Extracellular Vesicles in Hematological Malignancies: Communicators for Resistance and Treatment
Extracellular vehicles (EVs), the secret messengers of a cell, are cell-derived signaling vesicles present in most human fluids. They have been recently discovered and are known as signaling particles or a type of dialogue between cells known as microvesicles, exosomes, and apoptotic bodies. The EVs can carry different types of cargo that can be delivered to recipient cells and lead to changes in the phenotype of a cell which might be useful or harmful. Here, we discuss what these busybody cells are, how they are formed, and what they can do in drug resistance or how they have been used as a therapeutic tool in several hematological malignancies. These miniature molecules are a fascinating new field of research. They can be made artificially as therapeutic agents in the era of personalized medicine for overcoming the relapse of hematological malignancies
Introduction
Hematological malignancies are heterogeneous diseases involving blood and bone marrow and could also be organ- associated diseases that consistently show drug resistance and recurrence. These malignancies are responsible for 1.2 million new disease cases annually worldwide [1]. Today, hematological malignancies are treated with a range of medications as well as drug combinations according to the guidelines of the World Health Organization (WHO), including chemotherapy, immunotherapy, targeted therapy, and cell cycle inhibitors. Despite all the strategies and therapeutic advances in hematological malignancies in recent decades, cancer drug resistance is a major and significant obstacle in treatment management. Research on these major therapeutic obstacles has yielded extensive results. Some of the factors and mechanisms involved in therapeutic resistance include microRNAs, extracellular vesicles (EVs), and miRNA-package vesicles [2]. In this review, EVs, their biogenesis, and their importance in drug resistance in a set of hematological malignancies, such as multiple myeloma, chronic myelogenous leukemia (CML), and acute myelogenous leukemia (AML), and chronic lymphocytic leukemia (CLL), are reviewed. In hematological malignancies, the EVs of blood-derived tumors regulate the microenvironment, affect the immune cells, transmit tolerance signals, stop cell differentiation, and induce apoptosis in CD8 cytotoxic T-cells induce regulatory T-cells, promote disease development, and cause drug resistance. Awareness of these EVs leads to new hope for developing targeted therapy for hematological malignancies.
Extracellular Vesicles
The EVs are locked structures decorated with phospholipid bilayer membranes consisting of four sets of microvesicles, exosomes, apoptotic bodies, and oncosomes [3, 4]. The movements of EVs between cells act as cell-to-cell communication and convey a wide range of biomolecules embracing mRNAs, miRNAs, proteins, chaperones, lipids, and markers that indicate the birthplace cells. The changes induced by them can cause physiological or pathological conditions. Microvesicles are small membrane-bound vesicles usually measuring 100-1000 nm and being shed from a cell surface. On the other hand, exosome vesicles are more homogeneous and are smaller than microvesicles, with a size of 30-120 nm and originating from multivesicular bodies (MVBs). Recent research has shown that these EVs may cause epigenetic changes in the target cell by transferring products, such as the source cell receptor to the target cell, oncogenic products, transcription factors, or pathogens, as well as mRNA and miRNA to the target cell and change its function [5, 6].
Biogenesis
Although the basis for the formation of these submicron vesicles has been proven to some extent, there is a long way to understand this process entirely. In this section, the formation mechanisms of microvesicles and exosomes are briefly described. Microvesicles are directly formed by the budding of the plasma membrane [7], with changes in the cytoskeletal proteins and phospholipids symmetry playing an essential role. Phospholipid asymmetry of membrane layers is maintained by translocase activity. Some factors, such as complement-mediated lysis, stress, and oxidative injury, can increase intracellular calcium that activates calpain, gelsolin, and scramblase whilst inactivating the translocase enzyme. Gelsolin and calpain activate scramblase and cause some cuts in actin filaments, promoting membrane asymmetry and phosphatidylserine externalization. On the other hand, ADP-ribosylation factor 6 (ARF6) triggers a cascade signal that activates phospholipase D (PLD). The activated phospholipase invokes an ERK kinase enzyme into the plasma membrane, which is activated by phosphorylating the light chain myosin. The combination of these pathways causes the release and formation of microvesicles. Exosomes are formed along the path of the endosomal network. Early endosomes combine with endocytic vesicles and mix their content for degradation, exocytosis, and recycling [8]. Finally, late endosomes or MVBs spread from early endosomes. Their fusion with the plasma membrane leads to the release of exosomes. Exosomes may be released from apoptotic bodies. The reorganization of CD9 and CD63 into tetraspanin and ESCRTs machinery that includes proteins, TSG101, Alix, and CHMP4 also acted as a golden step for exosome release. Exosome release is affected by other factors, such as Rab27a, Rab27b, and environment PH gradients. New research has shown that SphK2/S1p signaling effectively regulates the content of exosomes [8, 9, 10].
Methods and Measuring
The EVs have cargo that can be secured and unharmed from the bloodstream. On the other hand, quick and safe access to EVs allows for analyzing their content and brings a fingerprint for precision medicine. Therefore, all these characteristics made EVs a powerful diagnostic tool [11]. In the first step, EVs should be enriched purely to determine their cargo. Some methods isolated EVs commonly and some others specifically. Polymer-based precipitants, such as polyethylene glycol, reduce the solubility of EVs and lead to precipitation, followed by isolation using spin centrifugation [12]. Ultracentrifugation, the current gold standard of isolation, has limitations [13]. Tangential flow filtration and immunoaffinity-based method prevent some of the drawbacks of the previously mentioned technique [14, 15]. Currently, the isolation of EVs based on microfluidic technology is developing to achieve high-purity enrichment [16]. Secondly, EVs should be identified in terms of size, concentration, and morphology. Nanoparticle tracking is based on light scattering and the Brownian motion of EVs in solution [17]. High-resolution flow cytometry uses a lipophilic dye to intensify the signals [18, 19]. For morphological analysis, cryogenic transmission electron microscopy can be used [20]. As the best method for detection, a resistive pulse sensor changes the measurement flow across the nano pore, when an EV passes through the pores and creates the size of the information [21]. Finally, EVs cargo can be assessed by western blot, enzyme-linked immunosorbent assay, RT- qPCR, and next-generation sequencing [11].
Interaction and Uptake Mechanism
Several mechanisms have been proposed for the interaction of EVs, but it is not clear exactly which mechanism and when it acts. Exosomes and recipient cells can interact through transmembrane proteins. One of the critical proteins in this interaction is the family of lectins. Siglecs, as members of the lectin family, react with exosomes from B-cells and macrophages that have the CD169 marker. Heparan sulfate proteoglycans (HSPGs) are another example that facilitates the internalization process by interacting with the fibronectin of exosomes. Some adhesion molecules that help uptake entail CD9 and CD81 or tetraspanins on exosomes. Other ways of interaction include the fusion of vesicles with plasma membranes and the release of their contents. In addition to plasma fusion, micropinocytosis, phagocytosis, as well as clathrin-mediated, caveolin-dependent, lipid-raft- dependent, and clathrin/caveolin-independent pathways as diverse types of endocytosis may be involved. Another way of interaction is the fusion of exosomes with the endocytic vesicles of the target cell. According to a theory, target cells can control the messages transmitted from exosomes by releasing other exosomes from themselves. Polymerization of the actin from cytoskeletal proteins is involved in the process of internalization (Figure 1) [22, 23, 24].
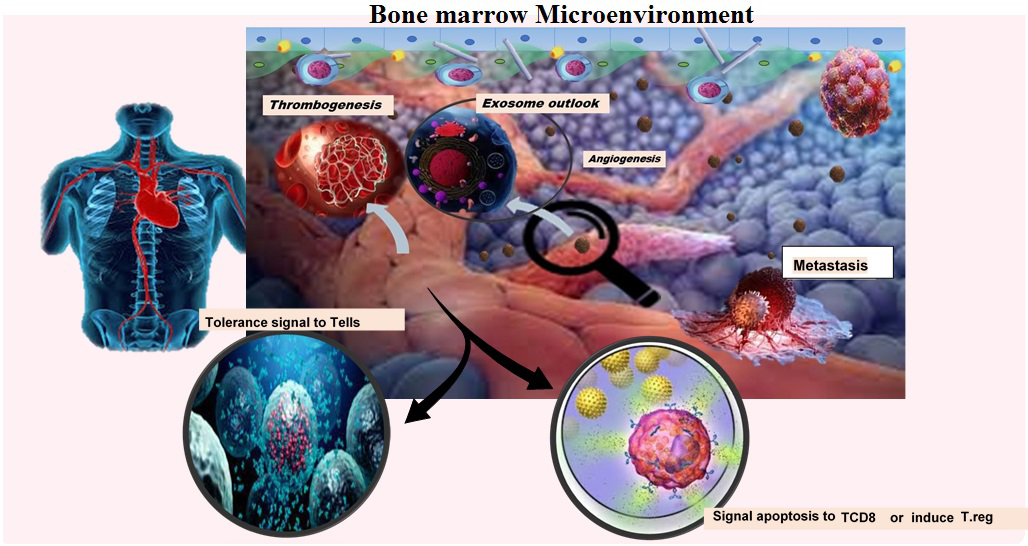
Figure 1: Extracellular vesicles and blood tumors progression. Extracellular vesicles are released from blood tumors by regulating bone marrow niche and manipulating immune cells; sending a tolerance signal to Tells, inducing regulatory T cells or apoptosis in TCD8 cells, thrombogenicity, and angiogenesis causes the development of hematological malignancies.
EVs and Drug Resistance
As a result of crosstalk between AML cells and microenvironment stroma cells, the production of IL-8 was induced by exosomes released from AML cells, causing drug resistance to etoposide. Moreover, exosomes from the bone marrow microenvironment might provide slight resistance to treatment [25]. The fibroblast growth factor (FGF) 2-FGFR1 signaling regulates the release of leukemia-protective exosomes from bone marrow stromal cells (BMSCs). On the other hand, the exosomes packed with FGF2 can induce resistance to tyrosine kinase inhibitors (TKIs) [26, 27]. Exosomes-holding apoptosis-related proteins encompass BCL-2, MCL-1, BCL-2-like, BAX, and BCL-X presenting in AML blasts relevant to relapse [28]. The VEGF/VEGFR packaging into exosomes may be pertinent to the treatment failure by inducing glycolysis in HUVECs and causing vascular remodeling (Figure 2) [29].
The PSMA3 and PSMA3-AS1-exosomes from BMSCs cause progressive resistance to proteasome inhibitors in multiple myeloma patients [30]. Bortezomib resistance occurs in multiple myeloma with del 17p, t(4;14) due to more circulating exosomes with circMYC than non- resistant patients [31]. Low levels of exosomal-microRNAs, including miR-16-5p, miR-15a-5p, miR-20a-5p, and miR- 17-5p are observed in bortezomib-resistant myeloma [32]. Delivering miR-365 with exosomes regulates the expression of apoptosis-related protein negatively in CML cells, leading to imatinib resistance [33]. In CLL, BCR signaling induced by IgM can increase exosomes with miRs content (miR-29, miR- 150, miR-155, and miR-233), which reduces the response to treatment. The effect of these miRs can be diminished by lowering the concentration of exosomes through ibrutinib therapy [34].
![Figure 2: Extracellular vesicles and malignancy drug resistance. The role of extracellular vesicles as a factor in the development of therapeutic resistance in hematological malignancies; AML, CLL, CML, and MM are briefly presented. AML: due to the association between AML malignant cells and the bone marrow microenvironment, the released exosomes are packed with IL-8 and FGF-2 and caused resistance to etoposide and TKI, respectively. CLL: the activated BCR signal releases exosomes with miRNAs contents, such as miR-29, -150, -15, and -233 that reduce therapeutic response. MM: in myeloma patients, exosomes with various cargo cause failure to bortezomib and PI. For instance, low miRNA levels, such as miR-16-5p, 15a-5p, 20a-5p, 17- 5p, and c-myc [in patients with t (4; 14) and Del 17p] as a therapeutic agent against bortezomib and the others with PSMA3 against to PI. CML: miR-365 from exosomes released from malignant cells in CML causes resistance to imatinib by negatively affecting the expression of apoptotic proteins.](/fulltextimages/9216/fig_2.png)
Figure 2: Extracellular vesicles and malignancy drug resistance. The role of extracellular vesicles as a factor in the development of therapeutic resistance in hematological malignancies; AML, CLL, CML, and MM are briefly presented. AML: due to the association between AML malignant cells and the bone marrow microenvironment, the released exosomes are packed with IL-8 and FGF-2 and caused resistance to etoposide and TKI, respectively. CLL: the activated BCR signal releases exosomes with miRNAs contents, such as miR-29, -150, -15, and -233 that reduce therapeutic response. MM: in myeloma patients, exosomes with various cargo cause failure to bortezomib and PI. For instance, low miRNA levels, such as miR-16-5p, 15a-5p, 20a-5p, 17- 5p, and c-myc [in patients with t (4; 14) and Del 17p] as a therapeutic agent against bortezomib and the others with PSMA3 against to PI. CML: miR-365 from exosomes released from malignant cells in CML causes resistance to imatinib by negatively affecting the expression of apoptotic proteins.
EVs as Therapeutic Target
The EVs can have robust therapeutic value as the result of connections between EVs and tumors. Several protocols can be used to put EVs as a therapeutic target. For instance, inhibiting the biogenesis of EVs (inhibitors for key enzymes or proteins involved in the formation of EVs, such as Alix), interfering with the secretion of EVs (dimethylamiloride as an inhibitor of Na2+/Ca2+ exchange), creating a barrier against their uptake (blocking surface adhesion molecules or another internalization receptor), and disrupting the way EVs affect recipient cells [35]. Assembling of EVs packed with miR-15a and -16 can advance the potency of Immunotherapy against Multiple myeloma. Their ability to block the connection between myeloma cells and the bone marrow microenvironment prevents Multiple myeloma survival. The miR-15a and -16 expressions can also be affected by targeting EV-LNC00461 in the circulation or tumor site. EV- miR-146a is another inhibitor of the connection between myeloma cells and stromal cells. EV-TGFB1 is a target point in AML patients by restarting the function of natural killer cells [36]. Cancerous cells in CML have a receptor for IL-13. Formulation of IL-13-expressing EVs containing BCR-ABL- suppressor RNAs can diminish the proliferation of CML cells. The use of exosomes embarrassing miR-328 can decrease the therapeutic resistance of these patients to imatinib [37]. Targeting myc-containing EVs, due to their role in the association of CLL cells with bone marrow niches, can inhibit the growth of these cells and result in resistance to treatment [36]. The HSPGs are a receptor for the internalization of EVs on target cells. Therefore, in CLL patients, pre-treating EVs with HS analog (e.g., low molecular weight heparin) can reduce their uptake into the target cell through this receptor [38]. Exosomes have natural properties, including a structure that protects their contents from nuclease and protease enzymes, a nanometer-sized phospholipid in the membrane that minimizes their detection by phagocytic immune cells, low immunogenicity, and markers that help them stabilize in the body’s biological fluids. Combining these properties has led to an avenue for applying vesicles as tools for therapeutic delivery [35].
Personalized Medicine in the Field of Hematology
Precision medicine, personalized medicine, genomic medicine, P4 medicine, and stratified medicine have almost the same meanings and can be used interchangeably. However, the term “personalized medicine” includes all the concepts and is more comprehensive. The general purpose of personalized medicine is to use appropriate doses of medicines at an accurate time based on the target genes of a patient and taking into account the patient-related factors, such as age, social status, ethical context, organ function, and the patient’s access to medical facilities. According to the pre- clinical and clinical data, the personalized medication aims to improve the anti-neoplastic effects and diminish the side effects of the medicine. Studies in the field of hematology are advancing, and personalized medicine is developing. The concept of personal medicine in the field of hematology in general is: the use of specific treatment methods that are related to different factors of each patient, in order to select the optimal diagnostic method, prognosis, classification and special treatment strategy for each individual and at the most appropriate time in blood malignancies based on the relationship of malignant cells with other cells.
Conclusion and Perspectives
The EVs are known as submicron molecules, which are of importance in multiple physiological and pathological processes due to being secretion from diverse cells. Their unique cargos, genetic and epigenetic contents, and several specious molecules can change the function of normal and cancerous recipient cells. Therefore, they play major regulatory roles in providing a microenvironment to tumor cells, disease progression, immunosuppression, and drug resistance. In this review, we discussed the biology of nano sized vesicles and their role in anti-cancer medical resistance and therapeutic purposes in some hematological malignancies. As a result, in the current era of personalized medicine, creating engineered EVs containing special personalized medications along with some elements that increase their uptake may generate different new treatment approaches. These developed EVs could be evaluated further to improve treatment and outcome. These technologies can overcome hematological malignancies recurrence and enhance the quality of life.
References
-
Auberger P, Tamburini-Bonnefoy J, Puissant A (2020) Drug resistance in hematological malignancies. Multidisciplinary Digital Publishing Institute 21(17): 6091.
-
Aharon A, Rebibo-Sabbah A, Tzoran I, Levin C (2014) Extracellular vesicles in hematological disorders. Rambam Maimonides medical journal 5(4): e0032.
-
Laurenzana I, Lamorte D, Trino S, De Luca L, Ambrosino C, et al. (2018) Extracellular vesicles: a new prospective in crosstalk between microenvironment and stem cells in hematological malignancies. Stem cells international 2018: 9863194.
-
Raposo G, Stahl PD (2019) Extracellular vesicles: a new communication paradigm? Nature Reviews Molecular Cell Biology 20(9): 509-510.
-
Gandham S, Su X, Wood J, Nocera AL, Alli SC, et al. (2020) Technologies and standardization in research on extracellular vesicles. Trends in biotechnology 38(10): 1066-1098.
-
Raposo G, Stoorvogel W (2013) Extracellular vesicles: exosomes, microvesicles, and friends. Journal of Cell Biology 200(4): 373-383.
-
Tricarico C, Clancy J, D’Souza-Schorey C (2017) Biology and biogenesis of shed microvesicles. Small GTPases 8(4): 220-232.
-
Enjeti AK (2019) Investigating the role of Microparticles/ Microvesicles/Extracellular vesicles in vascular biology, haemostasis and haemopoietic dysregulation. Blood Cells Mol Dis 8(5): 1-215.
-
Akers JC, Gonda D, Kim R, Carter BS, Chen CC (2013) Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. Journal of neuro-oncology 113(1): 1-11.
-
Shang M, Ji JS, Song C, Gao BJ, Jin JG, et al. (2017) Extracellular vesicles: a brief overview and its role in precision medicine. Extracellular Vesicles 1660: 1-14.
-
Zhao Z, Wijerathne H, Godwin AK, Soper SA (2021) Isolation and analysis methods of extracellular vesicles (EVs). Extracellular vesicles and circulating nucleic acids 2: 80-103.
-
Brown PN, Yin H (2017) Polymer-based purification of extracellular vesicles. Methods Mol Biol 1660: 91-103.
-
Momen-Heravi F (2017) Isolation of extracellular vesicles by ultracentrifugation. Methods Mol Biol 1660: 25-32.
-
Haraszti RA, Miller R, Stoppato M, Sere YY, Coles A, et al. (2018) Exosomes produced from 3D cultures of MSCs by tangential flow filtration show higher yield and improved activity. Molecular Therapy 26(12): 2838-2847.
-
Sharma P, Ludwig S, Muller L, Hong CS, Kirkwood JM, et al. (2018) Immunoaffinity-based isolation of melanoma cell-derived exosomes from plasma of patients with melanoma. Journal of extracellular vesicles 7(1): 1435138.
-
Kang YT, Purcell E, Palacios‐Rolston C, Lo TW, Ramnath N, et al. (2019) Isolation and profiling of circulating tumor‐associated exosomes using extracellular vesicular lipid–protein binding affinity based microfluidic device. Small 15(47): 1903600.
-
Gardiner C, Ferreira YJ, Dragovic RA, Redman CW, Sargent IL (2013) Extracellular vesicle sizing and enumeration by nanoparticle tracking analysis. Journal of extracellular vesicles 2.
-
Morales-Kastresana A, Jones JC (2017) Flow cytometric analysis of extracellular vesicles. Methods Mol Biol 1545: 215-225.
-
Suárez H, Gámez-Valero A, Reyes R, López-Martín S, Rodríguez MJ, et al. (2017) A bead-assisted flow cytometry method for the semi-quantitative analysis of Extracellular Vesicles. Scientific reports 7(1): 11271.
-
Cizmar P, Yuana Y (2017) Detection and characterization of extracellular vesicles by transmission and cryo- transmission electron microscopy. Methods Mol Biol 1660: 221-232.
-
Lane RE, Korbie D, Anderson W, Vaidyanathan R, Trau M (2015) Analysis of exosome purification methods using a model liposome system and tunable-resistive pulse sensing. Scientific reports 5(1): 1-7.
-
Beit‐Yannai E, Tabak S, Stamer WD (2018) Physical exosome: exosome interactions. Journal of cellular and molecular medicine 22(3): 2001-2006.
-
Gonda A, Kabagwira J, Senthil GN, Wall NR (2019) Internalization of exosomes through receptor-mediated endocytosis. Molecular Cancer Research 17(2): 337-347.
-
Schneider DJ, Speth JM, Penke LR, Wettlaufer SH, Swanson JA, et al. (2017) Mechanisms and modulation of microvesicle uptake in a model of alveolar cell communication. Journal of Biological Chemistry 292(51): 20897-20910.
-
Chen T, Zhang G, Kong L, Xu S, Wang Y, et al. (2019) Leukemia-derived exosomes induced IL-8 production in bone marrow stromal cells to protect the leukemia cells against chemotherapy. Life sciences 221: 187-195.
-
Mudgapalli N, Nallasamy P, Chava H, Chava S, Pathania AS, et al. (2019) The role of exosomes and MYC in therapy resistance of acute myeloid leukemia: challenges and opportunities. Molecular aspects of medicine 70: 21-32.
-
Javidi-Sharifi N, Martinez J, English I, Joshi SK, Scopim- Ribeiro R, et al. (2019) FGF2-FGFR1 signaling regulates release of Leukemia-Protective exosomes from bone marrow stromal cells. Elife 8: e40033.
-
Wojtuszkiewicz A, Schuurhuis GJ, Kessler FL, Piersma SR, Knol JC, et al. (2016) Exosomes secreted by apoptosis- resistant acute myeloid leukemia (AML) blasts harbor regulatory network proteins potentially involved in antagonism of apoptosis. Molecular & Cellular Proteomics 15(4): 1281-1298.
-
Yang C, Yang H, Liu J, Zhu L, Yu S, et al. (2019) Focus on exosomes: Novel pathogenic components of leukemia. American journal of cancer research 9(8): 1815-1819.
-
Xu H, Han H, Song S, Yi N, Qiu Y, et al. (2019) Exosome- transmitted PSMA3 and PSMA3-AS1 promote proteasome inhibitor resistance in multiple myeloma. Clinical Cancer Research 25(6): 1923-1935.
-
Luo Y, Gui R (2020) Circulating exosomal CircMYC is associated with recurrence and bortezomib resistance in patients with multiple myeloma. Turkish Journal of Hematology 37(4): 248-262.
-
Zhang L, Pan L, Xiang B, Zhu H, Wu Y, et al. (2016) Potential role of exosome-associated microRNA panels and in vivo environment to predict drug resistance for patients with multiple myeloma. Oncotarget 7(21): 30876-30891.
-
Min QH, Wang XZ, Zhang J, Chen QG, Li SQ, et al. (2018) Exosomes derived from imatinib-resistant chronic myeloid leukemia cells mediate a horizontal transfer of drug-resistant trait by delivering miR-365. Experimental Cell Research 362(2): 386-393.
-
Yeh YY, Ozer HG, Lehman AM, Maddocks K, Yu L, et al. (2015) Characterization of CLL exosomes reveals a distinct microRNA signature and enhanced secretion by activation of BCR signaling. Blood 125(21): 3297-3305.
-
Caivano A, La Rocca F, Laurenzana I, Trino S, De Luca L, et al. (2017) Extracellular vesicles in hematological malignancies: from biology to therapy. International Journal of Molecular Sciences 18(6): 1183.
-
Khalife J, Sanchez JF, Pichiorri F (2020) Extracellular vesicles in hematological malignancies: from biomarkers to therapeutic tools. Diagnostics 10(12): 1065.
-
Longjohn MN, Hudson J-AB, Smith NC, Rise ML, Moorehead PC, et al. (2021) Deciphering the messages carried by extracellular vesicles in hematological malignancies. Blood reviews 46: 100734.
-
Gargiulo E, Morande PE, Largeot A, Moussay E, Paggetti J (2020) Diagnostic and therapeutic potential of extracellular vesicles in b-cell malignancies. Frontiers in Oncology 10: 580874.
- How to Identify and Overcome Barriers in Developing Blood Systems?
- Why Was Transfusion Medicine Not Recognized as a Clinical Discipline?
- Outcomes of Lenalidomide Relapsed/Refractory Patients
- Is Transfusion Always Necessary?
- The Logistics of Production and Use of Blood and Blood Components
- The Challenge for Component Therapies