
Table I: Reported effects of coconut oil, soybean oil and lard intake relating to the induction of obesity and other metabolic disorders. M-male, F-Female, T-Total, *LDL-c – Low-density lipoprotein cholesterol, *HDL-c – High-density lipoprotein cholesterol, *PUFA – Polyunsaturated fatty acid, *MCT – Medium chain triglycerides, *NASH – Nonalcoholic steatohepatitis, *NAFLD – Nonalcoholic fatty liver disease, *TC – Total cholesterol, *TG – Triglyceride.
Regarding coconut oil or isolated medium chain fatty acids (MCFA), studies have shown their beneficial effects on energy metabolism [22, 23, 24], fat oxidation [25, 26], food intake [27, 28], and others even showed no harmful effects on serum cardiometabolic risk markers [29, 30, 31, 32]. Contrarily, other studies have shown the negative effects on lipid metabolism [33]. Poppitt and colleagues together with Kovacs and colleagues suggested a negative effect on food intake and satiety [34, 35]. Eyres and colleague reported that the consumption of saturated fatty acids is positively linked with increased low-density lipoprotein (LDL) cholesterol and also the development of cardiovascular diseases [36].
Considering calorie intake, calorie intake plays a vital role in diet-induced-obesity (DIO). Dauqan et al. [37] investigated the effect of coconut oil on weight gain and antioxidant enzymes activity in rat liver. Spraque-Dawley rats were fed with either normal pellets (containing 4 kcal%) or normal pellets together with coconut oil (containing 15 kcal%) for 4 and 8 weeks. It was observed that there was no significant difference in the weight, and also there was no significant difference in the superoxidize dismutase (SOD) of the control group and the treated group. However, Deol and colleagues demonstrated that 40 kcal% total fat from coconut oil induce significant increase in weight gain and adiposity of C57BL/6 male mice [38]. In addition Gunasekeran et al. [39] demonstrated that 35 kcal% fat of coconut oil did not only induce overweight in CD1 female mice when expose for 6 weeks but also have an impact on the developing embryo. It was observed that coconut oil causes low body weight in the embryo and affect infant growth and appearance. In a randomized controlled trial, Valente et al. [40] investigated effects of coconut oil consumption on body weight, energy metabolism and cardiometabolic risk markers. In this study, 15 adult women were given 25mL of coconut oil daily for 1 week, it was observe that coconut oil induces obesity but did not change energy metabolism and cardiometabolic risk markers. However, Chinwong and colleagues performed a randomized crossover trial in young healthy adults by administering 30mL of coconut oil to participants daily for 24 weeks. It was reported that daily consumption of 30mL coconut oil induces obesity and also significantly increased high-density lipoprotein cholesterol [41].
Soybean oil Soybean oil is gotten from the seed of Glycine max and it is edible [42]. Up till the year 2020, soybean oil production globally amounts to 56.52 million metric tons and China is the leading soybean oil producing country (15.5 million metric tons), followed by United States (11.0 million metric tons), Argentina (8.7 million metric tons) and Brazil (8.4 million metric tons) [43]. It is commonly used in food processing, salad preparation, snacks preparation and it is the most commonly used oil in many restaurants and cafeteria [44].
Soybean oil contains more of polyunsaturated fatty acids as shown in Table 2 and Figure 1.
| Fatty acids (%) | CO* | SO* | Lard |
|---|---|---|---|
| Butyric C4:0 | - | - | - |
| Caproic C6:0 | 0.5 | - | - |
| Caprylic C8:0 | 7.8 | - | - |
| Capric C10:0 | 6.7 | - | - |
| Lauric C12:0 | 47.5 | - | 1.7 |
| Myristic C14:0 | 18.1 | - | - |
| Palmitic C16:0 | 8.8 | 10 | 26 |
| Stearic C18:0 | 2.6 | 4 | 16.5 |
| Palmitoleic C16:1 | - | - | 2.3 |
| Oleic C18:1 | 6.2 | 23 | 36.6 |
| Linolenic C18:2 | 1.6 | 51 | 11.3 |
| α-Linolenic C18:3 | - | 7 | 0.7 |
| Arachidic C20:0 | 0.1 | - | 0.2 |
| Behenic C22:0 | - | - | 0.2 |
| Gadoleic C20:1 | Traces | - | 0.8 |
| Others | - | 5 | 3.6 |
| Sterol composition | |||
| Cholesterol | 1.8 | 1 | |
| Brassicasterol | 0.45 | 0.15 | |
| Campesterol | 7.2 | 20 | |
| Stigmasterol | 10.5 | 17.5 | |
| β-sitosterol | 35.2 | 54.6 | |
| D5-Avenasterol | 27.8 | 2.8 | |
| D7-Stigmasterol | 1.5 | 3.3 |
Table 4: Fatty acids and components of coconut oil, soybean oil, lard and other major vegetable oil.
It is also a good source of compounds which includes sterols, flavonoids and tocopherols [45]. Soybean oil contains phytosterols such as brassicasterol, egrosterol, campesterol, campestenol, stigmasterol, β-sitosterol, cycloartanol, cycloartenol and methylene cycloartanol [46]. However, soybean oil is a rich source of omega-6 fatty acids with an omega-6/omega-3 ratio (8:1) which is not healthy. Simopoulos reported that increase in omega-6 and decrease in omega-3, such as the ratio we have in soybean oil is linked to overweight or obesity [47]. Soybean consumption increases as a result of the initial nutritional recommendation that people should limit the intake of saturated fat and increase their intake of diets rich in polyunsaturated fat [48, 49]. In fact, the United States recorded >1000% increase in the consumption of soybean oil from 0.01 to 11.6kg/year/capital between 1909-1999 [50]. Nevertheless, the recommendation is valid till today and this has led to daily increase in the consumption of soybean oil. In 2015, the Food and Drug Administration (FDA) ordered that trans fat were not safe to eat and gave the food-makers three years to get rid of trans fat from the food supply. However, in June 18, 2018, trans fat were totally eliminated and this also lead to increase in the consumption of soybean oil.
There has been little attention as regards soybean oil been linked to obesity because the assumption is that polyunsaturated fatty acids are good. As a result of this assumption, it has attracted fewer studies. The fewer studies available have a contrasting result in which some linked soybean oil to obesity and metabolic syndrome while others only linked it to only metabolic syndrome [51, 52]. Experimental studies in mice have shown an interesting result as to whether soybean oil induces obesity/diabetes or not [38, 51, 53, 54, 55, 56]. Lee and colleagues discovered that there was no significant difference in the body weight when female obese Zucker rats were fed with soybean oil (control) and a mixture of tricaprylin and soybean oil, this can be as a result of both having similar total fatty acids composition [57]. Sena and group linked soybean oil to diabetes [51]. They evaluated the effect of soybean oil on glycaemic control in Goto-Kakizaki male rats, an animal model of type-2 diabetes. It was discovered, after soybean oil treatment, higher α-tocopherol levels, suggesting a positive soybean oil effect on treating type-2 diabetes [51]. Soybean oil has also been proven to have high amount of coenzyme Q.
Considering calorie intake, Costa et al. [54] investigated intra-abdominal adiposity and insulin resistance in Wistar rats fed with either 7mL or 19mL of soybean oil immediately after weaning for 60 days. It was reported in this study that there is a differential effect as a result of the differences in amount of soybean oil used in the study. Though, both groups of rats had an increased body growth and fat depots but the group that received 19mL in addition had insulin resistance. However, Deol and colleagues reported that there was significant greater weight gain after soybean oil (40 kcal%) were given to C57BL/6N mice for 8 weeks when compared to the control high fat diet which comprised of coconut oil [58]. In addition, Bezan et al. [59] performed an experiment evaluating the differential effect of normal low-fat or high- fat diet. Male Wistar rats received either 7 kcal% (soybean oil) or 45 kcal% (lard) It was observed that after 30 days of exposure, the male mice on the low-fat diet (7 kcal% soybean oil) did not develop insulin resistance and also reduced weight when compared with the group that received 45% fat (lard).
It was discovered that soybean oil induced hyperglycemia and obesity in female mice after daily intake of soybean oil for 19 weeks [60]. In fact, it was reported in the experiment after comparing soybean oil with other oils, body weight changes induced by the different oil as follows: soybean oil > palm oil ≥ lard ≥ rapeseed oil ≥ safflower oil ≥ perilla ˃ fish oil. It was confirmed that high intake of soybean oil led to obesity and was also a risk factor for dysregulation of glucose tolerance [60]. Deol and colleagues carried out an experiment and compared soybean oil, coconut oil and fructose to evaluate which one of it is most obesogenic and diabetogenic [38]. In their study, the male mice were divided into groups and fed either with soybean oil, coconut oil or fructose. It was observed that the soybean oil fed group showed a significant increase in body weight, adiposity, diabetes, glucose intolerance and insulin resistance when compared to the group with coconut oil or fructose experimental groups. It was suggested that the intake of soybean oil was more obesogenic and diabetogenic than coconut oil and fructose and thus the consumption of soybean oil was more detrimental to metabolic health. Other studies have confirmed this results, it has been established that the intake of soybean oil is associated with obesity and fatty liver [38, 52, 55, 61, 62, 63].
It should be noted that high ratio of dietary n-3/n-6 polyunsaturated fatty acids improves obesity-linked inflammation and insulin resistance. Liu et al. [64] conducted an experiment comparing high n-3/n-6 PUFA ratio (1:1, PUFA) and low n-3/n-6 PUFA ratio (1:4, PUFA). In this study, Sprague-Dawley male rats were exposed for 16 weeks receiving diets containing either of the PUFA ratio. It was observed that PUFA (1:1) diet-fed rats had decreased body and visceral fat weight, improved glucose tolerance and were insulin sensitive when compared with the low n-3/n-6 PUFA ratio. It was reported that high n-3/n-6 PUFA (1:1) diet plays a role in preventing obesity and metabolic disorders by suppressing the activation of TLR4 [64]. However, the ratio omega-6 to omega-3 in soybean oil is 8:1, this shed more light on the link of soybean oil to obesity.
Lard Lard is a white fat product derived from the adipose tissue of pigs, which is different from tallow (similar product obtained from cattle or sheep). It is composed of the fatty acids, lauric acid, C-12:0 (1.7%); palmitic acid, C-16:0 (26%); stearic acid, C-18:0 (16.5%); arachidic acid, C-20:0 (0.2%); Behenic acid, C-22:0 (0.2%); palmitoleic acid, C-16:1 (2.30%); elaidic acid, C-18:1 (36.6%); linoelaidic acid, C-18:2 (11.3%); linoleic acid, C-18:3 (0.70%); gadoleic acid, C-20:1 (0.8%); and others (3.6%).
In 2018, global lard production was about 6.5 million tonnes which increase by 2%, when compared to the previous year [11]. China is the country with the largest production of lard (2.6 million tonnes), followed by Germany (653 thousand tonnes) and Brazil (481 thousand tonnes) [11]. As regards to the concentration of fatty acids, lard is rich in monounsaturated and saturated fat, but contains little of polyunsaturated fat, as shown in Figure 1.
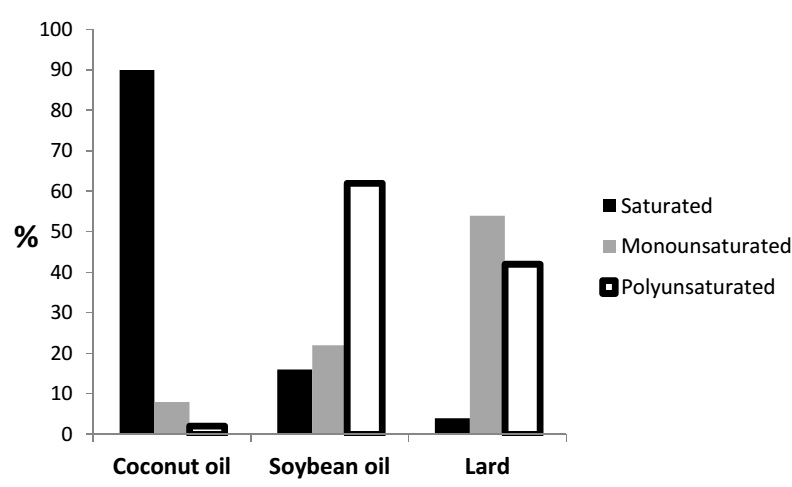
It is a popular ingredient in many kitchen and cafeteria, as cooking fat. It can also be used to produce biofuel and soap. Lard, together with other fat from animals, was initially used in industrial fermentation processes (brewing) as anti- foaming agents [65]. In China, it is one of the most common cooking-oils, and great effects on diuresis, blood clearance, blood circulation activation and detoxification have already been reported [66]. Studies have shown that high lard intakes were linked to increase in adiposity, insulin resistance which in turn led to the development of obesity [66, 67, 68, 69]. As a result of its high contents of monounsaturated and saturated fatty acids, there have been debates over the consumption of lard, considering the beneficial and the negative effects on biological systems. There is evidence that high lard intake was linked to increase in adiposity, insulin resistance which in turn leads to the development of obesity. Kubeck et al. [69] discovered that the intake of cholesterol-rich lard- based high-fat diet together with its interaction with gut microbiota induced obesity in male mice. It was suggested in this study that dietary cholesterol is a candidate ingredient that alters the crosstalk between gut microbiota and host metabolism. Wang et al. [66] performed an experiment to evaluate the differential effects of lard and soybean oil on osteopontin expression and inflammation of adipose tissue in diet-induced obese male rats. From their results, it was suggested that the replacement of lard by soybean oil in diets could reduce obesity-related inflammation and insulin resistance by decreasing the upregulation of osteopontin and macrophage infiltration, caused by high fat diets. Similarly, Matias et al. [70] also performed an experiment evaluating the differential effects of high sugar and high lard and the combination of both on nutritional, hormonal and cardiovascular parameters in rodents. From their results, it was reported that lard and the combination of both lard and sugar diet induced obesity. Different studies have also confirmed that lard promotes obesity by increasing the body weight and adipose tissue and also causing glucose metabolism changes and metabolic syndrome [71, 72, 73].
Considering calorie intake, Shao et al. [74] conducted an experiment in which male Sprague-Dawley rats were fed with either normal diet (100% standard rodent chow (3.49 kcal/g)) or high-fat lard diet (85% standard rodent chow supplement with 15% Lard (4.14 kcal/g)) for 6 weeks. It was reported that dietary high fat lard intake induces abnormal morphology and thyroid dysfunction in rats. Gondim and colleagues evaluated the effect of the consumption of lards on metabolic parameters in Wistar rats. In this study, Wistar rats were fed with either high-fat diet containing 20% of Lard or the control diet for 12 weeks. It was observed that daily lard consumption induces metabolic obesity [63]. In addition, Emelyanova et al. [75] conducted an experiment to determine the impact of high calories and fat content in diet on obesity, cardiac structure. In this study, C57BL/6J male mice were fed with high calorie, high lard fat-based diet (60% fat) or low calorie and fat diet (12% fat) for 10 weeks. It was observed that the group of mice fed with high lard fat-based diet had higher body weight, ventricular mass and thickness of posterior and septal wall when compared to the low-calorie and fat diet fed mice. It was observed that the higher body weight was as a result of excessive calories consumed.
Intracellular Metabolism and Biochemistry
Soybean oil is a rich source of omega-6 fatty acid with an unbalanced omega-6/omega-3 ratio (8:1), it has been reported that this unbalance ratio that favor omega-6 is highly prothrombotic and proinflammatory which revealed its linked to obesity, diabetes and atherosclerosis [76, 77]. Omega-6 fatty acids are the linoleic acid (LA, 18:2ω-6) while omega-3 fatty acids are the α-linolenic acid (ALA, 18:3ω-3). The difference between the two is based on the location of the first double bond. The metabolism of LA leads to arachidonic acid (AA, 20:4ω6) while ALA metabolized to eicosapentaenoic acid (EPA, 20:5ω3) and docosahexaenoic acid (DHA, 22:6ω3). In humans, haplotype D aids the production of AA and EPA from LA and ALA respectively. Fatty acid desaturase 1 and 2 (FADS1 and FADS2) also helps in fatty acid metabolism (47).In mammalian cells, Omega-6 fatty acids cannot be converted to omega-3 fatty acids as a result of lack of omega-3 desaturase (converting enzyme). Omega-3 and omge-6 fatty acids are metabolically and functionally different and hence their balance is very important in diet. In soybean oil, there is an imbalance in the ratio of omega-6 and omega-3 (8:1) and this affect the biosynthesis and increase the risk of diseases of inflammation such as obesity and cancer. The absorption of lipids has some interesting hallmark. First and foremost, digestion of lipids occurs in the lumen before it enters the enterocytes. Secondly, many of the lipids congregate into lipoproteins which are primarily called chylomicrons [78]. The chylomicrons are secreted into intercellular space, diffused into the lamina propria, moved to the lacteals and finally reach the lymphatic system. However, the intracellular metabolism of fatty acids occurs in either of the two pathways which are the monoacylglycerols (MAGs) pathway or the portal route. In the MAGs pathway, the long chain fatty acids (LCFAs) (≥ 16 carbons) get as far as the endoplasmic reticulum membrane where they are re-synthesized into triacylglycerols (TAGs) [79]. In contrast, the short chain fatty acids (SCFAs) and medium chain fatty acid (MCFAs) (6-12 carbons) avoid the MAG pathway, being transported to the liver for rapid oxidation through the portal circulation [80].
Our focus in this section is the intracellular metabolism of coconut oil, soybean oil and lard. Coconut oil is a good source of MCFAs and it follows the portal route. However, soybean oil and lard are quality source of long chain fatty acids (LCFAs) which implies that they both follow the MAG pathway. In the MAG route, the MAGs are esterified with fatty acyl-CoA in a step by step process to form diacylglycerols (DAGs), this process is catalyzed by monoacylglycerol acyltransferases (MGATs). TAGs are also formed from DAGs and this process is catalyzed by diacylglycerol acyltransferases (DGATs).
MGAT are highly expressed in the small intestine and are classified in the following subtypes; MGAT1, MGAT2. The MGAT2 subtypes are present in the intestine of mice and human while MGAT3 are only present in the intestine of humans [81, 82]. MGAT2 plays a critical role in the esterification of MAGs. Nelson and colleagues performed a study involving intestinespecific MGAT2null mice, and reported reduced uptake and esterification of MAGs and delayed fat absorption [83]. DGAT enzymes help in converting DAGs to TAGs. There are majorly two types which are DGAT1 and DGAT2. DGAT2 has been proven to be more important than DGAT1. As reported by Smith et al. [84] and also Chen et al. [85], Dgat1-knockout mice have no deficiency in fat absorption. However, report from Stone et al. [86] showed that Dgat2knockout mice reveal total absence of fat and die shortly after birth. Moreover, MGAT2 and DGAT1 have shown protective effects against diet-induced obesity and related metabolic conditions. Some drugs such as cholestyramine, orlistat, lomitapide, PF-04620110, JTP- 103237 have been identified as inhibitors of lipids digestion and absorption because they act as lipase, MTP, DGAT and MGAT inhibitors (Table 3).
| Publication | Drug Class (Examples) | Mechanism | Report |
|---|---|---|---|
| Hofmann et al. [167] | Bile acid sequestrant (cholestyramine) | reduce bile acid pool size and increase hepatic bile acid synthesis from cholesterol | Approved as LDL lowering therapy in humans |
| Heck,et al. [168] | Lipase inhibitor (orlistat) | Inhibits gastric and pancreatic lipases to impair TAG Digestion | Approved for obesity treatment in humans |
| Rizzo et al. [169] | MTP inhibitor (lomitapide) | Inhibits MTP large subunit to impair lipid transfer for chylomicron assembly and lipidation | Lomitapide approved as LDL lowering therapy in homozygous familial hypercholesterolaemia in humans |
| Dow et al. [170] | DGAT1 inhibitor (PF- 04620110) | Inhibits enterocyte DGAT1 to impair TAG re-esterification and absorption | Effective in treating obesity induced through diet and associated diabetes. |
| Okuma et al. [171] | MGAT2 inhibitor (JTP- 103237) | Inhibits enterocyte MGAT2 to impair TAG re-esterification and absorption | Effective in treating obesity induced through the diet and associated diabetes. |
Table III: Some inhibitors of lipids digestion and absorption.
Contrarily, in case of coconut oil, because they are good quality source of MCFAs, they bypass the MAG pathway and the fatty acids are conveyed to the liver from the portal blood. They are transported with albumin in the blood directly. In addition, one of the merits of this type of fat is that it has the ability of being absorbed quantitatively in the intestinal lumen with reduction in lipase activity. They do not go through processes like degradation and re-esterification [87] and this make them unique compared to the long chain fatty acids. It has been said that coconut oil has some thermogenic effects and also help in high density lipoprotein cholesterol (HDL-c) improvement [88, 89]. This report is based on the fact that coconut oil can undergo faster absorption and can be metabolized quickly, compared to other fatty acids [42]. The speed of absorption and metabolism are influenced because it is partly independent of the cartinine transport mechanism and it is rapidly oxidized for energy production [90].
Changes in Gene Expression
There have been reported cases of changes in gene associated with obesity and also dysregulation in the genes associated with cancers after coconut oil, soybean oil and lard consumption. Gene that has been dysregulated after the consumption of coconut oil is the HMGCR (3-hydroxy- 3-methylglutaryl-CoA reductase). HMGCR bring about the biosynthesis of cholesterol which has been reported to reduce the risk of cardiovascular disease [91, 92]. Study carried out by Liu et al. [93] showed dysregulation in HMGCR gene after the intake of coconut oil which alters the statin mechanism and tends to have a negative effect. Cyp7a1 (Cytochrome P450 Family 1 Subfamily A Member 1) play a significant role in the degradation of cholesterol to bile ducts. Dysregulation of this gene resulted in hypercholesterolemia, leading to cardiovascular disorder [94].
The fatty acid translocase Cd36 helps in the absorption of free fatty acids, playing a vital role in hepatic steatosis [95]. Moreover, fatty acid binding protein Fabp4, which links obesity induced by diet to insulin resistance, plays a significant role in maintaining hepatic metabolic balance [96, 97]. Deol et al. [38] described an increase in the level Cd36 and Fabp4 after the intake of soybean oil in a group of mice. Fatty acid binding protein 5, Fabp5, helps in the prevention of lipid dysregulation and also helps in detoxifying free fatty acids [98]. It was observed in the experiment carried out by Deol et al. [38] that there was a three-fold decrease in this gene when compared to the control group. Studies also showed an increase in Igfbp1, a regulator of Insulin-like growth factor 1 (IGF1) after the intake of soybean oil by mice. Ooi et al. [99] has earlier reported that increase in the hepatic expression of Igfp1 is linked to diabetes. There is evidence that soybean oil causes dysregulation in hepatic cytochrome P450 (Cyp) genes, causing a decrease of Cyp7b1 expression in mice after the intake of soybean oil [38, 100], which helps in converting cholesterol to bile acids [101].
According to Deol et al. [38], about 31 genes attributed to inflammation were dysregulated after mice were fed with either soybean oil or coconut oil. Cancer-promoting genes that were upregulated included Ctgf, H19, Mmp12, Mybl1, Vnn1, whereas cancer-inhibiting genes Cish, Dkk4, Onecut1, Scara5, Socs3, Wif1 were also suppressed. The dysregulation of cancer genes after the intake of soybean oil suggested that not only high consumption of soybean oil can cause obesity, but it can also increase the susceptibility to liver cancer. In addition, just like obesity, the prevalence of liver cancer has been recently increased globally [102, 103]. In this study, an increase in the expression of Acot1 and Acot2 genes was observed after the intake of coconut oil. Elevation in the expression of these genes can lead to accumulation of hepatic lipids. In the study, cell death activator Cidea, Cidec (CIDE family member) and Plin4 (PAT family member) were also up-regulated after the intake of coconut oil and these three genes have been linked to obesity [104].
Skrivan et al. [105] performed an experiment to assess the effect of different dietary fat types on intestinal digestibility of fatty acids and expression of lipid-related genes in broiler chickens. It was observed that there was an increase in the expression of fatty acid synthase in chicken fed palm oil when compared to chicken fed lard. There was also an increase in the expression of HMG-CoA reductase in the group fed palm oil compared to the group fed lard. The reduction in the expression of HMG-CoA reductase in the group fed lard revealed lard association with obesity and adiposity.
Sex differences in Obesity
Sex refers to the biological characteristics of the result of XX (female) or XY (male) sex chromosomes, which influence gonadal hormones levels and types. Sex differences play a role in the initiation and development of different diseases. Women are known to suffer more autoimmune diseases than men (such as scleroderma, Sjogren’s depression) [106, 107, 108, 109, 110, 111]. Also, autism occurs more in boys compare to girls [112]. However, in obesity, sex differences play a vital role.
Initially, it was believed that male mice had greater fat mass expansion than female in the visceral and subcutaneous region; this was believed to be as a result of sex hormones [113, 114, 115]. Recently, Salinero et al. [116] demonstrated that the sex differences in diet-induced obesity after the consumption of high-fat diet is much more dependent on the age of the mice. Feeding six-week old mice for 3 months with high-fat diet led to a higher increase in the body weight of the male compared to the female. However, in the adult mice (31 weeks), females presented higher body weight when compared to male, after the intake of high-fat diet. Another important issue that may contribute to sex differences in adipose tissue expansion is the number of pluripotent stem cells (adipocyte precursor cells) in the mouse gonadal or subcutaneous fat regions. As reported by Joe and colleagues (2009) [117], female mice have more adipocyte precursor cells than their male counterparts in gonadal (visceral) and inguinal (subcutaneous) fat depots. This result was also confirmed in other experiments by different authors [118, 119]. Wu et al. [119] reported that when mice was fed with high fat diet (45% calories as fat), there was an increase in the adipocyte precursor cells and mature adipocytes in the female mice, whereas the male mice did not show any increase of the mature fat cells in the gonadal fat depots.
National Health and Nutrition Examination Survey (NHANES) from 2013–2014 in US estimated an increase prevalence of obesity in women than in men (40.5 vs. 35.2%) [120]. In fact, male and female have differences in the adipose tissue distribution, such as while men amass large amount of visceral adipose tissue, women amass fat accumulation in the sub-cutaneous depots [121, 122]. Diet and hormonal status influences sex-specific fat distribution [123]. In addition, some studies also assessed the lineage tracing to trace the fate of proliferating adipocyte progenitor cells. The results from these studies showed that male gonadal fat revealed enlargement, whereas female showed adipocyte hyperplasia in both the gonadal and the subcutaneous fat regions after the intake of high-fat diets [124, 125]. Another important factor that influences obesity and adiposity are the gonadal hormones. The contribution of the gonadal hormones in obesity in women can be explained by comparing the women’s body condition before and after menopause: the reduced levels of ovarian hormones after menopause caused an increase in body condition. When comparing pre-menopausal and post-menopausal women, the effect of gonadal hormones on fat storage becomes very conspicuous. Immediately after menopause, the level of estrogen and other gonadal hormones reduces, promoting a natural increase in fat storage of the abdominal depots, tending to further increase the rate of cardiovascular diseases and insulin resistance [126, 127, 128, 129]. On the other hand, androgen levels have impact on metabolic diseases. Testosterone levels in men reduce with age, and low testosterone levels in men are linked to increased body fat and cardiovascular disease [130, 131, 132]. As reported by Santosa et al. [133], when estrogen was reduced (4 weeks, short term), there was no alteration in the fat storage but post-prandial circulating triglyceride level was affected. Interestingly, the suppression of testosterone level in men (for 4 weeks) led to the increase storage of fatty acids in the gluteal-femoral fat region [134]. Overall, hormone levels, estrogen and androgen receptors may likely influence gonadal hormone on fat region development and metabolism.
Public Health Policies
The prevalence of obesity has increased globally to the extent that nearly one third of the world population is now classified as being overweight or obese [135]. Obesity affects physiological functions of the body and has now been considered a public health threat. It also affects all ages and can occur in both sexes regardless of geographical locality, ethnicity or socioeconomical status. However, the prevalence of obesity is generally greater in elderly people and women [136]. Non-communicable diseases (NCD) Risk Factor Collaboration reported that by 2025, the prevalence of obesity will be 18% in men and 21% in women [137]. This high prevalence of obesity globally has been linked to dietary fat and it has led to different nutritional guidelines across the globe.
The increase rate in the consumption of coconut oil and soybean oil might be linked to the increase rate of obesity in the United States of America. In United State of America, it was estimated that a single individual with obesity incurred health cost of about US$ 1,901 per annum in 2014, amounting to US$ 149.4 billion, considering the total American obese population [138]. Given this scenario, the US Department of Agriculture [139] and the World Health Organization [140] recommended that the daily intake of saturated fat should not exceed 10% of the total calories.
As reported by Chooi et al. [141], China witnessed about 9-fold increase in the prevalence of obesity between 0.6% in 1980 and 5.3% in 2016. The high consumption of coconut oil, soybean oil and lard can be linked to this increase prevalence of obesity in China. As a result of this increase rate in obesity, the Chinese Nutrition Society recommended a new dietary guideline for Chinese residents in the form of Chinese Food Pagoda in 2016 [142]. It was recommended that the daily intake of cooking oil in adult should not exceed 25 – 30g. They recommended that for trans fatty acids, the daily intake should be less than 2g.
In Europe, the increase in the consumption of coconut oil, soybean oil and lard is alarming. Likewise, the prevalence of obesity in the region is also at increase rate. In Europe, total (direct and indirect) cost attributed to obesity was estimated to be 0.47-0.61% of the Gross Domestic Product (GDP) [143]. Though, the European Food Safety Authority (EFSA) did not specify the limit but it recommended that the consumption of saturated fat should be as low as possible [144]. Some arguments have come from the angle that coconut oil is a saturated fat, but it is composed of majorly medium chain fatty acids (MCFAs) about 60% and MCFAs helps in the reduction of weight, the EFSA. (2011) [145] and the American Heart Association (AHA) [146] team draw the inference that there is not enough evidence in human intervention studies to buttress the claim that MCFAs helps in weight management. In addition, the European Union Commission Regulation recommended that the content of free fatty acids present in the daily intake of soybean oil must not exceed 1.5% [147].
In Brazil, the Brazilian Society of Endocrinology and Metabolism (SBEM) and the Brazilian Association for the Study of Obesity and Metabolic Syndrome (ABESO) have also released a warning on the use of MCFAs as weight reducing agents. Both scientific agencies affirmed that there is not enough scientific proof to support this claim and so, the use of coconut oil should be restricted [148]. Additionally, the National Health Surveillance Agency [149] recommended that the content of free fatty acids present in the daily intake of soybean oil must not exceed 0.9%. However, the interpretation of nutrient-based recommendations to the general public has been an issue, because only few people accurately estimate, without guidance, their daily consumption of nutrients such as calories, fats, cholesterol, fiber, salt, or single vitamins. Hence, considering the public health threat that obesity is causing and its link to dietary fat, we strongly recommend that people should limit the intake of coconut oil, soybean oil and lard.
Conclusion
Coconut oil has a negative effect on lipid metabolism, it is positively linked with increased low-density-lipoprotein. Lard increase adiposity and insulin resistance which in turn lead to obesity. Although, coconut oil and lard induced obesity, studies have shown that soybean oil is more obesogenic than both and increased caloric intake causes adiposity, diabetes, glucose intolerance and insulin resistance. High intake of these dietary fats also leads to changes in the expression of genes linked to obesity and inflammation. Obesity is now a global threat affecting developed and developing countries and high intake of coconut oil, soybean oil and lard has been strongly linked to it. We, therefore, recommend that people should limit the intake of these dietary fats and oils. As regards coconut oil, the daily intake must not exceed 10% of the total calories; as regards soybean oil, the free fatty acids present in daily intake must not exceed 1.5%; while as for lard, the daily intake must not exceed 10% of total calories.
Future research studies that consider the components sex-biasing effects which include not only the gonad type but also the sex chromosome should be encouraged; as such research line is very important to ascertain the adequate nutritional recommendation to both male and female. This will certainly give an insight to the relationship between gender and obesity before, during and after the consumption of dietary fats.
References
-
Finucane MM, Stevens GA, Cowan MJ, Danaei G, Lin JK, et al. (2011) National, regional, and global trends in body- mass index since 1980, systematic analysis of health examination surveys and epidemiological studies with 960 country years and 9.1 million participants. Lancet 377(9765): 557-567.
-
World Health Organization (2017) Obesity and overweight. Geneva: WHO.
-
Ng M, Fleming T, Robinson M, Thomson B, Graetz N, et al. (2014) Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet 384(9945): 766–781.
-
Keys A (1957) Diet and the epidemiology of coronary heart disease. Journal of the American Medical Association 164(17): 1912–1919.
-
Kimura N, Keys A (1970) Coronary heart disease in seven countries. X. Rural southern Japan. Circulation 41: 1101–1112.
-
Eckel RH, Jakicic JM, Ard JD, de Jesus JM, Houston MN, et al. (2014) 2013 AHA/ACC guideline on lifestyle management to reduce cardiovascular risk: a report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. Circulation 129 (25 Suppl 2): S76-S99.
-
Chowdhury R, Warnakula S, Kunutsor S, Crowe F, Ward HA, et al. (2014) Association of dietary, circulating, and supplement fatty acids with coronary risk: a systematic review and meta-analysis. Annuals of Internal Medicine 160(6): 398–406.
-
Poudyal H, Brown L (2015) Should the pharmacological actions of dietary fatty acids in cardiometabolic disorders be classified based on biological or chemical function? Prog Lipid Res 59: 172-200.
-
Aued-Pimentel S, Castro FD, Sousa RJ, Mello MRA, Abe- Matsumoto LT (2015) Commercialised coconut oil in Sao Paulo city, Brazil: evaluation of authenticity and nutritional labeling. Journal of Agriculture and Life Sciences 2(1): 76-83.
-
Statista (2020a) Coconut oil consumption in the United States from 2000-2019.
-
(2019) Global Trade.
-
DebMandal M, Mandal S (2011) Coconut (_Cocos_ _nucifera L_.: arecaceae): in health promotion and disease prevention. Asian Pacific Journal of Tropical Medicine 4(3): 241–247.
-
Tridge (2020) Top producing countries of coconut (in million metric tons).
-
Mosley M (2018) Is coconut oil a superfood? BBC News.
-
Mustapha K (2018) The coconut oil debate. New Straits Times.
-
Noone Y (2018) Why Dr Michael Mosley now thinks that coconut oil may be good for you. SBS.
-
(2018) Piping Rock.
-
Dumolt JH, Rideout TC (2017) The lipid-lowering effects and associated mechanisms of dietary phytosterol supplementation. Curr Pharm Des 23(34): 5077-5085.
-
Martins CM, Fonseca FA, Ballus CA, Figueiredo-Neto AM, Meinhart AD, et al. (2013) Common sources and composition of phytosterols and their estimated intake by the population in the city of São Paulo, Brazil. Nutrition 29(6): 865-871.
-
Nevin KG, Rajamohan T (2008) Influence of virgin coconut oil on blood coagulation factors, lipid levels and LDL oxidation in cholesterol fed Sprague-Dawley rats. European e-Journal of Clinical Nutrition and Metabolism 3**:** e1–e8.
-
Nevin KG, Rajamohan T (2004) Beneficial effects of virgin coconut oil on lipid parameters and in vitro LDL oxidation. Clin Biochem 37(9): 830–835.
-
Bendixen H, Flint A, Raben A, Hoy C, Mu H, et al. (2002) Effect of 3 modified fats and a conventional fat on appetite, energy intake, energy expenditure, and substrate oxidation in healthy men. Am J Clin Nutr 75(1): 47–56.
-
White MD, Papamandjaris AA, Jones PJ (1999) Enhanced postprandial energy expenditure with medium- chain fatty acid feeding is attenuated after 14 d in premenopausal women. Am J Clin Nutr 69(5): 883–889.
-
Alexandrou E, Herzberg GR, White MD (2007) High- level medium-chain triglyceride feeding and energy expenditure in normal-weight women. Can J Physiol Pharmacol 85(5): 507–513.
-
Van Wymelbeke V, Himaya A, Louis-Sylvestre J, Fantino M (1998) Influence of medium-chain and long-chain triacylglycerols on the control of food intake in men. Am J Clin Nutr 68(2): 226–234.
-
Van Wymelbeke V, Louis-Sylvestre J, Fantino M (2001) Substrate oxidation and control of food intake in men after a fat substitute meal compared with meals supplemented with an isoenergetic load of carbohydrate, long-chain triacylglycerols, or medium-chain triacylglycerols. Am J Clin Nutr 74(5): 620–630.
-
Nosaka N, Kasai M, Nakamura M, Takahashi I, Itakura M, et al. (2002) Effects of dietary medium-chain triacylglycerols on serum lipoproteins and biochemical parameters in healthy men. Biosci Biotechnol Biochem 66(8): 1713–1718.
-
Assunção ML, Ferreira HS, dos Santos AF, Cabral C, Florencio T (2009) Effects of dietary coconut oil on the biochemical and anthropometric profiles of women presenting abdominal obesity. Lipids 44(7): 593–601.
-
Liau KM, Lee YY, Chen CK, Rasool AHG (2011) An open- label pilot study to assess the efficacy and safety of virgin coconut oil in reducing visceral adiposity. ISRN Pharmacol 2011: 949686.
-
Vijayakumar M, Vasudevan DM, Sundaram KR, Krishnan S, Vaidyanathan K, et al. (2016) A randomized study of coconut oil versus sunflower oil on cardiovascular risk factors in patients with stable coronary heart disease. Indian Heart J 68(4): 498–506.
-
Tholstrup T, Ehnholm C, Jauhiainen M, Petersen M, Hoy C, et al. (2004) Effects of medium-chain fatty acids and oleic acid on blood lipids, lipoproteins, glucose, insulin, and lipid transfer protein activities. Am J Clin Nutr 79(4): 564–569.
-
Poppitt SD, Strik CM, MacGibbon AKH, McArdle BH, Budgett S, et al. (2010) Fatty acid chain length, postprandial satiety and food intake in lean men. Physiol Behav 101(1): 161–167.
-
Kovacs EM, Westerterp-Plantenga MS, de Vries M, Brouns FJPH, Saris WHM (2001) Effects of 2-week ingestion of (-)-hydroxycitrate and (-)-hydroxycitrate combined with medium-chain triglycerides on satiety and food intake. Physiol Behav 74(4-5): 543–549.
-
Eyres L, Eyres MF, Chisholm A, Brown RC (2016) Coconut oil consumption and cardiovascular risk factors in humans. Nutr Rev 74(4): 267–280.
-
Dauqan E, Sani HA, Abdullah A, Kasim ZM (2011) Effect of different vegetable oils (red palm olein, palm olein, corn oil and coconut oil) on lipid profile in rat. Food Nutrition Sciences 2(4): 253–258.
-
Deol P, Evans JR, Dhahbi J, Chellappa K, Han DS, et al. (2015) Soybean oil is more obesogenic and diabetogenic than coconut oil and fructose in mouse: potential role for the liver. PLoS One 10(7): e0132672.
-
Gunasekaran R, Shaker MR, Mohd-Zin SW, Abdullah A, Ahmad-Annuar A, et al. (2017) Maternal intake of dietary virgin coconut oil modifies essential fatty acids and causes low body weight and spiky fur in mice. BMC Complement Altern Med 17(1): 79.
-
Valente FX, Candido FG, Lopes LL, Dias DM, Calvalho SDL, et al. (2018) Effects of coconut oil consumption on energy metabolism, cardiometabolic risk markers, and appetitive responses in women with excess body fat. Eur J Nutr 57(4): 1627–1637.
-
Chinwong S, Chinwong D, Mangklabruks A (2017) Daily consumption of virgin coconut oil increases high-density lipoprotein cholesterol levels in healthy volunteers: A randomized crossover trial. Evid Based Complement Alternat Med 2017: 7251562.
-
Fernandes JCB, Draghi PF (2016) Thermal stability of soybean oil: when must we discard it? MOJ Food Processing and Technology 2(5): 170-174.
-
Statista (2020b)
-
Roccisano D, Henneberg M (2012) Soy consumption and obesity. Food and Nutrition Science 3(2): 260–266.
-
Navas PB, Fregapane G, Salvador A (2013) Quality indexes, mayor and minor constituents and oxidative stability of sesame and soybean virgin oils. Revista de la Facultad de Agronomia (LUZ) 30: 284–303
-
Yang R, Xue L, Zhang L, Wang X, Qi X, et al. (2019) Phytosterol Contents of Edible Oils and Their Contributions to Estimated Phytosterol Intake in the Chinese Diet. Foods 8(8): 334.
-
Simopoulos AP (2016) An increase in the omega-6/ omega-3 fatty acid ratio increases the risk for obesity. Nutrients 8(3): 128.
-
Page AEB, Chamberlain FL, Keys A, Stamler J, Stare FJ (1961) Dietary fat and its relation to heart attacks and strokes. Circulation 23: 133–136.
-
Kritchevsky D (1998). History of recommendations to the public about dietary fat. J Nutr 128(2 Suppl): 449S–452S.
-
Blasbalg TL, Hibbeln JR, Ramsden CE, Majchrzak SF, Rawlings RR (2011) Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am J Clin Nutr 93(5): 950–962.
-
Sena CM, Proenca T, Nunes E, Santos MS, Seica RM (2008) The effect of soybean oil on glycaemic control in Goto-kakizaki rats, an animal model of type 2 diabetes. Medicinal Chemistry 4(3): 293–297.
-
Alvheim AR, Torstensen BE, Lin YH, Lillefosse HH, Lock EJ, et al. (2013) Dietary linoleic acid elevates endogenous 2-arachidonoylglycerol and anandamide in Atlantic salmon (_Salmo salar L_.) and mice, and induces weight gain and inflammation in mice. Br J Nutr 109(8): 1508–1517.
-
Zhang XH, Hua JZ, Wang SR, Sun CH (2007) Post-weaning isocaloric hyper-soybean oil versus a hypercarbohydrate diet reduces obesity in adult rats induced by a high-fat diet. Asia Pac J Clin Nutr 16: 368–373.
-
Costa CA, Carlos AS, dos Santos AS, Monteiro AMV, de Moura EG, et al. (2011) Abdominal adiposity, insulin and bone quality in young male rats fed a high-fat diet containing soybean or canola oil. Clinics (Sao Paulo) 66(10): 1811–1816.
-
Midtbo LK, Ibrahim MM, Myrmel LS, Aune UL, Alvheim AR, et al. (2013) Intake of farmed Atlantic salmon fed soybean oil increases insulin resistance and hepatic lipid accumulation in mice. PLoS One 8(1): e53094.
-
da Costa, CAS, Carlos AS, dos Santos AS, de Moura EG, Nascimento-Saba CC (2013) High-fat diets containing soybean or canola oil affect differently pancreas function of young male rats. Horm Metab Res 45(9): 652–654.
-
Lee KT, Akoh CC, Flatt WP, Lee JH (2000) Nutritional effects of enzymatically modified soybean oil with caprylic acid versus physical mixture analogue in obese Zucker rats. J Agric Food Chem 48(11): 5696–5701.
-
Deol P, Fahrmann J, Yang J, Evans JR, Rizo A, et al. (2017) Omega-6 and omega-3 oxylipins are implicated in soybean oil-induced obesity in mice. Sci Rep 7(1): 12488.
-
Bezan PN, Holland H, de Castro GS, Cardoso JFR, Ovidio PP, et al. (2018) High dose of a conjugated linoleic acid mixture increases insulin resistance in rats fed either a low fat or a high fat diet. Exp Clin Endocrinol Diabetes 126(6): 379-386.
-
Ikemoto S, Takahashi M, Tsunoda N, Maruyama K, Itakura H, et al. (1996) High-fat diet-induced hyperglycemia and obesity in mice: differential effects of dietary oils. Metabolism 45(12): 1539–1546.
-
Alvheim AR, Malde MK, Osei-Hyiaman D, Lin YH, Pawlosky RJ, et al. (2012) Dietary linoleic acid elevates endogenous 2-AG and Anandamide and induces obesity. Obesity (Silver Spring) 20(10): 1984–1994.
-
Henkel J, Coleman CD, Schraplau A, Johrens K, Weber D, et al. (2017) Induction of steatohepatitis (NASH) with insulin resistance in wild-type B6 mice by western-type diet containing soybean oil and cholesterol. Mol Med 23: 70-82.
-
Gondim PN, Rosa PV, Okamura D, Silva VDO, Andrade EF, et al. (2018) Benefits of fish oil consumption over other sources of lipids on metabolic parameters in obese rats. Nutrients 10(1): 65.
-
Liu H-Q, Qui Y, Mu Y, Zhang X-J, Liu L, et al. (2013) A high ratio of dietary n-3/n-6 polyunsaturated fatty acids improves obesity-linked inflammation and insulin resistance through suppressing activation of TLR4 in SD rats. Nutr Res 33(10): 849-858.
-
Hill RM, Christiano SP (1996) Polymeric Materials Encyclopedia, CRC Press.
-
Wang X, Cheng M, Zhao M, Ge A, Guo F, et al. (2013) Differential effects of high-fat-diet rich in lard oil or soybean oil on osteopontin expression and inflammation of adipose tissue in diet-induced obese rats. Eur J Nutr 52(3): 1181-1189.
-
Simopoulos AP (1994) Is insulin resistance influenced by dietary linoleic acid and trans fatty acids? Free Radic Biol Med 17(4): 367–372.
-
Almeida FRCL, Machado GS, Borges ALCC, Rosa BO, Sources DO (2014) Consequences of different dietary energy sources during follicular development on subsequent fertility of cyclic gilts. Animal_._ 8(2): 293-299.
-
Kubeck R, Bonet-Ripoll C, Hoffmann C, Walker A, Muller VM, et al. (2016) Dietary fat and gut microbiota interactions determine diet-induced obesity in mice. Mol Metabol 5(12): 1162-1174.
-
Matias AM, Estevam WM, Coelho PM, Haese D, Kobi JBBS, et al. (2018) Differential effects of high sugar, high lard or a combination of both on nutritional, hormonal and cardiovascular metabolic profiles of rodents. Nutrients 10(8): 1071.
-
Buettner R, Schölmerich J, Bollheimer LC (2007) High- fat diets: Modeling the metabolic disorders of human obesity in rodents. Obesity 15(4): 798–808.
-
Hariri N, Thibault L (2010) High-fat diet-induced obesity in animal models. Nutr Res Rev 23(2): 270–299.
-
Arias-Mutis OJ, Marrachelli VG, Ruiz-Saurí A, Alberola A, Morales JM, et al. (2017) Development and characterization of an experimental model of diet- induced metabolic syndrome in rabbit. PLoS One 12(5): e0178315.
-
Shao S, Zhao Y, Song Y, Xu C, Yang J, et al. (2014) Dietary high-fat intake induces thyroid dysfunction and abnormal morphology in rats. Acta Pharnacologica Sinica 35(11): 1411-1420.
-
Emelyanova L, Boukatina A, Myers C, Oyarzo J, Lustgarten J, et al. (2019) High calories but not fat content of lard-based diet contribute to impaired mitochondrial oxidative phosphorylation in C57BL/6J mice heart. PLos One 14(7): e0217045.
-
Simopoulos AP (2013) Dietary Omega-3 Fatty Acid Deficiency and High Fructose Intake in the Development of Metabolic Syndrome, Brain Metabolic Abnormalities, and Non-Alcoholic Fatty Liver Disease. Nutrients 5(8): 2901-2923.
-
Donahue SM, Rifas-Shiman SL, Gold DR, Jouni ZE, Gillman MW, et al. (2011) Prenatal fatty acid status and child adiposity at age 3 years: Results from a US pregnancy cohort. Am J Clin Nutr 93(4): 780-788.
-
Tso P, Balint JA (1986) Formation and transport of chylomicrons by enterocytes to the lymphatics. The American Journal of Physiology 250(6 Pt 1): G715-G726.
-
Kayden HJ, Senior JR, Mattson FH (1967) The monoglyceride pathway of fat absorption in man. The Journal of Clinical Investigation 46(11): 1695-1703.
-
Babayan VK (1987) Medium chain triglycerides and structured lipids. Lipids 22(6): 417-420.
-
Yen CL, Farese RV (2003) MGAT2, a monoacylglycerol acyltransferase expressed in the small intestine. J Biol Chem 278(20): 18532-18537.
-
Yue YG, Chen YQ, Zhang Y, Wang H, Qian Y, et al. (2011) The acyl coenzymeA: monoacylglycerol acyltransferase 3 (MGAT3) gene is a pseudogene in mice but encodes a functional enzyme in rats. Lipids 46(6): 513-520.
-
Nelson DW, Gao Y, Yen MI, Yen CL (2014) Intestine-specific deletion of acyl-CoA:monoacylglycerol acyltransferase (MGAT) 2 protects mice from diet induced obesity and glucose intolerance. The Journal of Biological Chemistry 289(25): 17338-17349.
-
Smith SJ, Cases S, Jensen DR, Chen HC, Sande E, et al. (2000) Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nature Genetics 25(1): 87-90.
-
Chen HC, Ladha Z, Smith SJ, Farese RV (2003) Analysis of energy expenditure at different ambient temperatures in mice lacking DGAT1. American Journal of Physiology, Endocrinology and Metabolism 284(1): E213-E218.
-
Stone SJ, Myers HM, Watkins SM, Brown BE, Feingold KR, et al. (2004) Lipopenia and skin barrier abnormalities in DGAT2-deficient mice. The Journal of Biological Chemistry 279(12): 11767-11776.
-
Ferreira L, Lisenko K, Barros B, Zangeronimo M, Pereira L, et al. (2014) Influence of medium-chain triglycerides on consumption and weight gain in rats: A systematic review. Journal of Animal Physiology and Animal Nutrition 98(1): 1-8.
-
Babu AS, Veluswamy SK, Arena R, Guazzi, M, Lavie CJ, et al. (2014) Virgin coconut oil and its potential cardioprotective effects. Postgraduate Medicine 126(7): 76-83.
-
Lockyer S, Stanner S (2016) Coconut oil - a nutty idea. Nutrition Bulletin 41(1): 42-54.
-
Rubin M, Moser A, Vaserberg N, Greig F, Levy Y, et al. (2000) Long-chain fatty acids, in long-term home parental nutrition: A double-blind randomized cross- over study. Nutrition 16(2): 95-100.
-
Jacobson TA, Ito MK, Maki KC, Orringer CE, Bays HE, et al. (2014) National Lipid Association recommendations for patient-centered management of dyslipidemia: part 1 - executive summary. Journal of Clinical Lipidology 8(5): 473-488.
-
Stone NJ, Robinson JG, Lichtenstein AH, Bairey-Merz CN, Blum CB, et al. (2014) American College of Cardiology/ American Heart Association Task Force on Practice. 2013 ACC/AHA guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults: a report of the American College of Cardiology/ American Heart Association Task Force on Practice Guidelines. Circulation 25(2): S1-S45.
-
Liu R, Cheng M, Kothapalli KSD, Wang Z, Mendralla E, et al. (2019) Glycerol derived process contaminants in refined coconut oil induce cholesterol synthesis in HepG2 cells. Food and Chemical Toxicology 127(1): 135-142.
-
Pullinger CR, Eng C, Salen G, Shefer S, Batta AK, et al. (2002) Human cholesterol 7α-hydroxylase (CYP7A1) deficiency has a hypercholesterolemic phenotype. The Journal of Clinical Investigation 110(1): 109-117.
-
Su X, Abumrad NA (2009) Cellular fatty acid uptake: a pathway under construction. Trends in Endocrinology and Metabolism 20(2): 72-77.
-
Hotamisligil GS, Johnson RS, Distel RJ, Ellis R, Papaioannou VE, et al. (1996) Uncoupling of obesity from insulin resistance through a targeted mutation in aP2, the adipocyte fatty acid binding protein. Science 274(5291): 1377-1379.
-
Queipo-Ortuno MI, Escote X, Ceperuelo-Mallafre V, Garrido-Sanchez L, Miranda M, et al. (2012) FABP4 dynamics in obesity: discrepancies in adipose tissue and liver expression regarding circulating plasma levels. PLoS One 7(11): e48605.
-
Hoekstra M, Stitzinger M, van Wanrooij EJ, Michon IN, Kruijt JK, et al. (2006) Microarray analysis indicates an important role for FABP5 and putative novel FABPs on a Western-type diet. Journal of Lipid Research 47(10): 2198-2207.
-
Ooi GT, Tseng LY, Tran MQ, Rechler MM (1992) Insulin rapidly decreases insulin-like growth factor-binding protein-1 gene transcription in streptozotocin-diabetic rats. Molecular Endocrinology 6(12): 2219-2228.
-
Watanabe M, Houten SM, Mataki C, Christoffolete MA, Kim BW, et al. (2006) Bile acids induce energy expenditure by promoting intracellular thyroid hormone activation. Nature 439(7075): 484-489.
-
Watanabe M, Houten SM, Wang L, Moschetta A, Mangelsdorf DJ, et al. (2004) Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. The Journal of Clinical Investigation 113(10): 1408- 1418.
-
Siegel R, Ma J, Zou Z, Jemal A (2014) Cancer statistics, 2014. CA: a Cancer Journal for Clinicians 64(1): 9-29.
-
National Cancer Institute (2014).
-
Krahmer N, Farese RV, Walther TC (2013). Balancing the fat: lipid droplets and human disease. EMBO Molecular Medicine 5(7): 973-983.
-
Skrivan M, Marounek M, Englmaierova M, Cermak L, Vickova J, et al. (2018) Effect of dietary fat type on intestinal digestibility of fatty acids, fatty acids profiles of breast meat and abdominal fat, and mRNA expression of lipid-related genes in broiler chickens. PLos One 13(4): e0196035.
-
Voskuhl R (2011) Sex differences in autoimmune diseases. Biology of Sex Differences 2(1):1.
-
Klein SL, Flanagan KL (2016) Sex differences in immune responses. Nature Reviews Immunology 16(10): 626-638.
-
Jaillon S, Berthenet K, Garlanda C (2019) Sexual dimorphism in innate immunity. Clin Rev Allergy Immunol 56(3): 308-321.
-
Alswat KA (2017) Gender disparities in osteoporosis. J Clin Med Res 9(4): 382-387.
-
Rainville JR, Tsyglakova M, Hodes GE (2018) Deciphering sex differences in the immune system and depression. Frontiers in Neuroendocrinology 50(1): 67- 90.
-
Laws KR, Irvine K, Gale TM (2018) Sex differences in Alzheimer’s disease. Curr Opin Psychiatry 31(2): 133- 139.
-
Werling DM, Geschwind DH (2013) Sex differences in autism spectrum disorders. Curr Opin Neurol 26(2): 146-153.
-
Hong J, Stubbins RE, Smith RR, Harvey AE, Núñez NP, et al. (2009) Differential susceptibility to obesity between male, female and ovariectomized female mice. Nutr J 8(1): 11.
-
Grove KL, Fried SK, Greenberg AS, Xiao XQ, Clegg DJ, et al. (2010) A microarray analysis of sexual dimorphism of adipose tissues in high-fat diet-induced obese mice. International Journal of Obesity 34(6): 989-1000.
-
Vieira PVJ, Strissel KJ, Xie C, Chang E, Bennett G, et al. (2012) Adipose tissue inflammation and reduced insulin sensitivity in ovariectomized mice occurs in the absence of increased adiposity. Endocrinology 153(9): 4266-4277.
-
Salinero AE, Anderson BM, Zuloaga KL (2018) Sex differences in the metabolic effects of diet-induced obesity vary by age of onset. Int J Obes 42(5): 1088-1091.
-
Joe AWB, Yi L, Even Y, Vogl AW, Rossi FMV (2009) Depot-specific differences in adipogenic progenitor abundance and proliferative response to high-fat diet. Stem Cells 27(10): 2563-2570.
-
Jeffery E, Wing A, Holtrup B, Sebo Z, Kaplan JL, et al. (2016) The adipose tissue microenvironment regulates depot-specific adipogenesis in obesity. Cell Metab 24(1): 142-150.
-
Wu Y, Lee MJ, Ido Y, Fried SK (2017) High-fat diet- induced obesity regulates MMP3 to modulate depot- and sex-dependent adipose expansion in C57BL/6J mice. Am J Physiol Endocrinol Metab 312(1): E58-E71.
-
Flegal KM, Kruszon-Moran D, Carroll MD, Fryar CD, Ogden CL, et al. (2016) Trends in obesity among adults in the United States, 2005 to 2014. JAMA 315(21): 2284- 2291.
-
Karastergiou K, Smith SR, Greenberg AS, Fried SK (2012) Sex differences in human adipose tissues - the biology of pear shape. Biol Sex Differ 3(1): 13.
-
Karpe F, Pinnick KE (2015) Biology of upper-body and lower-body adipose tissue-link to whole-body phenotypes. Nat Rev Endocrinol 11(2): 90-100.
-
Karastergiou K, Fried SK (2017) Cellular mechanisms driving sex differences in adipose tissue biology and body shape in humans and mouse models. Adv Exp Med Biol 1043(1): 29-51.
-
Wang QA, Tao C, Gupta RK, Scherer PE (2013) Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat Med 19(10): 1338-1344.
-
Jeffery E, Church CD, Holtrup B, Colman L, Rodeheffer MS, et al. (2015) Rapid depot-specific activation of adipocyte precursor cells at the onset of obesity. Nat Cell Biol 17(4): 376-385.
-
Zamboni M, Armellini F, Milani MP, De Marchi M, Todesco T, et al. (1992) Body fat distribution in pre- and post-menopausal women: metabolic and anthropometric variables and their inter-relationships. Int J Obes Relat Metab Disord 16(7): 495-504.
-
Garaulet M, Pérez-Llamas F, Baraza JC, Garcia- Prieto MD, Fardy PS, et al. (2002) Body fat distribution in pre- and post-menopausal women: metabolic and anthropometric variables. J Nutr Health Aging 6(2): 123- 126.
-
Maas AHEM, Appelman YEA (2010) Gender differences in coronary heart disease. Neth Heart J 18(12): 598-602.
-
Lima R, Wofford M, Reckelhoff JF (2012) Hypertension in postmenopausal women. Current Hypertension Report 14(3): 254-260.
-
Araujo AB, Dixon JM, Suarez EA, Murad MH, Guey LT, et al. (2011) Clinical review: endogenous testosterone and mortality in men: a systematic review and meta- analysis. J Clin Endocrinol Metab 96(10): 3007-3019.
-
Blouin K, Boivin A, Tchernof A (2008) Androgens and body fat distribution. J Steroid Biochem Mol Biol 108(3-5): 272-280.
-
Ruige JB, Mahmoud AM, De Bacquer D, Kaufman JM (2011) Endogenous testosterone and cardiovascular disease in healthy men: a meta-analysis. Heart 97(11): 870-875.
-
Santosa S, Bonnes SL, Jensen MD (2016) Acute female hypogonadism alters adipose tissue fatty acid storage factors and chylomicronemia. J Clin Endocrinol Metab 101(5): 2089-2098.
-
Santosa S, Bush NC, Jensen MD (2017) Acute testosterone deficiency alters adipose tissue fatty acid storage. J Clin Endocrinol Metab 102(8): 3056-3064.
-
Global Health Data Exchange (2017) Global Burden of Disease Study 2015 (GBD 2015) Obesity and Overweight Prevalence 1980-2015. Seattle, Institute for Health Metrics and Evaluation (IHME): United States.
-
Swinburn BA, Sacks G, Hall KD, McPherson K, Finegood DT, et al. (2011) The global obesity pandemic: shaped by global drivers and local environments. Lancet 378(9793): 804-814.
-
NCD-RisC (2016) Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 387(10026): 1377-1396.
-
Kim DD, Basu A (2016) Estimating the Medical Care Costs of Obesity in the United States: Systematic Review, Meta-Analysis, and Empirical Analysis. Value in Health 19(5): 602-613.
-
United States Department of Agriculture (2015) Dietary Guidelines for Americans – 2015–2020.
-
World Health Organization (2018) Healthy diet.
-
Chooi YC, Ding C, Magkos F (2019) The epidemiology of Obesity. Metabolism 92(1): 6-10.
-
Wang S, Lay S, Yu H, Shen S (2016) Dietary guidelines for Chinese residents. Comments and comparisons. J Zhejiang Univ Sci B 17(6): 649-656.
-
von Lengerke T, Krauth C (2011) Economic costs of adult obesity: a review of recent European studies with a focus on subgroup-specific costs. Maturitas 69(3): 220- 229.
-
European Food Safety Authority (2019) Dietary reference values for nutrients - Summary report.
-
European Food Safety Authority (2011) Scientific Opinion on the substantiation of health claims related to medium-chain triglycerides and reduction in body weight (ID 643, 677, 1614) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA Journal 9(6): 2240.
-
Sacks FM, Lichtenstein AH, Wu JHY, Appel LJ, Creager MA, et al. (2017) Dietary fats and cardiovascular disease: A presidential advisory from the American Heart Association. Circulation 136(3): e1–e23.
-
European Union. Commission Regulation (2012) No 231/2012 of 9 March 2012 laying down specifications for food additives listed in Annexes II and III to Regulation (EC) No 1333/2008 of the European Parliament and of the Council. OJEC 55, L83/198.
-
SBEM, ABESO (2015) Posicionamento oficial da Sociedade Brasileira de Endocrinologia e Metabologia (SBEM) e da Associação Brasileira para o Estudo da Obesidade e da Síndrome Metabólica (ABESO) sobre o uso do óleo de coco para perda de peso.
-
Agencia Nacional de Vigilancia Sanitaria (ANVISA) (2005) RDC Resolution 270, of September 22, 2005. Approves the technical regulation for vegetable oils, vegetable fats and cream vegetable. Official Gazette of the Federal Republic of Brazil, Brasília. DF.
-
Ippagunta S, Hadenfeldt TJ, Milner JL, Hargrave- Barnes KM (2011) Dietary conjugated linoleic acid induces lipolysis in adipose tissue of coconut oil-fed mice but not soy oil-fed mice. Lipids 46(9): 821-830.
-
Diaz-Villasenor A, Granados O, Gonzalez-Palacios B, Tovar-Palacio C, Torre-Villalvazo I, et al. (2013) Differential modulation of the functionality of white adipose tissue of obese Zucker (fa/fa) rats by the type of protein and the amount and type of fat. J Nutr Biochem 24(11): 1798-1809.
-
Ippagunta S, Angius Z, Sanda M, Barnes KM (2013) Dietary CLA-induced lipolysis is delayed in soy oil-fed mice compared to coconut oil-fed mice. Lipids 48(11): 1145-1155.
-
Patrone V, Minuti A, Lizier M, Miragoli F, Lucchini F, et al. (2018) Differential effects of coconut versus soy oil on gut microbiota composition and predicted metabolic function in adult mice. BMC Genomics 19(1): 808.
-
Olivares-Garcia V, Torre-Villalvazo I, Velazquez- Villegas L, Aleman G, Lara N, et al. (2015) Fasting and postprandial regulation of the intracellular localization of adiponectin and of adipokines secretion by dietary fat in rats. Nutr Diabetes 5(11): e184.
-
Feranil AB, Duazo PL, Kuzawa CW, Adair LS (2011) Coconut oil predicts a beneficial lipid profile in pre- menopausal women in the Philippines. Asia Pacific Journal of Clinical Nutrition 20(2): 190-195.
-
Kinsella R, Maher T, Clegg ME (2017) Coconut oil has less satiating properties than medium chain triglyceride oil. Physiology and Behavior 179(1): 422-426.
-
Deol P, Kozlova E, Valdez M, Ho C, Yang E-W, et al. (2020) Dysregulation of hypothalamic gene expression and the oxytocinergic system by soybean oil diets in male mice. Endocrinology 161(2): bqz044.
-
Zhao M, Zang B, Cheng M, Ma Y, Yang Y (2013) Differential responses of hepatic endoplasmic reticulum stress and inflammation in diet-induced obese rats with high-fat diet rich in lard oil or soybean oil. PLoS One 8(11): e78620.
-
Wang J, Yan S, Xiao H, Zhou H, Liu S, et al. (2017) Anti-obesity effect of a traditional Chinese dietary habit- blending lard with vegetable oil while cooking. Scientific Reports 7(1): 14689.
-
Tung YT, Hsu YJ, Chien YW, Huang CC, Huang WC, et al. (2019) Tea seed oil prevents obesity, reduces physical fatigue, and improves exercise performance in High- Fat-Diet-Induced obese ovariectomized mice. Molecules 24(5): 980.
-
Gao Y, Li X, Gao Q, Fan L, Jin H, et al. (2019) Differential effects of olive oil, soybean oil, corn oil and lard oil on carbon tetrachloride-induced liver fibrosis in mice. Bioscience Reports 39(10): BSR20191913.
-
Yan S, Zhou H, Liu S, Wang J, Zeng Y, et al. (2020) Differential effects of Chinese high-fat dietary habits on lipid metabolism: mechanisms and health implications. Lipids in Health and Disease 19(1): 30.
-
Bueno AA, Oyama LM, de Oliveira C, Pisani LP, Ribeiro EB, et al. (2008) Effects of different fatty acids and dietary lipids on adiponectin gene expression in 3T3-L1 cells and C57BL/6J mice adipose tissue. Pflugers Archiv – European Journal of Physiology 455(4): 701-
-
Firestone D (2013) Physical and Chemical Characteristics of Oils, Fats, and Waxes. 3rd (Edn.), IL: AOCS Press, Urbana.
-
Sankararaman S, Sferra TJ (2018) Are we going nuts on coconut oil? Current Nutrition Reports 7(3): 107-115.
-
Derewiaka D, Sosinska E, Obiedzinski M, Krogulec A, Czaplicki S, et al. (2013) Determination of the adulteration of butter. European Journal of Lipid Science and Technology 113(8): 1005-1011.
-
Hofmann AF, Poley JR (1972) Role of bile acid malabsorption in pathogenesis of diarrhea and steatorrhea in patients with ileal resection. I. Response to cholestyramine or replacement of dietary long chain triglyceride by medium chain triglyceride. Gastroenterology 62(5): 918-934.
-
Heck AM, Yanovski JA, Calis KA (2000) Orlistat, a new lipase inhibitor for the management of obesity. Pharmacotherapy 20(3): 270-279.
-
Rizzo M (2010) Lomitapide, a microsomal triglyceride transfer protein inhibitor for the treatment of hypercholesterolemia. IDrugs 13(2):103-111
-
Dow RL, Li J, Pence MP, Gibbs EM, LaPerle JL, et al. (2011) Discovery of PF-04620110, a potent, selective, and orally bioavailable inhibitor of DGAT-1. ACS Medical Chemistry Letters 2(5): 407-412.
-
Okuma C, Ohta T, Tadaki H, Hamada H, Oda T, et al. (2015) JTP-103237, a novel monoacylglycerol acyltransferase inhibitor, modulates fat absorption and prevents diet- induced obesity. Eur J Pharmacol 758(1): 72-81.
- Investigation of Polymorphisms in PPAR-Ɣ and TRHR Genes and their Impact on Turkish Diabetic and Obese Individuals
- The Impact of Aircraft Noise Exposure on the Efficacy of Empagliflozin Therapy in an Animal Model of Obesity
- Rooibos Mitigates Metabolic and Inflammatory Dysfunctions in Mice Fed a High-Carbohydrate Diet
- Synergistic Effect of Combined Leaf Extract of Vernonia amygdalina, Ocimum gratissimum, and Zingiber officinale Tuber on Phytochemical Profile, Antioxidant Activity, Serum Insulin, and Biochemical Parameters in Streptozotocin-Induced Diabetic Rats
- Investigation of Cardiovascular Responses to Aerobic Exercise in Obese University Students
- A Look at the Phase Angle Obtained by Electrical Bioimpedance