Rooibos Mitigates Metabolic and Inflammatory Dysfunctions in Mice Fed a High-Carbohydrate Diet
The global rise in metabolic disorders is strongly associated with excessive carbohydrate intake and physical inactivity. Rooibos (Aspalathus linearis), a South African plant, has been recognized for its antioxidant, immunomodulatory, and hepatoprotective properties, though its role in obesity-related metabolic dysfunction remains unclear. This study evaluated the effects of Rooibos tea on metabolic disturbances induced by a high-carbohydrate (HC) diet in mice. Male Balb-C mice were divided into four groups: Control (C, balanced diet), HC (high-carbohydrate diet), HCR (HC diet plus Rooibos tea for 8 weeks), and HCRT (HC diet followed by Rooibos tea for the final 2 weeks). Body weight and food intake were monitored. Blood was analyzed for glucose, glucose tolerance test, lipids, liver enzymes, cytokines (TNF-α, IL-6, IL-10), and nitric oxide (NO). Liver were collected to assess weight, lipid content, cytokine, NO, and histological changes. The HC diet increased adiposity, glucose intolerance, hepatic triglycerides, inflammatory markers, and induced hepatic steatosis. Rooibos supplementation significantly improved these alterations, reducing inflammation, improving glucose tolerance, and attenuating liver damage. These findings support Rooibos tea as a potential dietary strategy to prevent or ameliorate HC diet-induced metabolic dysfunction.
Abbreviations
WHO: World Health Organization; AT: Adipose Tissue; WAT: White Adipose Tissue; TNF: Tumor Necrosis Factor; ROS: Reactive Oxygen Species; MAFLD: Metabolic Dysfunction-Associated Fatty Liver Disease; T2DM: Type
2 Diabetes Mellitus; EAT: Epididymal Adipose Tissue; RAT: Retroperitoneal Adipose Tissue; GLU: Glucose; TC: Total Cholesterol; TAG: Triglycerides; AST: Aspartate Aminotransferase; ALT: Alanine Aminotransferase; GTT: Glucose Tolerance Test.
Introduction
Changes in dietary patterns, such as the increased consumption of high carbohydrate diets and processed foods, combined with low or no physical activity have been identified as major contributors to the rise in body weight and obesity. According to the World Health Organization (WHO), the incidence of obesity among adults has tripled since 1975. In 2021, an estimated 1 billion adults were classified as overweight or obese [1]. If current trends continue, it is projected that by 2050, the number of adults living with overweight or obesity will reach approximately 3.8 billion, representing more than half of the anticipated global adult population [1, 2].
Obesity is strongly related to health risk , and it is diagnosed mainly by body mass index (>30) [2]. One of the main alterations in obesity is the excessive adipose tissue (AT), which is a multifunctional organ that exerts beyond roles other than just storage energy in form of lipids [3]. Different types of adipose tissue (e.g.: White, Beige and brown) perform distinct metabolic functions and differ in cellular composition and structural characteristics. White adipose tissue (WAT) is composed mainly of adipocytes containing a large unilocular lipid droplet and is primarily involved in energy storage and mobilization, as well as the secretion of adipokines [4]. When energy metabolism becomes imbalanced, the abnormal secretion of adipokines often leads to adipose tissue dysfunction and the development of obesity. Obesity is typically associated with elevated levels of C-reactive protein, pro-inflammatory cytokines (e.g., IL- 1β, tumor necrosis factor (TNF) and interleukin-6 (IL-6)), reactive oxygen species (ROS), pro-inflammatory leukocyte phenotypes, and an imbalanced production of lipid mediators compared to healthy individuals [4, 5, 6].
In individuals with obesity, systemic low-grade inflammation, along with more pronounced inflammation within white adipose tissue, promotes insulin resistance and contributes to harmful crosstalk between adipose tissue and other organs [7, 8]. In this context, the liver is one of the organs most likely to be affected. Metabolic dysfunction- associated fatty liver disease (MAFLD/NAFLD) is among the most prevalent chronic liver conditions worldwide and has increasingly become the leading form, affecting up to 38% of the global adult population [9]. MAFLD is diagnosed based on the presence of hepatic steatosis in combination with at least one of the following metabolic conditions: overweight/ obesity, type 2 diabetes mellitus (T2DM), or metabolic dysfunction [10]. Several distinct metabolic profiles may contribute to the development of MAFLD, including disorders in lipid, glucose, and bile acid metabolism; trace element accumulation; mitochondrial dysfunction, and alterations in immune metabolism [11, 12, 13, 14, 15, 16, 17].
Considering that MAFLD involves both inflammatory processes and oxidative stress, some studies have highlighted the potential use of natural anti-inflammatory and antioxidant agents, particularly those derived from plants [18]. In this context, the search for natural plant- based compounds with promising effects in the treatment or prevention of metabolic diseases appears to be of great interest, as the currently available therapeutic options are not fully effective. One such candidate is Rooibos (Aspalathus linearis), for which scientific evidence has demonstrated several medicinal and therapeutic properties, including antioxidant, anti-inflammatory, anticancer, and chemopreventive effects, among others [19].
Rooibos is a plant from the Fabaceae family that grows predominantly in South Africa [20]. The unfermented product retains its green color and is referred to as green rooibos. During fermentation, the color changes from green to red due to the partial oxidation of constituent polyphenols, which is why the final product is often called red tea [21].
Rooibos contains a wide variety of compounds, such as dihydrochalcones, flavanones, flavones, flavonols, lignans, hydroxycinnamic acids, phenolic carboxylic acids, linearthin, aspalathin, nothofagin among others [22, 23]. Moreover, studies using animal models have shown that rooibos tea possesses potent antioxidant, immunomodulatory, hypoglycemic, hypolipidemic, and gastrointestinal integrity- enhancing properties [21, 22, 23, 24, 25, 26, 27, 28].
Given the numerous biological activities of rooibos, we believe it may be a promising candidate for the treatment and/ or prevention of metabolic disorders. Therefore, the present study aimed to evaluate the effects of Rooibos administration on metabolic, immunological, and histological alterations in animals fed a high-carbohydrate diet.
Methods
Animals and Treatment
Thirty-two (32) male Balb-C mice, with an initial body weight ranging between 19 - 25 g, were used in this study. The animals were housed in the animal facility of the Biochemistry Laboratory at the Federal University of Triângulo Mineiro under controlled conditions (12-hour light/dark cycle, ventilated shelving system, temperature maintained at 22 ± 2°C). All procedures were approved by the Ethics Committee on the Use of Animals (CEUA) at the Federal University of Triângulo Mineiro (Protocol Number: 23085.002424/2018-59).
The animals were randomly assigned to the following experimental groups (n=8). Group 1 (C): Fed a normocaloric Nuvital diet (4.0 Kcal/g) composed of carbohydrates (65.8%), fats (3.1%), and proteins (31.1%), with water provided ad libitum; Group 2 (HC): Fed a high-carbohydrate (HC) diet (4.4 Kcal/g) composed of carbohydrates (74.2%), fats (5.8%), and proteins (20.0%), with water ad libitum; Group 3 (HCR): Fed the HC diet with Rooibos tea provided ad libitum in place of water; Group 4 (HCRT): Fed the HC diet with water ad libitum for six weeks, followed by Rooibos tea replacing water during the final two weeks.
The HC diet consisted of a previously standardized mixture containing 45% normocaloric Nuvital diet, 45% sweetened condensed milk, and 10% refined sugar [29]. Water/tea and food intake were monitored daily, while body weight was recorded weekly. After eight weeks on their respective diets, the animals underwent the designated experimental procedures.
Preparation of Rooibos Tea
Commercially available Rooibos red tea was used. The tea was prepared daily by infusing 10 g of tea leaves in 1000 mL of boiling water for 5 minutes. The solution was then filtered to remove solid residues, cooled to room temperature, and provided to the animals.
Collection of Biological Material
At the end of the experimental period, the animals were anesthetized with a solution of 10% ketamine (0.01 mg/g body weight) and 2% xylazine (0.1 mg/g body weight). Blood was collected via cardiac puncture. Epididymal adipose tissue (EAT), retroperitoneal adipose tissue (RAT), and liver were excised and weighed. A portion of the liver was fixed in formalin and subsequently stored in 70% ethanol for histological analysis. Another portion was stored at −80°C for cytokine analysis. Serum samples were stored at −20°C.
Biochemical Assays
Glucose (GLU), total cholesterol (TC), triglycerides (TAG), aspartate aminotransferase (AST), and alanine aminotransferase (ALT) levels were measured using commercial kits (Bioclin®). The results were expressed in mg/dL.
Glucose Tolerance Test (GTT)
After 7 weeks of treatment, the glucose tolerance test was performed on the HC, HCR, and HCRT groups. Animals were weighed and fasted for 6 hours with free access to water. At time zero (0 min), blood samples were collected via caudal vein puncture to determine baseline glycemia. Subsequently, a 20% glucose solution was administered intraperitoneally at a dose of 2 g glucose/kg body weight.
Blood glucose levels were measured at 30, 60, and 120 minutes using a glucometer (FreeStyle Optium Neo ®). The results were expressed in mg/dL.
Lipid Extraction and Quantification
A portion of the liver from each animal was used for total lipid (TL) quantification by the Folch method [30]. After total lipid quantification, the extracted fat was resuspended in 1 mL of isopropyl alcohol for hepatic total cholesterol (TCh) and hepatic triglyceride (TAGh) assays, using commercial kits (Bioclin®). The results were expressed in g/100g of liver.
Cytokine Assay
Serum levels of the pro-inflammatory cytokines TNF-α and IL-6, as well as the anti-inflammatory cytokine IL-10, were measured using ELISA kits (BD Biosciences ®). Liver tissues stored at -80°C were homogenized in PBS buffer containing protease inhibitors and 1% NP-40. After homogenization and centrifugation (12,000 RCF for 30 min), the supernatant was used to cytokine quantification using ELISA kits (BD Biosciences ® ). The results were expressed in pg/mL.
Protein and Nitric Oxide (NO) Quantification
Protein concentration was determined using a NanoDrop 2000 spectrophotometer (Protein A280 software), with PBS (0.2 µL) serving as the blank. Nitric oxide quantification was performed using the Griess reaction [31]. The results were expressed in mM/mg.
Histology
Liver samples were evaluated using hematoxylin and eosin (H&E) staining. Images were captured with a ZEISS microscope equipped with an AxioCam ICc 5 camera and analyzed using AxioVision SE64 software.
Statistical Analysis
Group comparisons were performed using one-way ANOVA followed by Tukey’s post-hoc test. Results are expressed as mean ± standard error (SE), with significance set at p < 0.05.
Results
Although no statistically significant differences were observed in body weight (Figure 1) or food intake among the experimental groups, analysis of adipose tissue distribution revealed a marked increase in RAT weight in the HC (~92%), HCR (~130%), and HCRT (~84%) groups compared to the C group. In contrast, EAT and liver weights remained unchanged across all groups.
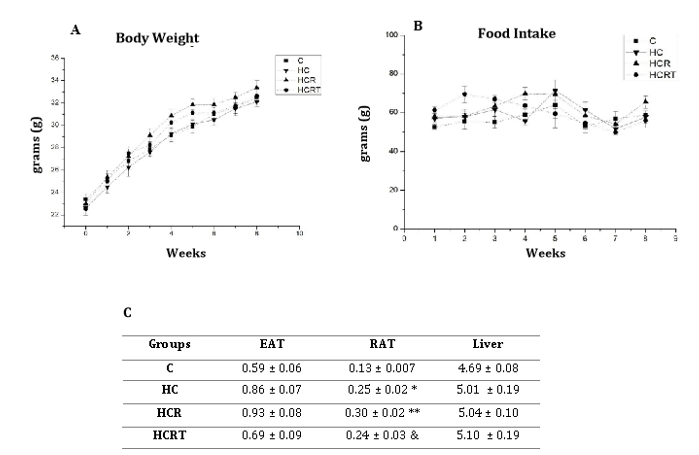
Figure 1: 1A: Body weight of all experimental groups per week; 1B: Food intake of all experimental groups per week; 1C. Weight (g/100g BW) of epididymal adipose tissue (EAT), retroperitoneal adipose tissue (RAT), and liver. C, control diet and water;HC, high-carbohydrate diet and water; HCR, high-carbohydrate diet and Rooibos tea; HCRT, high-carbohydrate diet and water for 6 weeks, followed by Rooibos tea replacing water in the last 2 weeks. Results are expressed as mean ± standard error (n = 8 animals per group). *p<0.05 HC vs C; **p<0,05 HCR vs C; & p<0,05 HCRT vs C.
In the assessment of carbohydrate metabolism, the HC group showed a ~59% increase in random blood glucose levels compared to the C group, while Rooibos treatment did not significantly alter this parameter. Further assessment through the glucose tolerance test (GTT) revealed no significant differences in glycemic levels among the groups at 0 and 30 minutes. In contrast, at 60 minutes following glucose administration, the HCR and HCRT groups demonstrated a significant reduction in blood glucose levels of approximately 30% and 38%, respectively, compared to the HC group. Similarly, at 120 minutes, both the HCR and HCRT groups exhibited an ~21% reduction in glucose levels relative to the HC group. The area under the curve (AUC) was also significantly reduced in the HCR (~37%) and HCRT (~31%) groups compared to the HC group (Figure 2).
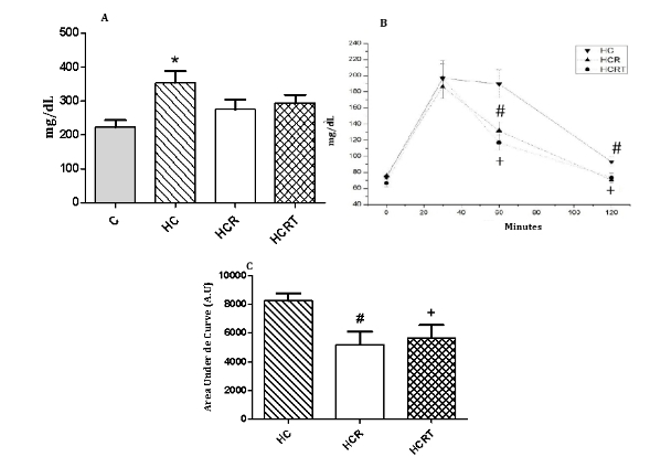
Figure 2: 2A: Random blood glucose levels of all experimental groups; 2B. Glucose tolerance test (GTT) of the experimental groups. 2C. Area under Curve of GTT experiemental groups. C, control diet and water; HC, high-carbohydrate diet and water; HCR, high-carbohydrate diet and Rooibos tea; HCRT, high-carbohydrate diet and water for 6 weeks, followed by Rooibos tea replacing water in the last 2 weeks. Results are expressed as mean ± standard error (n = 8 animals per group). *p<0.05 HC vs C; #p<0.05 HCR vs HC; +p<0.05 HCRT vs HC.
Lipid metabolism was evaluated by measuring serum lipid profiles, liver function biomarkers, and hepatic lipid content. TC levels were approximately ~41% higher in the HC group compared to the C group. TAG levels were reduced in the HCRT (42%) and (34%) compared to the C and HC group respectively (Table 1). TAGh content showed a significant increase in the HC (~45%) and HCR (~34%) groups relative to C (Table 2). No significant differences were observed in the liver enzymes AST and ALT or other hepatic lipid parameters in among the experimental groups (Table 2).
| Groups | TAG | TC | AST | ALT |
|---|---|---|---|---|
| c | 326.73 ± IS.SO | IS3 .28 ± I6.29 | SI.SO ± 4.49 | 23.16 ± 3.10 |
| HC | 291.I2 ± 30 .80 | 2I6.83 ± I3.94* | 40 .42 ± 4.83 | I7.!6 ± 1.60 |
| HCR | 229 .02 ± 27.16 | I69 .85 ± I6.34 | 4S .42 ± 2.02 | 22 .40 ± 1.03 |
| HCRT | I90 .70 ± Il.84 &,+ | I63.83 ± 6.75 | 40 .28 ± 2.SI | 20 .40 ± 2.56 |
Table 1: ** Serum levels of triglycerides (TAG), cholesterol (CTO), AST and ALT in the experimental groups.
C: Control diet and water; HC: high-carbohydrate diet and water; HCR: high-carbohydrate diet and Rooibos tea; HCRT: high- carbohydrate diet and water for 6 weeks, followed by Rooibos tea replacing water in the last 2 weeks. Results are expressed as mean ± standard error (n = 8 animals per group). &p<0.05 HCRT vs C; + p<0,05 HCRT vs HC; *p<0.05 HC vs C. Table 1: Serum levels of triglycerides (TAG), cholesterol (CTO), AST and ALT in the experimental groups.
| Groups | TL | TAGb | TCb |
|---|---|---|---|
| c | 1.04 ± 0.01 | 4.08 ± 0.25 | 1.26 ± 0.05 |
| HC | 1.28 ± 0.07 | 5.95 ± 0.36* | 1.33 ± 0.04 |
| HCR | 1.28 ± 0.04 | 5.47 ± 0.31** | 1.36 ± 0.05 |
| HCRT | 1.30 ± 0.09 | 4.71 ± 0.35 | 1.24 ± 0.06 |
Table 2: ** Quantification of hepatic total lipids (TL), triglycerides (TAGh), and cholesterol (CTOh).
C: control; HC: high-carbohydrate diet and water; HCR: high-carbohydrate diet and Rooibos tea; HCRT: high-carbohydrate diet and water for 6 weeks, followed by Rooibos tea replacing water in the last 2 weeks. Results are expressed as mean ± standard error (n = 8 animals per group). *P<0.05 HC vs C; P<0.05 HCR vs C. Table 2:** Quantification of hepatic total lipids (TL), triglycerides (TAGh), and cholesterol (CTOh).
Systemic inflammation was assessed by measuring several cytokines following Rooibos treatment. Serum TNF-α levels were elevated ~83% in the HC group compared to the C. Notably, TNF-α levels were reduced by about 45% in the HCR group relative to HC. To further explore local inflammatory responses, we also quantified TNF-α in liver tissue. Hepatic TNF-α levels decreased by approximately ~28% and ~30% in the HCR and HCRT groups, respectively, compared to C. A decrease of ~29% in TNF-α levels was also observed in the HCRT group compared to the HC group. It is important to note that there was a tendency for TNF-α levels to decrease (p = 0.08) in the HCR group compared to the HC group. No significant differences were observed in IL-6 and IL-10 concentrations in either serum or liver across the experimental groups (Tables 3 & 4).
| TNF-alfa | IL-6 | IL-10 | |
|---|---|---|---|
| c | 13.85±2.68 | 0.08±0.009 | 1.23±0.12 |
| HC | 25.36±1.8* | 0.07±0.21 | 1.10±0.11 |
| HCR | 13.96±0.84# | 0.104±0.01 | 1.40±0.16 |
| HCRT | 18.16±3.90 | 0.09±0.008 | 1.48±0.12 |
| Groups | TNF-alfa | IL-6 | IL-10 |
| c | 55.24±2.62 | 3.11±0.28 | 1.23±0.12 |
| HC | 54.17±2.49 | 3.75±0.18 | 1.10±0.11 |
| HCR | 39.58±5.32** | 2.73±0.24 | 1.40±0.16 |
| HCRT | 38.38±4.25&,+ | 3.22±0.21 | 1.48±0.12 |
Table 3: ** Serum cytokine levels of TNF-alpha, IL-6, and IL-10.
C: control; HC: high-carbohydrate diet and water; HCR: high-carbohydrate diet and Rooibos tea; HCRT: high-carbohydrate diet and water for 6 weeks; followed by Rooibos tea replacing water in the last 2 weeks.Results are expressed as mean ± standard deviation (n = 8 animals per group).*P<0.05 HC vs C; #P<0.05 HCR vs HC. Table 3: Serum cytokine levels of TNF-alpha, IL-6, and IL-10.
C: control; HC: high-carbohydrate diet and water; HCR: high-carbohydrate diet and Rooibos tea; HCRT: high-carbohydrate diet and water for 6 weeks; followed by Rooibos tea replacing water in the last 2 weeks.Results are expressed as mean ± standard deviation (n = 8 animals per group). p<0.05 HCR vs C; &p<0.05 HCRT vs C; + p<0.05 HCRT vs HC. Table 4:** Hepatic tissue cytokine levels of TNF-alpha, IL-6, and IL-10.
To evaluate oxidative stress and the potential antioxidant effects of Rooibos, we measured NO levels in liver tissue. The HC diet led to a significant increase in hepatic NO levels, with values approximately 45% higher than the control (C) group.
Rooibos supplementation effectively mitigated this response, reducing NO levels by about 29% in the HCR group and 35% in the HCRT group compared to HC (Figure 3).
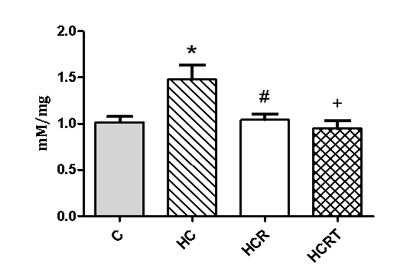
Figure 3: Nitric Oxide Levels in Hepatic Tissue among the experimental groups. C, control; HC, high-carbohydrate diet and water;HCR, high-carbohydrate diet and Rooibos tea;HCRT, high-carbohydrate diet and water for 6 weeks, followed by Rooibos tea replacing water in the last 2 weeks.Results are expressed as mean ± standard deviation (n = 8 animals per group).*P<0.05 HC vs. C; #P<0.05 HCR vs HC; +P<0.05 HCRT vs HC.
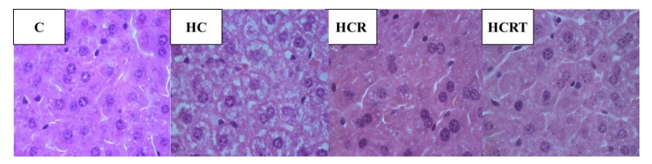
Finally, liver histology was assessed. Mice in the HC group displayed marked hepatocellular changes characterized by pale, swollen hepatocytes indicative of glycogen accumulation and lipid steatosis. Notably, Rooibos supplementation (HCR and HCRT groups) attenuated these histological alterations, with liver sections showing improved cellular morphology and reduced signs of steatosis (Figure 4). Overall, these results demonstrate that the high- carbohydrate diet effectively induced classic metabolic disturbances associated with adiposity and hepatic steatosis, while Rooibos treatment significantly mitigated these effects.
Discussion
In our study, food intake, water/tea consumption (data not shown), body, EAT and liver weight remained unchanged among the groups. However, RAT weight was significantly higher in mice submitted to HC diet. These findings are consistent with previous studies, demonstrating that rats fed a powdered Purina Chow diet containing corn oil and condensed milk exhibited increased body fat accompanied by an elevated cell number in retroperitoneal and inguinal, but not epididymal, fat pads [32]. Similarly, Oliveira MC, et al. [29] showed that an HC diet induces rapid adipose tissue expansion without significant changes in overall body weight.
We hypothesized that Rooibos supplementation might prevent adipose tissue expansion in the HCR and HCRT groups, given that its phenolic compounds inhibit adipogenesis-related transcription factors in vitro [33]. However, this effect was not observed in vivo, likely due to Rooibos compound digestion, metabolism, and lower systemic bioavailability compared to direct in vitro exposure. Supporting this, Kotzé-Hörstmann LM, et al. [34] found that green Rooibos extract did not prevent visceral adiposity or affect body weight in animals on high-calorie diets. Further research, including dose optimization and direct cell and animal experiments comparisons, is needed to better define Rooibos’s impact on adipose tissue metabolism.
Given the increase in retroperitoneal adipose tissue (RAT), we examined its potential impact on carbohydrate and lipid metabolism. The HC group showed elevated random blood glucose levels compared to control. Notably, Rooibos supplementation (HCR and HCRT groups) significantly lowered glycemia at 60 and 120 minutes during the glucose tolerance test (GTT) versus the HC group. These results suggest that the HC diet induces insulin resistance, while Rooibos may improve insulin sensitivity and glucose uptake.
Diets high in simple carbohydrates, like the HC diet, cause rapid fat gain linked to metabolic problems such as insulin resistance and inflammation [29]. Fat tissue expansion attracts macrophages that release pro-inflammatory cytokines (IL-6, TNF-α, etc.) and lower adiponectin, an anti-inflammatory hormone that normally protects against insulin resistance [35, 36]. These inflammatory signals, especially TNF-α, disrupt insulin signaling by reducing GLUT4 and IRS function, worsening glucose intolerance [37]. Given these mechanisms, we propose that the improvement in glucose metabolism observed with Rooibos treatment in HC diet-fed animals may be mediated, at least in part, by its anti-inflammatory properties. Rooibos contains bioactive compounds like aspalathin and quercetin that exert anti- inflammatory effects by downregulating TNF-α, IL-6, NF-κB expression, and inhibiting protein kinases and arachidonic acid pathway enzymes [36, 38]. Although in our study we did not observe an increase in the proinflammatory cytokine IL-6 we found an increase in TNF-α levels in the HC group, which were reduced in mice treated with Rooibos tea. The lack of change in IL-6 levels suggesting limited systemic inflammatory response in this model [39, 40]. Additionally, aspalathin and nothofagin inhibit SGLT2, reducing renal glucose reabsorption and lowering blood glucose levels [41]. Rooibos-derived flavones also inhibit hepatic α-glucosidase more effectively than acarbose, limiting intestinal glucose absorption and postprandial hyperglycemia [41]. Moreover, phenylpyruvic acid from Rooibos may protect pancreatic β-cells by reducing apoptosis or stimulating neogenesis, potentially enhancing insulin secretion and glycemic control [37, 41].
Regarding lipid metabolism, serum lipid analysis revealed no significant differences in TAG between control (C) and HC groups. However, TC levels increased in the HC group, consistent with the known effects of refined carbohydrates on insulin resistance, lipogenesis, and inflammatory pathways [29]. Rooibos tea reduced TAG levels, likely due to its bioactive compounds, as aspalathin, which activates AMPK and enhances β-oxidation, and quercetin, which exerts anti-inflammatory and anti-atherogenic effects [36, 38].
Hepatic inflammation typically involves increased cytokines like IL-1β, IL-6, IL-12, and TNF-α from adipocytes, Kupffer cells, and steatotic hepatocytes, leading to fibrosis and liver injury, however no significant TNFα and IL-6 elevation was detected in liver from HC group. Findings imply that the local inflammatory response in this model is relatively limited [39, 40, 41, 42, 43]. However, Rooibos supplementation effectively reduced TNF-α levels in liver, aligning with findings from LPS-induced liver injury models [44]. This anti- inflammatory effect is likely mediated by Rooibos flavonoids (e.g.: luteolin, rutin, and quercetin) which inhibit Kupffer cell activation and pro-inflammatory cytokine synthesis [44]. No significant changes in IL-10 levels were observed in either serum or hepatic tissue suggesting that Rooibos acts mainly by suppressing pro-inflammatory mediators rather than enhancing anti-inflammatory pathways, as similarly reported in other tissues after Rooibos treatment [45].
Oxidative stress, a key contributor to metabolic dysfunction, was evidenced in the HC group by elevated hepatic nitric oxide (NO) levels, reflecting oxidative damage, as similarly reported with hypercaloric diets [46]. Rooibos supplementation significantly reduced NO levels in HCR and HCRT groups, indicating antioxidant activity. This effect is consistent with prior findings on Rooibos flavonoids, especially aspalathin, known for their potent free radical- scavenging properties [47, 48]. The antioxidant properties of Rooibos flavonoids may support improved glycemic control by mitigating oxidative stress, a known driver of insulin resistance [36, 47, 48].
Although liver weight remained unchanged, hepatic triglyceride levels were elevated in HC and HCR groups, pointing to disrupted lipid metabolism, consistent with reports linking high-glycemic diets to hepatic lipogenesis, oxidative stress, and injury [49]. Histological analysis revealed alterations in the HC group, characterized by swollen hepatocytes, glycogen accumulation, and lipid vacuoles, confirming progression toward MAFLD via de novo lipogenesis and fatty acid overload [50, 51, 52]. Elevated hepatic TNF-α and NO levels further supported the presence of MAFLD, as previously observed in high-fat diet models [10, 53]. Notably, Rooibos-treated animals, particularly the HCR group, showed histological improvement, aligning with studies reporting its protective effects against diet- induced steatosis, likely via polyphenols such as resveratrol and quercetin that enhance lipid oxidation and suppress lipogenesis [54, 55, 56, 57]. Overall, our data suggest that earlier and sustained Rooibos supplementation may offer greater protection against diet-induced hepatic injury, favoring prevention over reversal of established pathology.
Conclusion
In conclusion, Rooibos tea effectively countered key biochemical, immunological, and hepatic alterations induced by a high-carbohydrate diet, likely due to its potent antioxidant, anti-inflammatory, and hepatoprotective properties. With its low cost, safety, and broad spectrum of bioactivity, Rooibos stands out as a promising and accessible adjuvant strategy for the prevention and management of metabolic and liver-related disorders. Future studies optimizing dosage, treatment duration, and delivery methods could further enhance its therapeutic potential.
References
-
Ng M, Fleming T, Robinson M, Thomson B, Graetz N, et al. (2025) Global, regional, and national prevalence of adult overweight and obesity, 1990-2021, with forecasts to 2050: a forecasting study for the Global Burden of Disease Study 2021. Lancet 405(10481): 2025-2038.
-
World Health Organization (2025) Obesity and Overweight. World Health Organization, Switzerland.
-
PalaciosMarin I, Serra D, JimenezChillaron JC, Herrero L, Todorčević M (2023) Childhood obesity: Implications on adipose tissue dynamics and metabolic health. Obesity Reviews 24(8): 13627-13627.
-
Tilg H, Moschen AR (2006) Adipocytokines: mediators linking adipose tissue, inflammation and immunity. Nature Reviews Immunology 6(10): 772-783.
-
Soták M, Clark M, Suur BE, Börgeson E (2024) Inflammation and resolution in obesity. Nature Reviews Endocrinology 20(10): 365-380.
-
Pilkington AC, Paz HA, Wankhade UD (2021) Beige Adipose Tissue Identification and Marker Specificity— Overview. Frontiers in Endocrinology 12: 599134- 599134.
-
Hagberg CE, Spalding KL (2024) White adipocyte dysfunction and obesity-associated pathologies in humans. Nature Reviews Molecular Cell Biology 25(4): 270-289.
-
Xourafa G, Korbmacher M, Roden M (2023) Interorgan crosstalk during development and progression of type 2 diabetes mellitus. Nature Reviews Endocrinology 20(1): 27-49.
-
Wong VWS, Ekstedt M, Wong GLH, Hagström H (2023) Changing epidemiology, global trends and implications for outcomes of NAFLD. Journal of Hepatology 79(3): 612-630.
-
Zhao J, Smith A, Li R (2024) MAFLD as part of systemic metabolic dysregulation. Hepatology International 18(4): 467-479.
-
Tanase DM, Gosset L, Lefranc G (2020) The intricate relationship between type 2 diabetes mellitus (T2DM), insulin resistance (IR), and nonalcoholic fatty liver disease (NAFLD). Journal of Diabetes Research 2020(1): 1-16.
-
He H, Liao S, Zeng Y, Liang L, Chen J, Tao C (2022) Causal relationships between metabolicassociated fatty liver disease and iron status: Twosample Mendelian randomization. Liver International 42(12): 2759-2768.
-
Tong J, Li X, Zhang Y (2022) Targeting a novel inducible GPX4 alternative isoform to alleviate ferroptosis and treat metabolicassociated fatty liver disease. Acta Pharmaceutica Sinica B 12(9): 3650-3666.
-
Lan Y, Chen W, Zhang L (2021) Association between blood copper and nonalcoholic fatty liver disease according to sex. Clinical Nutrition 40(4): 204-2052.
-
Cioffi F, Rossi G, Bianchi A (2022) Altered mitochondrial quality control in rats with metabolic dysfunction- associated fatty liver disease (MAFLD) induced by highfat feeding. Genes 13(2): 315-315.
-
Kazankov K, Jørgensen SMD, Thomsen KL (2018) The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nature Reviews Gastroenterology & Hepatology 16(3): 145-159.
-
Marinović S, Bošnjak B, Šućur V (2023) NKG2D-mediated detection of metabolically stressed hepatocytes by innatelike T cells is essential for initiation of NASH and fibrosis. Science Immunology 8(87): add1599-add1599.
-
Ajuwon OR, Oguntibeju OO, Marnewick JL (2014) Amelioration of lipopolysaccharide-induced liver injury by aqueous rooibos (Aspalathus linearis) extract via inhibition of proinflammatory cytokines and oxidative stress. BMC Complementary and Alternative Medicine 14(1): 392-392.
-
Marnewick J, Joubert E, Joseph S, Swanevelder S, Swart P, et al. (2005) Inhibition of tumour promotion in mouse skin by extracts of rooibos (Aspalathus linearis) and honeybush (Cyclopia intermedia), unique South African herbal teas. Cancer Letters 224(2): 193-202.
-
Wynberg R (2017) Making sense of access and benefit sharing in the rooibos industry: Towards a holistic, just and sustainable framing. South African Journal of Botany 110: 39-51.
-
McKay DL, Blumberg JB (2006) A review of the bioactivity of south African herbal teas: rooibos (Aspalathus linearis) and honeybush (Cyclopia intermedia). Phytotherapy Research 21(1): 1-16.
-
Smith C, Swart A (2018) Aspalathus linearis (Rooibos) - a functional food targeting cardiovascular disease. Food & Function 9(10): 5041-5058.
-
Walters NA, de Villiers E, Joubert E, Beer D (2017) Improved HPLC method for rooibos phenolics targeting changes due to fermentation. Journal of Food Composition and Analysis 55: 20-29.
-
Nehme R, Champagne C (2023) Aspalathus linearis (Rooibos) Targets Adipocytes and ObesityAssociated Inflammation. Nutrients 15(7): 1751-1751.
-
Moosa S, Smith C (2018) Rooibos tea extracts inhibit osteoclast formation and activity through the attenuation of NFκB activity in RAW264.7 murine macrophages. Food & Function 9(6): 3301-3312.
-
Smith SM, Marchion MB, Yates MY, Kumar Y (2013) Aspalathin improves hyperglycemia and glucose intolerance in obese diabetic ob/ob mice. European Journal of Nutrition 52(6): 1599-1605.
-
Orlando P, Richter T (2019) AspalathinRich Green Rooibos Extract Lowers LDLCholesterol and Oxidative Status in HighFat DietInduced Diabetic Vervet Monkeys. Molecules 24(9): 1713-1713.
-
Pretorius L, Smith C (2024) Green rooibos (Aspalathus linearis) promotes gut health: Insight into mechanisms. Journal of Ethnopharmacology 319: 117379-117379.
-
Oliveira MC, Santos A (2013) Acute and sustained inflammation and metabolic dysfunction induced by high refined carbohydratecontaining diet in mice. Obesity 21(5): e32-e40.
-
Folch J, Lees M, Sloane Stanley GH (1957) A simple method for the isolation and purification of total lipides from animal tissues. Journal of Biological Chemistry 226(1): 497-509.
-
Sawicki E, Stanley TW, Pfaff J, D’Amico A (1963) Comparison of fifty-two spectrophotometric methods for the determination of nitrite. Talanta 10(6): 641-655.
-
Triscari J, Nauss-Karol C, Levin BE, Sullivan AC (1985) Changes in lipid metabolism in diet-induced obesity. Metabolism 34(6): 580-587.
-
Sanderson M, Snyman T, et al. (2014) Effects of fermented rooibos (Aspalathus linearis) on adipocyte differentiation. Phytomedicine 21(2): 109-117.
-
Kotzé-Hörstmann LM, Bedada DT, Johnson R, Mabasa L, Sadie-Van Gijsen H (2022) The effects of a green Rooibos (Aspalathus linearis) extract on metabolic parameters and adipose tissue biology in rats fed different obesogenic diets. Food & Function 13(24): 12648-12663.
-
Gianopoulos I, Mantzoros CS, Daskalopoulou SS (2024) Adiponectin and adiponectin receptors in atherosclerosis. Endocrine Reviews 45(4): bnae021- bnae021.
-
Johnson R, de Beer D, V Dludla P, Ferreira D, Muller CJF (2018) Aspalathin from rooibos (Aspalathus linearis): a bioactive C-glucosyl dihydrochalcone with potential to target the metabolic syndrome. Planta Medica 84(12- 13): 926-932.
-
Singla P (2010) Metabolic effects of obesity: a review. World Journal of Diabetes 1(3): 76-76.
-
Jomova K, Vondrakova D, Lawson M, Valko M (2025) Flavonoids and their role in oxidative stress, inflammation, and human diseases. Chemico-Biological Interactions 413: 111489-111489.
-
Lumeng CN, Bodzin JL, Saltiel AR (2007) Obesity induces a phenotypic switch in adipose tissue macrophage polarization. Journal of Clinical Investigation 117(1): 175-184.
-
Skurk T, Alberti-Huber C, Herder C, Hauner H (2007) Relationship between adipocyte size and adipokine expression and secretion. Journal of Clinical Endocrinology & Metabolism 92(3): 1023-1033.
-
Sasaki M, Nishida N, Shimada M (2018) A beneficial role of rooibos in diabetes mellitus: a systematic review and meta-analysis. Molecules 23(4): 839-839.
-
Manne V, Handa P, Kowdley KV (2018) Pathophysiology of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Clinics in Liver Disease 22(1): 23-37.
-
Casagrande BP, Moreira PR (2019) Age-dependent hepatic alterations induced by a high-fat high-fructose diet. Inflammation Research 68(5): 359-368.
-
Ajuwon OR, Katengua-Thamahane E, Van Rooyen J, Oguntibeju OO, Marnewick JL (2013) Protective effects of rooibos (Aspalathus linearis) and/or red palm oil (Elaeis guineensis) supplementation on tert-butyl hydroperoxide-induced oxidative hepatotoxicity in Wistar rats. Evidence-Based Complementary and Alternative Medicine 2013: 1-19.
-
Lawal AO, Akinrinde AS (2019) The cardiovascular protective effects of rooibos (Aspalathus linearis) extract on diesel exhaust particles induced inflammation and oxidative stress involve NF-κB- and Nrf2-dependent pathways modulation. Heliyon 5(3): e01426-e01426.
-
Rahman MM, Gan SH (2017) Cardamom powder supplementation prevents obesity, improves glucose intolerance, inflammation and oxidative stress in liver of high carbohydrate high fat diet induced obese rats. Lipids in Health and Disease 16(1): 1-11.
-
Uličná O, Vančová O, Kucharská J, Janega P, Waczulíková I (2006) Rooibos tea (Aspalathus linearis) partially prevents oxidative stress in streptozotocin-induced diabetic rats. Physiological Research 55(2): 157-164.
-
Uličná O, Vančová O, Kucharská J, Janega P, Waczulíková I (2019) Rooibos tea (Aspalathus linearis) ameliorates the CCl4-induced injury to mitochondrial respiratory function and energy production in rat liver. General Physiology and Biophysics 38(1): 15-25.
-
Pompili S, Baratta F (2020) Long-term abuse of a high- carbohydrate diet is as harmful as a high-fat diet for development and progression of liver injury in a mouse model of NAFLD/NASH. Nutrition 75-76: 110782- 110782.
-
Ishimoto T, Lanaspa MA (2013) High-fat and high- sucrose (Western) diet induces steatohepatitis that is dependent on fructokinase. Hepatology 58(5): 1632- 1643.
-
Yki-Järvinen H (2010) Nutritional modulation of nonalcoholic fatty liver disease and insulin resistance: human data. Current Opinion in Clinical Nutrition and Metabolic Care 13(6): 709-714.
-
Li ZZ, Berk M, McIntyre TM, Feldstein AE (2009) Hepatic lipid partitioning and liver damage in nonalcoholic fatty liver disease: role of stearoyl-CoA desaturase. Journal of Biological Chemistry 284(9): 5637-5644.
-
Lee SB, Kim SJ (2019) Intermittent restraint-induced sympathetic activation attenuates hepatic steatosis and inflammation in a high-fat diet-fed mouse model. American Journal of Physiology-Gastrointestinal and Liver Physiology 317(6): G811-G823.
-
Beltrán-Debón R, Rull A (2011) Continuous administration of polyphenols from aqueous rooibos (Aspalathus linearis) extract ameliorates dietary- induced metabolic disturbances in hyperlipidemic mice. Phytomedicine 18(5): 414-424.
-
Aguirre L, Portillo MP (2014) Effects of resveratrol and other polyphenols in hepatic steatosis. World Journal of Gastroenterology 20(23): 7366-7375.
-
Boqué N, Campión J (2012) Screening of polyphenolic plant extracts for anti-obesity properties in Wistar rats. Journal of the Science of Food and Agriculture 93(5): 1226-1232.
-
Layman JI, Pereira DL, Chellan N, Huisamen B, Kotzé SH (2019) A histomorphometric study on the hepatoprotective effects of a green rooibos extract in a diet-induced obese rat model. Acta Histochemica 121(5): 646-656.
- Investigation of Polymorphisms in PPAR-Ɣ and TRHR Genes and their Impact on Turkish Diabetic and Obese Individuals
- The Impact of Aircraft Noise Exposure on the Efficacy of Empagliflozin Therapy in an Animal Model of Obesity
- Synergistic Effect of Combined Leaf Extract of Vernonia amygdalina, Ocimum gratissimum, and Zingiber officinale Tuber on Phytochemical Profile, Antioxidant Activity, Serum Insulin, and Biochemical Parameters in Streptozotocin-Induced Diabetic Rats
- Investigation of Cardiovascular Responses to Aerobic Exercise in Obese University Students
- A Look at the Phase Angle Obtained by Electrical Bioimpedance
- Metabolic Syndrome and Pre-Metabolic Syndrome among Health Care Workers in Yemen: Prevalence and Associated Risk Factors