Apoptosis Induction of Ganoderic Acid-A by Downregulating miR- 125b and miR-365a-3p in Nalm-6 cells
Acute lymphoblastic leukemia is the most common leukemia in children, which is associated with a high relapse rate despite prevalent therapies. Ganoderic acid-A (GAA) is one of the bioactive compounds of Ganoderma lucidum, which possesses potential antileukemic properties. This study aimed to investigate the effect of the GAA extract on the expression of microRNA and the apoptosis induction in the BCP-ALL cell line. In this case-control study, NALM-6 Cells were treated with the GAA extract and L-asparaginase separately. The cell viability and apoptosis rates were examined using MTT and flow cytometry, respectively. Moreover, the effect of the GAA on the expression of miR-125b and miR-365a-3p, in comparison to L-asparaginase, was studied using RT-PCR. The GAA extract inhibited the cell growth in a dose- and time-dependent manner. Moreover, the GAA extract significantly induced apoptosis compared to L-asparaginase (P=0.0021 vs. P=0.0112). The results showed that the treatment of Nalm-6 cells with GAA significantly reduced the expression of miR-125b (P< 0.0001) and miR-365a-3p (P= 0.0001). Furthermore, it was demonstrated that the downregulation of miR-125b and miR-365a-3p was surprisingly higher in the GA-treated Nalm-6 cells than in L-asparaginase-treated Nalm-6 cells. According to the findings, the GAA extract has antileukemic effects and can be used as a promising agent with fewer side effects in all treatments.
Introduction
B-cell Precursor Acute Lymphoblastic Leukemia (BCP- ALL) is a type of hematologic malignancy in children 2 to 10 years [1, 2]. Although antileukemic therapies such as L-asparaginase lead to complete remission in more than 85% of patients, about half of the cases relapse with an unfavorable prognosis [3, 4]. Moreover, the adverse side- effects of treatments are of great concern for patients and reduce their life expectancy [5]; hence, looking for treatment regimens with further effects and fewer cytotoxic effects is of paramount importance [6]. Ganoderma lucidum (GL) is locally known as Reishi and lingzi and has long been used to prevent and treat various diseases [7, 8, 9, 10]. Medical studies have documented a wide range of biological functions (e.g., antitumor) for GL [11, 12]. One of the main compounds of GL is ganoderic acid (GA) [13]. Recent studies have indicated that GA increases apoptosis in cancer cells by altering miRNA expression [14, 15, 16].
MicroRNAs (miRNA or miR) are single-stranded, non- coding endogenous RNAs having negative regulatory effect on the target gene expression, which reduce the level of the target protein by binding to the 3ʹ-UTR region of mRNA. Investigating the miRNAs expression level is helpful in the diagnosis, prognosis, and treatment of hematologic malignancies [17, 18].
In this regard, miR-125-5p (miR-125b) is one of the first known miRs whose expression has been altered in different cancers; hence, it has been extensively studied [19]. Depending on the type of cancer, it can act as an oncogene or suppressor tumor. In BCP-ALL, AML, and MDS, it induces tumors associated with a dismal prognosis [20, 21, 22].
The miR-365a-3p gene is a newly-found microRNA, whose expression pattern and biological role depend on the type of cancer [23]. In leukemia [24], pancreatic cancer [25], and hepatocellular carcinoma [26], its expression is increased; however, it plays the role of a tumor suppressor in breast cancer [27], melanoma [28], and ovarian cancer [29]. Studies have revealed an increase in the expression of miR-365a-3p and miR-125-5p in patients with ALL [23]. We selected these two microRNAs according to previous studies and with regard to the role of miR-365a-3p and miR- 125-5p in leukemogenesis. By increasing apoptosis rate and changing the miRNAs expression level, GAA seems to have a strong cytotoxic effect on Nalm-6 cells related to the previous treatment of ALL. Accordingly, this study aimed to investigate the effect of treatment with the GAA extract on the apoptosis rate and the expression level of miR-365a-3p and miR-125- 5p compared to L-asparaginase in the Nalm-6 cell line.
Materials and Methods
Reagents
This case-control study was approved by the Ethics Committee of the Shiraz University of Medical Sciences (Code: IR.SUMS.REC.1399.838). The B‐cell precursor leukemia (Nalm‐6) cell line was purchased from the American Type Culture Collection (ATCC; Gaithersburg, Maryland, USA). All cell culture materials and reagents were purchased from the Gibco Life Technologies (Waltham, MA) and Sigma‐Aldrich (Munich, Germany). The GAA extract was obtained from the Sigma Aldrich Company (St. Louis, MO, USA) and was prepared in DMSO. L-Asparaginase powder vial 10000 IU (Zydus Cadila Company, India) was prepared in PBS.
Cell Culture
NALM-6 was cultured in the RPMI-1640 medium supplemented with 10% fetal bovine serum and 100 U/ml penicillin-streptomycin and 2mM glutamine in a humidified 5% CO2 incubator at 37°C under standard cell culture conditions. The culture medium was changed according to the standard techniques, and the cells were passaged when they achieved 80% confluency. The Trypan blue exclusion method was used to assess cell viability.
MTT Assay
The antiproliferative effects of the GAA extract on the B-ALL cell line, Nalm-6, were first evaluated. In short, 2×10^4 cells/well in 150μL growth medium were seeded in 96-well plates (cells in the logarithmic growth phase). The cells were treated with appropriate amounts of the GAA extract stock solution to reach the concentrations of 25, 50, 100, 200, and 400 μg/ml. The untreated cells were defined as the control group. After the treatment, the cells were incubated under cell culture conditions for 24, 48, and 72 hours. Then the MTT solution (5 mg/ml) was added and incubated for an additional 4 hours. After incubation, supernatants were removed, and the remaining water-insoluble formazan crystals were dissolved in 150 μl dimethyl sulfoxide (DMSO) for 10 min by shaking gently. Finally, the optical density was measured at 570 nm wavelength using a Stat Fax 2100 microplate photometer (Stat Fax 2100, SKU: 8036-10-0020, USA). IC50 was calculated as the concentration of compounds, causing a 50% inhibition of cell viability. At least three independent experiments were performed in quadruplicate. DMSO was used to prepare GAA and bring it to the desired concentrations. Given the consecutive dilutions of the initial stock, the DMSO concentration was <0.01; hence, there was no need to control DMSO.
Flow Cytometry
To determine the early and late apoptotic populations in the Nalm6 cell-line induced by the GAA treatment, we performed flow cytometry using Annexin-V/PI double staining kit (BD Biosciences, San Jose, CA). Moreover, the standard L-asparaginase treatment was used to compare the effect of GAA on Nalm-6 cells. To perform this test, the Nalm-6 cells in the logarithmic phase were inoculated into 24-well plates and treated with GAA and L-asparaginase
separately for 48h. Then the treated cells were collected after centrifugation at 3000 rpm for 10 min and were washed twice with PBS. Afterward, the cells were resuspended in 100 μl of a binding buffer, and 5 μl of Annexin-V was then added to the cell suspension incubated for 15 min at room temperature in dark. The cells were rinsed and resuspended in 200 μl of the binding buffer. In the next ohase, 5 μl of PI was added before reading the values by flow cytometry. Annexin-V positive and PI-negative cells were considered to be in the early apoptotic phase, and the cells with Annexin-V and PI-positive were considered to undergo late apoptosis. The reaction of Annexin V-FITC and PI binding was analyzed quantitatively by using a FACScalibur flow cytometer (BD Biosciences) and FlowJo software (TreeStar LLC).
RNA Extraction
Total RNA was isolated 48 h after the treatment with GAA extract and L-Asparaginase using TRIzol reagent (Thermo Fisher Scientific, MA). RNA quality was assessed with regard to 260/280 and 260/230 ratios by NanoDrop spectrophotometer (Thermo Scientific NanoDrop2000, Finland), and the extracted RNA was stored at -80 ˚C.
cDNA Synthesis and Real-Time PCR
According to the manufacturer’s protocol, specific cDNAs of the selected miRNA were synthesized by using Parsgenom RTreagent Kit (Pars genome, Iran). First, the poly-A tail was added to miRNAs with polyA polymerase at 37 °C. Then RT- enzyme, reaction buffer, and miR-specific primers for cDNA synthesis were mixed with RNA polyA tail (Table 1). Finally, they were incubated at 45 °C for 60 min and inactivated at 85 ºC for 1 min. Subsequently, Real Time-PCR was performed to analyze the cDNAs using Parsgenome Kit.
| miRNAs | Sequence |
| miR-125b | 5' UCCCUGAGACCCUAACUUGUGA 3' |
| miR-365a-3P | 5' UAAUGCCCCUAAAAAUCCUUAU3' |
Table 1: The primer sequences used for real-time polymerase chain reaction assay.
Real Time-PCR Assay
To evaluate the expression of miR-125b and miR-365a-3p genes, we calculated the quantitative real-time polymerase chain reaction (qRT-PCR) in a Rotor-Gene Q system (Qiagen, Hilden, USA). The qRT-PCR was run under the following thermal cycling conditions: initial denaturation at 95 ˚C for
15 min followed by 45 cycles of 95˚C for 30 sec, 63˚C for 30 sec, and 72 ˚C for 30 sec. All the qRT-PCR reactions were performed in triplicate. A melting curve analysis was applied to verify the specificity of the products, and the values of the relative quantification were calculated based on the 2^-ΔΔCt relative expression formula. Additionally, the U6 rRNA gene was used as an internal control.
Statistical Analysis
For the primary selection of miRNAs to be included in this study, in addition to some information extracted from past studies, the characteristics for the selected miRNAs, including target genes, the underlying cell process, the expression or non-expression of miR in cancerous cells, the function of the selected miRNAs as oncomiR, were evaluated by programs such as Targetscan, miRanda, and MiRTARbase.
All experiments were repeated at least three times and were expressed as mean ± SEM. Statistical significance was assessed by GraphPad Prism 8.0.2 (GraphPad Software, Inc., La Jolla, CA, USA) using the SPSS software version 19.0 (IBM Corp.). The student’s t-test analyzed the difference between the two groups. The significance level was set to be P <0.05.
Results
Effect of GAA on Cell Viability
The cytotoxic effects of different concentrations of the GAA extract (25, 50, 100, 200, and 400 μg/ml) on NALM-6 cells were assessed using the MTT assay after three different time points, namely 24, 48, and 72h. According to the results, GAA could inhibit the growth of NALM-6 cells both in time‐ and dose‐dependent manners. We determined the optimal concentration of GAA by investigating its IC‐50. In this study, the GraphPad software was used to calculate IC50. The IC50 of the GAA extract in NALM-6 cell lines after 48 h of incubation was approximately 130 μg/ml. Accordingly, we used 130 μg/ ml of GAA after 48 hours for all subsequent experiments. The data are not presented for 24 and 72h. Induction of Apoptosis in BCP-ALL Cell-Lines by GAA Extract Nalm6 cells were incubated with 130 μg/ml GAA for 48 hours. As shown in Figure 2-A, GAA illustrated a significant percentage of early and late‐stage apoptotic cells after 48-hour treatment that was 40.3%. According to Figure 2-B, the percentage of apoptosis of GAA-treated cells was impressively increased compared to untreated cells.
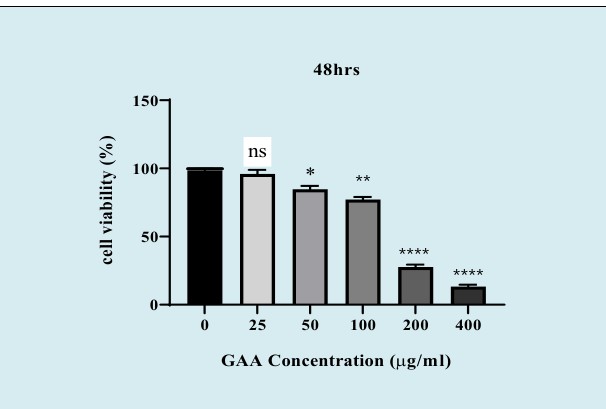
Figure 1: The cytotoxic effects of GAA extract on NALM-6 cell lines. The cells were treated with Increasing concentrations of GAA (25, 50, 100, 200 and 400 μg/ml) and their cell viability were measured after 48h incubation, using MTT assay. The percentage of cell viability was significantly decreased compared with the untreated cell line in a dose-dependent manner. Values are given as mean ±SEM of three independent experiments. (N.S: nonsignificant, *P < 0.05, **P < 0.01, and ⃰ ⃰ ⃰ ⃰ P<0.0001). MTT; 3‐ (4, 5‐dimethylthiazol‐2‐yl) ‐2, 5‐diphenyl‐tetrazolium bromide.
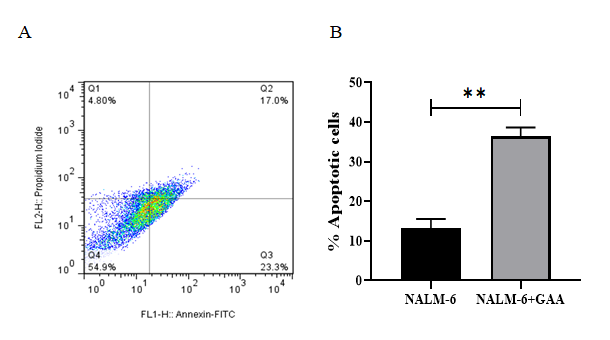
Figure 2: Results from the induction of apoptosis by the GAA extract on NALM-6 cell line. The NALM-6 cells were exposed to intended concentration of the GAA extract and then the induction of apoptosis after 48 h incubation was evaluated. (A) Flow cytometry images, Q1, Q2, Q3 and Q4 represent necrotic cells, late apoptosis, early apoptosis and live cells, respectively. (B) The apoptotic cells percentage was increased in GAA treatment compared with untreated cells (P=0.0021). The error bars in the diagram represent the standard deviations of two separate tests.
Comparing Apoptosis in Cells Treated With GAA Extract and L-Asparaginase
Nalm6 cells were incubated with 8 μM L-Asparaginase for 48h. As shown in Figure 3, the percentage of apoptosis in the NALM-6 cells treated with L-Asparaginase (L-ASP) was approximately 33.4%. In contrast, the apoptosis rate in the GAA-treated cells was 40.3%. Compared with the L-ASP treated cells, more extensive apoptosis was observed for the cells treated with GAA (130 μg/mL) after 48h, indicating the lower cytotoxic effect of L-Asparaginase in acute lymphoblastic leukemia cell-lines relative to GAA.
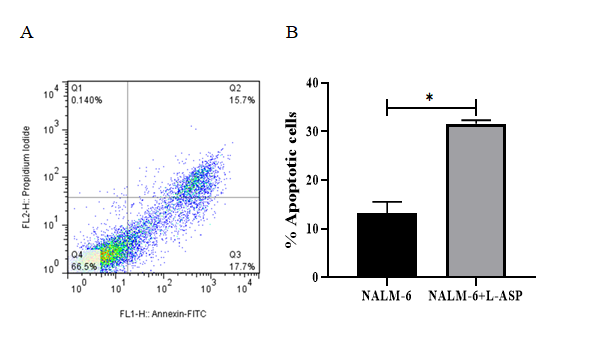
Figure 3: Results from the induction of apoptosis by the L-Asparginase on NALM-6 cell line. The cells were exposed to 8 μM of the L-Asparginase and then the induction of apoptosis after 48 h incubation was evaluated. (A) Flow cytometry images, Q1, Q2, Q3 and Q4 represent necrotic cells, late apoptosis, early apoptosis and live cells, respectively. (B) The apoptotic cells percentage was increased in L-ASP treatment compared with untreated cells (P=0.0112). The error bars in the diagram represent the standard deviations of two separate tests.
Effect of GAA on the Expression of Mir‐125b and Mir‐365a-3p
Following the real‐time PCR results, miR‐125b (median fold‐change of expression [FC] = 0.135, P< 0.0001) and miR‐365a-3p ([FC] = 0.252, P = 0.0001) significantly decreased in the GAA-treated Nalm6 cells compared with untreated Nalm6 cells. Moreover, miR‐125b (median fold‐ change of expression [FC] = 0.206, P=0.0023) and miR‐365a- 3p ([FC] = 0.514, P = 0.0459) significantly decreased in L-asparginase-treated Nalm6 cells compared with untreated Nalm6 cells (Figures 4, 5). On the other hand, the expressions of miR‐125b and miR‐365a-3p significantly reduced in the GAA-treated Nalm6 cells compared to the L-asparginase- treated Nalm6 cells. The results are presented as mean ± SD for the three independent experiments in triplicate.
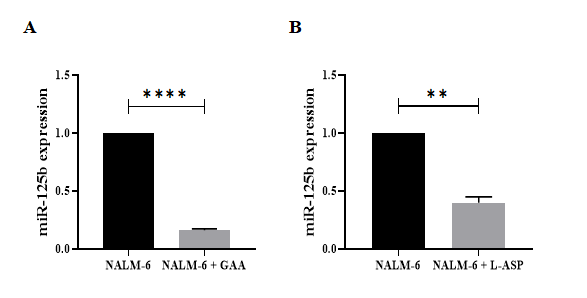
Figure 4: The expression levels of miR‐125b in GAA‐ and L-ASP-treated Nalm6 cell line. Significant differences of alteration in the expression levels of miR‐125b were measured between untreated and treated Nalm6 cells by quantitative RT‐PCR. (A) miR-125b (p < 0.0001) were confirmed down-regulated in GAA treatment compared with untreated cells; also, (B) the expression levels of miR-125b (p=0.0023) was decreased in L-ASP treatment compared with untreated cells. Interestingly, results showed that the expression levels of miR- 125b reduced by GAA treatment compared to L-ASP treatment. U6 used as internal control. (⃰ ⃰ P<0.01,⃰ ⃰ ⃰ ⃰ P<0.0001). RT PCR; real-time polymerase chain reaction.
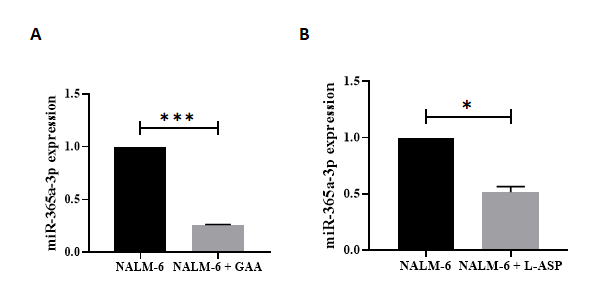
Figure 5: The expression levels of miR‐365a-3p in GAA‐ and L-ASP-treated Nalm6 cell line. Significant differences of alteration in the expression levels of miR‐365a-3p were measured between untreated and treated Nalm6 cells by quantitative RT‐PCR. (A) miR‐365a-3p (p = 0.0001) were confirmed down-regulated in GAA treatment compared with untreated cells; also, (B) the expression levels of miR‐365a-3p (p=0.0495) was decreased in L-ASP treatment compared with untreated cells. Interestingly, results showed that the expression levels of miR‐365a-3p reduced by GAA treatment compared to L-ASP treatment. U6 used as internal control. ( ⃰ P < 0.05, ⃰ ⃰ ⃰ P<0.001). RT PCR; real-time polymerase chain reaction.
Discussion
In recent years, remarkable progress has been observed in treating acute lymphoblastic leukemia, which has led to substantial outcomes and improvements for the patients. Nonetheless, the exclusive toxicities resulting from antileukemic therapeutic regimens with no early intervention are life-threatening. Accordingly, this study highlights the advances in novel therapies for acute lymphoblastic leukemia and supportive supplements. Recently, herbal compounds have been considered for their anti-cancer properties [30, 31].
The GAA extract is a triterpenoid compound from Ganoderma lucidum, having impressive antitumor activities in various cancer cells [32]. The present study revealed the promising antileukemic impacts on B cell precursor-acute lymphoblastic leukemia cells applied by the GAA extract. Much experimental evidence previously investigated different biological activities of this herbal extract, including the antioxidant effect, apoptosis induction, and metastasis inhibition with low toxicity or no side effects [14, 16]. However, this is the first report exhibiting the antiproliferative impact of GAA in comparison to the antileukemic agent, L-asparaginase, and its effect on the expression of miR-365a- 3p and miR-125-5p in BCP-ALL cell-lines.
The present findings suggest that the GAA extract suppress the growth of Nalm-6 cells and reduces cell viability (Figure1). For this purpose, five different concentrations of the GAA extract (25,50,100,200 and 400 μg/ml) were selected based on the previous studies and their effects on Nalm-6 cells at three different times (namely 24, 48, and 72 hours) were evaluated. It was found that GAA plays a role in reducing cell viability both in time‐ and dose‐dependent manners. Due to the low rate of cell viability after 24h and its high rate after 72h, the subsequent experiments were performed 48h later. The IC50 of the GAA extract was determined to be 130 μg/ml. Moreover, in vitro treatment with 130 μg / ml, GAA effectively reduces the viability of Nalm-6 cells after 48 hours. Therefore, GAA maybe exerts a cytotoxic activity by inducing apoptosis in the BCP-ALL cells.
Apoptosis is a process involved in cell death, and many researchers have considered this process as an underlying mechanism in preventing the proliferation of malignant tumor cells [33]. Numerous studies have examined the effect of the GL ingredients on the apoptosis induction in the acute lymphoblastic leukemia cell lines [9, 34]. In this study, among other components of Ganoderma lucidum, we selected GA as a potential factor inhibiting leukemic cell proliferation. The results indicated that the apoptosis rate of Nalm-6 cells after being exposed to GAA was about 40.3% (Figure 2A); however, the percentage of apoptosis in the L-asparaginase- treated cells was 33.4% (Figure 3A). L-ASP is a substantial component of chemotherapeutic regimens. However, the side- effects attributed to this drug have affected its therapeutic outcomes. Virtually, the findings illustrate a noticeable difference between the apoptosis rates in the GAA-treated and L-ASP-treated cells, implying that the GAA extracts significantly suppress cell proliferation and induce apoptosis compared to L-asparaginase. A study examined apoptosis induced by plant compounds, including cinnamon, ginger, and green tea in the Nalm-6 cells. It showed that cinnamon and green tea did not significantly differ in the apoptosis rate compared to the untreated cells. Ginger-induced apoptosis on Nalm-6 cells was about 37% [35]. Compared to this study, our findings showed that the percentage of GAA-induced apoptosis in Nalm-6 cells was higher than the percentage of ginger apoptosis, even though the amount of ginger was higher than GAA. Since the role of GAA in apoptosis induction and mechanism of action as well as its actual target in BCP- ALL are unexplored. Further experiments are required to determine the mechanism by which GAA exerts its inhibitory effect on BCP-ALL cells via apoptosis induction.
We predicted that the apoptosis induction in Nalm- 6 cells by GAA might be due to the altered expression of miRNAs (miRNA or miR). In this study, the expressions of miR-125-5p and miR-365a-3p in cells treated to GAA and L-asparaginase were compared with the untreated Nalm-6 cells for the first time. Interestingly, the exposure of Nalm-6 cells to GAA significantly reduced miR-125-5p and miR-365a-3p expression. According to the existing evidence, miRNAs regulates various cellular processes, the most important of which are apoptosis, proliferation, and differentiation [19]. Over the last decade, researchers have conducted numerous studies on the effect of miRNAs in the diagnosis and follow-ups, assessed the risk of leukemia relapse, and suggested that miRNAs can be used as a predictive biomarker [36].
It is worth noting that miR-125-5p (miR-125b) is one of the first detected miRNAs, including two subtypes of miR- 125b-1 and miR-125b-2. MiR-125b-1 is involved in several common chromosomal abnormalities, including t (11; 14) leading to BCP-ALL or AML, and miR-125b-2 is elevated in trisomy 21 patients associated with AML7 [37]. Moreover, miR-125b acts as an oncomir or tumor suppressor in malignant cells [38]. Many studies have demonstrated that the expression impairment of miR-125-5p is markedly related to the reduction of apoptosis and promotion of cell proliferation in hematological malignancies [39]. Although the possible mechanism of miR-125b in the induction of tumor genesis has not yet been detected, several potential mechanisms may explain the effects of miR-125-5p dysregulation on cellular processes in leukemic cells: (1) Promoting cell proliferation via PI3K/AKT/ERK signaling pathway [40], (2) Blocking the differentiation of pre-B by targeting MAP3K11 activity, (3) Reducing cell death by targeting p53 tumor suppressor function with decreased mRNA and protein levels , and (4) Inhibiting apoptosis by increasing impressive anti-apoptosis factors Bcl-2 and c-myc and decreasing pro-apoptosis factors BAK1 and Caspase-3 [40]. A study also exhibited that the high levels of miR-125-5p expression were associated with increased relapse and death in acute myeloblastic leukemia patients [41]. The regulation of miR-125-5p expression seems to be one of the therapeutic targets. In our study, GAA downregulated the expression of miR-125-5p in Nalm-6 cells compared to the control cells, as determined by qRT-PCR (Figure 4.A). However, the L-asparaginase had less effect on reducing miR-125b expression (Figure 4.B) (median fold‐change of expression [FC] = 0.135, P<0.0001 vs [FC] = 0.206, P=0.0023 for GAA and L-ASP, respectively). MiR-125b, which plays a critical role in cancer progression, may be a key target of GAA for its chemopreventive activity in ALL. To the best of our knowledge, this is the first study evaluating the expression of miR-125-5p in Nalm-6 cells treated with GAA. The observations in a study on human leukemia cells demonstrated that GL induced mitochondrial Permeability and apoptosis mediated by Bcl-2 down- regulation, Bax translocation, mitochondrial cytochrome c-release, and caspase-3 activation [41]. It was speculated that miR-125 might be associated with the increased expression of anti-apoptotic protein Bcl-2 in pre-B-cell acute lymphoblastic leukemia. Accordingly, GAA can inhibit Bcl-2 by downregulating oncogenic miR-125b. This is, while the mechanisms involved in the regulation of this miRNA have not yet been elucidated.
Interestingly, similar to miR-125b, a decrease in the miR- 365a-3p expression was observed in the acute lymphoblastic leukemia cell lines. In this study, the expression of miR-365a- 3p in GAA‐treated and L-Asparaginase-treated Nalm6 cell lines was followed by quantitative RT‐PCR. Our findings indicated that the expression of the miR-365a-3p decreased after treating the Nalm-6 cells with GAA extract compared to untreated Nalm-6 cells (Figure 5.A). When the Nalm- 6 cells are treated with L-asparaginase, they express less downregulation than those treated with GAA (Figure 5.B) ([FC] = 0.252, P = 0.0001 vs [FC] = 0.514, P = 0.0459 for GAA and L-ASP, respectively). Previous studies have documented that the miR-365 expression increased in several malignant tumor cells [42]. A study revealed the higher levels of miR- 365 in exosomes derived from imatinib-resistant CML cells compared to sensitive cells, which was caused by inhibiting the expression of pro-apoptotic protein BAX and cleaved Caspase-3 in the sensitive CML cells [43]. In another study, proapoptotic BCL2 associated X (BAX) was one of the downstream targets of onco-miR-365, which up regulation miR-365 suppressed BAX protein level in cells of cutaneous squamous cell carcinoma (CSCC) which downregulation BAX finally caused tumor cells resistant to apoptosis and CSCC expansion [44]. These findings were consistent with those of the present study, suggesting that miR-365 acts as an oncogene as such miR-365 may promote tumor cells by suppressing the apoptotic pathway (Figure 6).
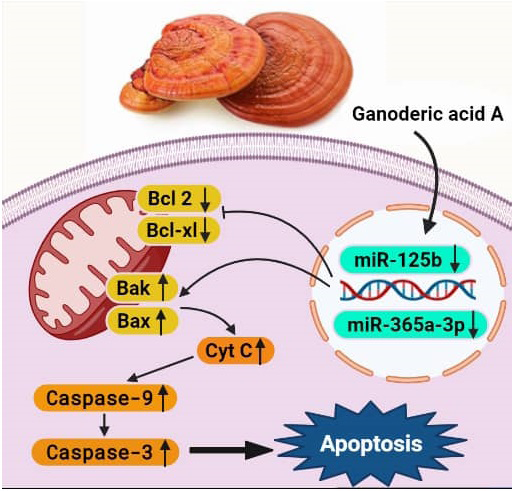
Furthermore, in some cancer cells, miR-365 plays the role of a tumor suppressor. Since our study was the first study evaluating the miR-365 expression and the inhibitory effect of GAA on the microRNA expression in Nalm-6 cells, the action mechanism of GAA in reducing miR-365 expression is not yet known.
Conclusion
The present study illustrates the strong cytotoxic effect of the GAA on acute lymphoblastic leukemia cell lines caused by inducing apoptosis. Moreover, the findings also indicate that GAA alters the expression of miR-125-5p and miR-365a- 3p. The findings confirm the similar effect of L-Asparaginase on the Nalm-6 cell line. Accordingly, the GAA can be used as a combination with fewer side effects in treating acute lymphoblastic leukemia. However, further studies are recommended to delve into this issue.
References
-
Allemani C, Weir HK, Carreira H, Harewood R, Spika D, et al. (2015) Global surveillance of cancer survival 1995- 2009: analysis of individual data for 25,676,887 patients from 279 population-based registries in 67 countries (CONCORD-2). Lancet 385(9972): 977-1010.
-
Blackburn LM, Bender S, Brown S (2019) Acute Leukemia: Diagnosis and Treatment. Semin Oncol Nurs 35(6): 150950.
-
Bhojwani D, Pui CH (2013) Relapsed childhood acute lymphoblastic leukaemia. Lancet Oncol 14(6): e205-e217.
-
Stein AS, Schiller G, Benjamin R, Jia C, Zhang A, et al. (2019) Neurologic adverse events in patients with relapsed/refractory acute lymphoblastic leukemia treated with blinatumomab: management and mitigating factors. Ann Hematol 98(1): 159-167.
-
Pui CH (2020) Precision medicine in acute lymphoblastic leukemia. Front Med 14(6): 689-700.
-
Minaeeian G, Ranjbaran R, Tandel P, Zare F and Tamaddon G (2021) Effects of L-asparaginase on The Expression of EBF1 Gene, miR-181b, and miR-17 and Its Association with Apoptosis in the Nalm-6 Cell Line. International Journal of Medical Research & Health Sciences 10(11): 88-96.
-
Müller CI, Kumagai T, O’Kelly J, Seeram NP, Heber D, et al. (2006) Ganoderma lucidum causes apoptosis in leukemia, lymphoma and multiple myeloma cells. Leuk Res 30(7): 841-848.
-
Babu PD, Subhasree RS (2008) The sacred mushroom Reishi-a review. American-Eurasian Journal of Botany 1(3): 107-110.
-
Li P, Liu L, Huang S, Zhang Y, Xu J, et al. (2020) Anti-cancer Effects of a Neutral Triterpene Fraction from Ganoderma lucidum and its Active Constituents on SW620 Human Colorectal Cancer Cells. Anticancer Agents Med Chem 20(2): 237-244.
-
Sohretoglu D, Huang S (2018) Ganoderma lucidum Polysaccharides as An Anti-cancer Agent. Anticancer Agents Med Chem 18(5): 667-674.
-
Liang C, Tian D, Liu Y, Li H, Zhu J, et al. (2019) Review of the molecular mechanisms of Ganoderma lucidum triterpenoids: Ganoderic acids A, C2, D, F, DM, X and Y. Eur J Med Chem 174: 130-141.
-
Gill BS, Kumar S, Navgeet (2016) Evaluating anti-oxidant potential of ganoderic acid A in STAT 3 pathway in prostate cancer. Mol Biol Rep 43(12): 1411-1422.
-
Yao X, Li G, Xu H, Lü C (2012) Inhibition of the JAK- STAT3 signaling pathway by ganoderic acid A enhances chemosensitivity of HepG2 cells to cisplatin. Planta Med 78(16): 1740-1748.
-
Wu QP, Xie YZ, Deng Z, Li XM, Yang W, et al. (2012) Ergosterol peroxide isolated from Ganoderma lucidum abolishes microRNA miR-378-mediated tumor cells on chemoresistance. PLoS One 7(8): e44579.
-
Tandel P, Sharifiyazdi H, Farzadfard E, Zareifar S, Mohammadinezhad S, et al. (2021) Changes in Expression of miR-128, miR-144-3p, miR-181b, and miR-451 in Response to Treatment of Pediatric Acute Lymphoblastic Leukemia (B-ALL). Journal of Isfahan Medical School 38(603): 914-920.
-
Farzadfard E, Kalantari T, Tamaddon G (2020) Serum Expression of Seven MicroRNAs in Chronic Lymphocytic Leukemia Patients. J Blood Med 11: 97-102.
-
Ranjbar R, Karimian A, Aghaie Fard A, Tourani M, Majidinia M, et al. (2019) The importance of miRNAs and epigenetics in acute lymphoblastic leukemia prognosis. Journal of cellular physiology 234(4): 3216-3230.
-
Bousquet M, Harris MH, Zhou B, Lodish HF (2010) MicroRNA miR-125b causes leukemia. Proc Natl Acad Sci U S A 107 (50): 21558-21563.
-
So AY, Sookram R, Chaudhuri AA, Minisandram A, Cheng D, Xie C, et al. (2014) Dual mechanisms by which miR- 125b represses IRF4 to induce myeloid and B-cell leukemias. Blood 124(9): 1502-1512.
-
Swellam M, Hashim M, Mahmoud MS, Ramadan A, Hassan NM (2018) Aberrant Expression of Some Circulating miRNAs in Childhood Acute Lymphoblastic Leukemia. Biochem Genet 56(4): 283-294.
-
Hamada S, Masamune A, Miura S, Satoh K, Shimosegawa T (2014) MiR-365 induces gemcitabine resistance in pancreatic cancer cells by targeting the adaptor protein SHC1 and pro-apoptotic regulator BAX. Cell Signal 26(2): 179-185.
-
Jin Y, Wong YS, Goh BK, Chan CY, Cheow PC, et al. (2019) Circulating microRNAs as Potential Diagnostic and Prognostic Biomarkers in Hepatocellular Carcinoma. Sci Rep 9(1): 10464.
-
Liu F, Zhuang L, Wu R, Li D (2019) miR-365 inhibits cell invasion and migration of triple negative breast cancer through ADAM10. J BUON 24(5): 1905-1912.
-
Zhu Y, Wen X, Zhao P (2018) MicroRNA-365 Inhibits Cell Growth and Promotes Apoptosis in Melanoma by Targeting BCL2 and Cyclin D1 (CCND1). Med Sci Monit. 24: 3679-3692.
-
Wang Y, Xu C, Wang Y, Zhang X (2017) MicroRNA-365 inhibits ovarian cancer progression by targeting Wnt5a. American journal of cancer research 7(5): 1096-1106.
-
Khazir J, Riley DL, Pilcher LA, De-Maayer P, Mir BA (2014) Anticancer agents from diverse natural sources. Nat Prod Commun 9(11): 1655-1669.
-
Sun W, Shahrajabian MH, Cheng Q (2020) Traditional Iranian and Arabic herbal medicines as natural anti- cancer drugs. Agrociencia 54(1): 129-142.
-
Yang Y, Zhou H, Liu W, Wu J, Yue X, et al. (2018) Ganoderic acid A exerts antitumor activity against MDA-MB-231 human breast cancer cells by inhibiting the Janus kinase 2/signal transducer and activator of transcription 3 signaling pathway. Oncol Lett 16(5): 6515-6521.
-
Xu X, Lai Y, Hua ZC (2019) Apoptosis and apoptotic body: disease message and therapeutic target potentials. Bioscience reports 39(1).
-
Radwan FF, Hossain A, God JM, Leaphart N, Elvington M, et al. (2015) Reduction of myeloid-derived suppressor cells and lymphoma growth by a natural triterpenoid. J Cell Biochem 116(1): 102-114.
-
Rahimi Babasheikhali S, Rahgozar S, Mohammadi M (2019) Ginger extract has anti-leukemia and anti-drug resistant effects on malignant cells. J Cancer Res Clin Oncol 145(8): 1987-1998.
-
Hosseinahli N, Aghapour M, Duijf PHG, Baradaran B (2018) Treating cancer with microRNA replacement therapy: A literature review. J Cell Physiol 233(8): 5574- 5588.
-
Chapiro E, Russell LJ, Struski S, Cave H, Radford-Weiss I, et al. (2010) A new recurrent translocation t (11; 14) (q24; q32) involving IGH@ and miR-125b-1 in B-cell progenitor acute lymphoblastic leukemia. Leukemia 24(7): 1362-1364.
-
Vafadar A, Mokaram P, Erfani M, Yousefi Z, Farhadi A, et al. (2019) The effect of decitabine on the expression and methylation of the PPP1CA, BTG2, and PTEN in association with changes in miR-125b, miR-17, and miR- 181b in NALM6 cell line. J Cell Biochem 120(8): 13156- 13167.
-
Jiang Y, Luan Y, Chang H, Chen G (2018) The diagnostic and prognostic value of plasma microRNA-125b-5p in patients with multiple myeloma. Oncol Lett 16(3): 4001- 4007.
-
Liu Z, Smith KR, Khong HT, Huang J, Ahn EY, et al. (2016) miR-125b regulates differentiation and metabolic reprogramming of T cell acute lymphoblastic leukemia by directly targeting A20. Oncotarget 7 (48): 78667- 78679.
-
Zhang Y, Zeng C, Lu S, Qin T, Yang L, Chen S, et al. (2016) Identification of miR-125b targets involved in acute promyelocytic leukemia cell proliferation. Biochem Biophys Res Commun 478(4): 1758-1763.
-
Knackmuss U, Lindner SE, Aneichyk T, Kotkamp B, Knust Z, et al. (2016) MAP3K11 is a tumor suppressor targeted by the oncomiR miR-125b in early B cells. Cell Death Differ 23(2): 242-252.
-
Murray MY, Rushworth SA, Zaitseva L, Bowles KM, Macewan DJ (2013) Attenuation of dexamethasone- induced cell death in multiple myeloma is mediated by miR-125b expression. Cell Cycle 12(13): 2144-2153.
-
Li Q, Wu Y, Zhang Y, Sun H, Lu Z, et al. (2016) miR-125b regulates cell progression in chronic myeloid leukemia via targeting BAK1. Am J Transl Res 8(2): 447-459.
-
Jiang L, Deng T, Wang D, Xiao Y (2018) Elevated Serum Exosomal miR-125b Level as a Potential Marker for Poor Prognosis in Intermediate-Risk Acute Myeloid Leukemia. Acta Haematol 140(3): 183-192.
-
Lyu J, Zhao L, Wang F, Ji J, Cao Z, et al. (2019) Discovery and Validation of Serum MicroRNAs as Early Diagnostic Biomarkers for Prostate Cancer in Chinese Population. Biomed Res Int 2019: 9306803.
-
Min QH, Wang XZ, Zhang J, Chen QG, Li SQ, et al. (2018) Exosomes derived from imatinib-resistant chronic myeloid leukemia cells mediate a horizontal transfer of drug-resistant trait by delivering miR-365. Exp Cell Res 362(2): 386-393.
-
Zhou L, Gao R, Wang Y, Zhou M, Ding Z (2017) Loss of BAX by miR-365 Promotes Cutaneous Squamous Cell Carcinoma Progression by Suppressing Apoptosis. Int J Mol Sci 18(6): 1157.
- How to Identify and Overcome Barriers in Developing Blood Systems?
- Why Was Transfusion Medicine Not Recognized as a Clinical Discipline?
- Outcomes of Lenalidomide Relapsed/Refractory Patients
- Is Transfusion Always Necessary?
- The Logistics of Production and Use of Blood and Blood Components
- The Challenge for Component Therapies