Geographical Assignment and Molecular Tracking in Wildlife Offences: A Case Study of Seized Elephant Tail Hair
Tracking geographic origin of parts traded in wildlife offences is a critical aspect of wildlife management which enables to strengthen ground level enforcement and in determining changes taking place in population demography for planning long-term conservation goals. Asia and Africa are habitats to very rich faunal diversity and many species in these continents are victims of wildlife trafficking. Of the different species, elephants are more prone to illicit trafficking in India as well as in Africa because of the high demand for products like ivory, meat, tail hairs, etc. Use of nuclear markers has been well documented in tracking African elephant parts like ivory but precluded its use in parts like tail hair that lack root part, hence the source of nuclear DNA is lost. In such cases, population-based mtDNA haplogroups has been an alternate approach in tracking the source of origin. This study presents an effort to implement DNA-based methods for the forensic identification of species as well as a source of origin of elephant tail hair confiscated from different locations in India and Nepal. A total of 15 hair samples seized in India and Nepal were undertaken for study and were subjected to species identification using Cyt b gene (ca.350 bp) and compared with published haplogroups in different populations for the source of origin. Out of these, eight samples showed success rate with good PCR amplification and seven samples were eventually dropped because PCR failed. This may have been due to either PCR inhibitor, or DNA template is of low molecular weight. Based on obtained sequences of Cyt b and after alignment with the reference sequences from Asian and African elephant taken from the previous study, it was observed that all the investigated samples belong to the African elephant. For the geographical origin of these samples, we used reported available haplotype homology of African elephant. It was inferred that seven samples seized from India showed geographical origin from different locations of the north, central and east African regions while the sample seized in Nepal indicate its origin from Central-West African region. Thus, the study emphasis on the use of mtDNA marker to identify species from confiscated samples and source of origin. We suggest a need of establishing genetic database of species across its range which are prone to poaching at the global level to track back the actual hot-spots of illegal hunting to effectively enforce the laws and improve conservation efforts of protecting such key wildlife species
Introduction
Wildlife crime is one of the biggest threats to conservation efforts globally. Trafficking of wildlife parts and products are mainly driven by demand in international and local markets for consumption of meat, use in Traditional Chinese Medicine (TCM), trophies, jewelry and ornamental items [1]. Due to the high demand of wildlife products (skin, bones, antlers, whiskers, claws, ivories, etc.) in the international market, extensive exploitation of wildlife has been reported especially in Asia and Africa [2, 3]. Literature clearly reveals consequences of poaching for illegal trade on population demography because most of these are sex- biased, and males are known to be poached more than females [4]. Due to skewed poaching, such populations have high inbreeding coefficients because random mating was absent. Hence, identifying such populations prone for illegal trafficking is crucial so as to strengthen ground enforcement and plan adaptive strategies if needed to restore random mating for achieving long-term conservation goals. Presently, with the advancement in molecular techniques, ecology and use of spatial variation in genetic profiles using nuclear and mtDNA markers, it is possible to effectively determine poaching hot spots by tracking back the geographic origin of traded parts at different scale and the same has been demonstrated in turtles, sharks and elephant [5, 6, 7]. Of the different species reported in wildlife trade, elephants (African and Asian species) are more prone to poaching for ivory, meat and other parts like tail hairs [7, 8]. Elephants are classified as vulnerable by the International Union for Conservation of Nature (IUCN) and included in CITES (Appendix 1) as species of concern due to increasing illicit activities [8]. To effectively minimize such illegal poaching across the world it is important to identify ‘hot spots’ of poaching and to enforce stringent Wildlife Protection Laws in such areas for better conservation. As per the Wildlife Protection (Act) of India (1972), possession of any wildlife parts or product is illegal, but the law is restricted to species inhabiting in the Indian territory. Internationally traded species are governed by CITES treaties and regulations. The utility of DNA-based species identification in wildlife forensics has been a powerful tool especially in bush meat and pet shop [9, 10]. Wasser et al. (2004) [11] demonstrated the use of nuclear markers for tracking elephant ivory back to the geographic region of origin. However, some of the elephant parts reported in wildlife trade like tail hair usually lack root which is a source of nuclear DNA and precluded the use of these markers. In such cases, population-based haplogroups observed in mtDNA genome may be an alternate approach for tracking back to population and has been demonstrated its use in the meat traded and in molecular tracking. The present study reports an investigative approach for establishing the species identification and geographic origin of elephant tail hairs confiscated by government authorities in India and Nepal using DNA forensics and the assessment of the utility of the extensive database of mtDNA genome available at NCBI (USA).
Material and Methods
Case history
In March 2011, during a routine check-up, the Customs Department of India apprehended two parcels sent from Africa via Air-mail. Once the packets were opened for preliminary screening, they were found to be containing 15 kg of thick hairs or synthetic thread-like material. Based on the preliminary examination, the seized material was suspected to be elephant tail hairs and the authorities forwarded the samples for forensic analysis to the Wildlife Institute of India (WII). In an another case, the Department of National Park and Wildlife Conservation, Kathmandu, Nepal also seized a parcel, and that was also forwarded WII us for forensic examination. The morphology of the material suggested the presence of tail hairs of an elephant, and it was not possible to differentiate based on hair characteristics between Asian and African elephants due to lack of adequate reference data. All these hair were without any root. Hence, a combined approach of molecular biology and spatial genetic characteristics of mtDNA haplogroups was then used to investigate the species and geographical tracking of the seized samples.
DNA extraction, PCR amplification and sequencing
A total of 15 samples were analyzed in the present study (Table 1).
| Sample ID | Area | Total number of samples used in study(n=) | Amplification of Cyt b locus in used samples |
|---|---|---|---|
| ETH 01-ETH 12 | India | 12 | 07 (ETH 01-ETH 07) |
| ETH 13-ETH 15 | Nepal | 3 | 01 (ETH 13) |
Table 1: Detail of elephant seizure confiscated in India and Nepal and result of Cyt b PCR amplification.
All samples were cleaned using alcohol and cut into 20 mm kept in different tubes and Genomic DNA was extracted using Qiagen DNeasy Tissue Kit (QIAGEN, Germany) as per the manufacturer’s protocol. A fragment of the cytochrome b gene (~350 bp) was amplified using the universal PCR primers developed by Meyer et al. (1995) [12] for species identification. All PCR reactions were carried out on an ABI 9600 Fast thermo cycler (Applied Biosystems, USA) in a total reaction volume of 25 µl containing 1x PCR buffer, 25 mM MgCl2, 10 mM dNTPs, 10 µM of each primer, 2.5 U Taq polymerase (ABI, Fermentas) and 3 µl of total DNA template. The PCR cycling conditions were as follows: initial denaturation at 94°C for 3 min, followed by 35 cycles of denaturation at 94°C for 30 s, primer annealing at 53°C for 45 s, primer extension at 72°C for 35 s, and a final extension at 72°C for 10 min. PCR products were subjected to electrophoresis on 2% agarose gel to visualize the presence of amplification using gel electrophoresis. The PCR amplification with a forward primer (Cyt b 381, H, 5’- CCATCCAACATCTCAGCATGATGAAA-3’) was used for the cycle sequencing reaction using the Big Dye Terminator Cycle Sequencing kit® v 3.1. Sequencing reaction products were purified using a standard ethanol precipitation method and sequenced on an ABI 3130 Genetic Analyzer (Applied Biosystems, USA) following the manufacturer’s recommendations. Positive and negative controls were performed throughout all DNA extraction and PCR amplifications. Duplicates were analyzed for each sample.
Data analysis
Sequences were validated using SEQUENCHER 4.8 (Gene Codes Corporation, Ann Arbor, MI, USA). Cyt b sequences of Asian elephants (Elephas maximus) used in this study were previously sequenced in our Forensic Laboratory and sequences of African elephants were taken from previous studies (Table 2).
| Taxon | Elephant Cyt b haplotype | Geographic origin | GenBank accession |
|---|---|---|---|
| L.a. africana | Hap-01 | Tanzania, Mozambique | AY741324,AY741323 |
| L.a. africana | Hap-02 | Uganda, Namibia, Botswana, Democratic Republic of Congo (DRC), Zimbabwe | AY741078,AY741076, |
| L.a. africana | Hap-02 | Uganda, Namibia, Botswana, Democratic Republic of Congo (DRC), Zimbabwe | AY741325,AY742801, |
| L.a. africana | Hap-02 | Uganda, Namibia, Botswana, Democratic Republic of Congo (DRC), Zimbabwe | AY742800 |
| L.a. cyclotis | Hap-03 | Angolia, Sudan | AY741072,AY741073 |
| L.a. cyclotis | Hap-04 | Chad, Camerron-01, Democratic Republic of Congo (DRC) Camerron-02, Uganda | AY741080,AY359269, |
| L.a. cyclotis | Hap-04 | Chad, Camerron-01, Democratic Republic of Congo (DRC) Camerron-02, Uganda | AY359270,AY359267, |
| L.a. cyclotis | Hap-04 | Chad, Camerron-01, Democratic Republic of Congo (DRC) Camerron-02, Uganda | AY359279 |
| L.a. cyclotis | Hap-05 | Democratic Republic of Congo (DRC),Central African Republic (CAR), Republic of Congo (RC) | AY359268,AY359272, |
| L.a. cyclotis | Hap-05 | Democratic Republic of Congo (DRC),Central African Republic (CAR), Republic of Congo (RC) | AY359273 |
| L.a. cyclotis | Hap-06 | Gabon | AY359274 |
| L.a. cyclotis | Hap-07 | South Africa | AY741320 |
| L.a. africana | Hap-08 | Zimbabwe | AY741321,AY741322 |
| L.a. cyclotis | Hap-09 | Liberia | AY741079 |
| L.a. africana | Hap-10 | Ivory Coast | AY741327 |
| L.a. africana | Hap-11 | Republic of Congo (RC)-Gabon, Gabon | AY359266,AY359278 |
| E. maximus | Hap-12,Hap-13 | India | EMU23740,AY791138.1 |
| L.a. africana | Hap-14 | Kenya | JQ438446,JQ438441 |
| JQ438134,JQ438131 | ||
| Ghana | JQ438737,JQ438738 | |
| JQ438754,JQ438740 |
Table 3: Details of published region specific different cytochrome b based geographic haplotype of forest (Loxodonta Table 2: Det
Table 2: Details of published region specific different cytochrome b based geographic haplotype of forest (Loxodonta Table 2: Details of published region specific different cytochrome b based geographic haplotype of forest (Loxodonta africana cyclotis), Savana (Loxodonta africana africana) African elephant and its Accessions number. We then aligned to reference sequences retrieved from NCBI/GenBank (http://www.ncbi.nlm.nih.gov/database) using CLUSTAL W implemented in BioEdit version 7.0.5.3 [13]. The sequence alignment was used to generate neighbor-joining trees using the MEGA 5 software [14].
Results and Discussion
Of the total samples, we selected randomly 15 tail hair from India (n=12) and Nepal (n=3) in this study. We observed visible DNA on gel only for eight samples
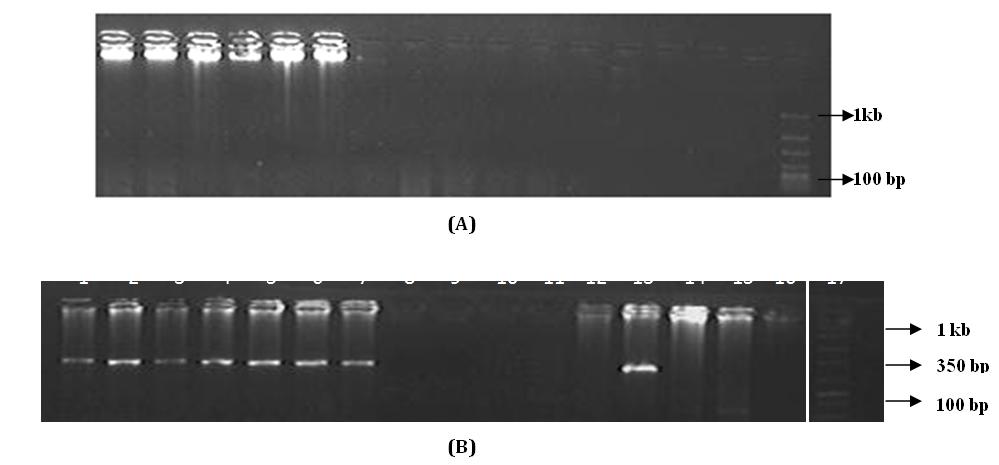
inhibitor. Therefore, all these samples were again put PCR with a lower fragment of the mitochondrial gene (ca.159 bp) [15] but again we did not get PCR amplification. Therefore, all non-amplified samples were left from the study and may have PCR inhibitor. After the sequences retrieved from case samples were BLAST on NCBI, seized samples showed 100 % similarity with the African elephant and indicated that examined all tail hair samples were of African elephants (Table 3).
| Sample | Species with highest similarity | Common name | Query coverage (%) | Similarity (%) |
| Sample | Common name | |||
| ETH 01-ETH 07, ETH13 | Loxodonta africana | African elephant | 100 | 100 |
| ETH13 | Loxodonta africana | African elephant | 100 | 100 |
| ETH 01-ETH 07 | E. maximus | Asian elephant | 100 | 98 |
| ETH13 | E. maximus | Asian elephant | 100 | 98 |
Table 4: NCBI sequence similarity in the Cyt b locus between elephant seizure with African elephant (Loxodonta Table 3: NCBI sequ
Table 3: NCBI sequence similarity in the Cyt b locus between elephant seizure with African elephant (Loxodonta Table 3: NCBI sequence similarity in the Cyt b locus between elephant seizure with African elephant (Loxodonta africana) and Asian elephant (Elephas maximus). The phylogenetic tree generated based on Kimura 2 parameter also suggest that seized samples match with African elephant with high boot strap value (Figure 2).
![Figure 2: Haplotypes based Neighbor-joining tree representing the phylogenetic relationships and geographic distribution of Cyt b haplotypes of publicly available elephant sequences and sequences derived from elephant tail hairs apprehended in India (ETH-01 to ETH-07) and Nepal (ETH-13). Details of Haplotypes origin and accessions number given in (Table 2). Numbers near internal (%) derived from 1,000 bootstrap values. The taxonomic status of the African elephant (_Loxodonta africana_) has been debated since the turn of the 20th century [16], and up to 22 subspecies have been described [17,18]. However, modern taxonomy and recent molecular studies refers to two types, with namely the savannah (_Loxodonta africana africana_) and the forest (_Loxodonta africana cyclotis_) African elephants. Their names were reflecting the habitat in which they are found. With the advances in molecular genetics, the emphasis has been in determining the source of origin of traded parts, so as likely impacts on population demography may be understood and appropriate measures may be taken for avoiding any inbreeding through establishing connectivity among the population in meta-population framework. Population assignment in molecular genetics has been a powerful tool in determining source of origin of traded parts based on spatial genetic information within the range of species by using nuclear or mtDNA markers. Wasser et al. (2004) demonstrated how elephant ivory may be tracked back to geographic region using nuclear markers (i.e. multi locus genotyping). However, use of this approach has precluded in the items made of hair lacking root part as this is source of nuclear DNA in such articles. Alternatively, mtDNA haplogroups have also been used in population assignment for the parts reported in the trade. We compare the haplotypes of seized samples for Cyt b sequences with reported haplogroups available for same gene data of different populations of African elephants of known geographic origin [19] (Table 2). It was possible to concluded that 7 samples of elephant tail hair examined from the seizure in India, (ETH-02,ETH-03), (ETH-05, ETH-06) and (ETH-01,ETH-04) matched with (North- Central and Central-East forest/savannah), (Central-West forest) and (Central Africa forest) African elephant haplotypes with (57%), (64%) and (43%) bootstrap value and displayed 99.09% to 100% similarity. Whereas, remaining one sample (ETH-07) matched with (South- Eastern and Central Savannah) African elephants haplotypes with 53% boot strap values and displayed 100% similarity. In a case of sample seized from Nepal(ETH-13), it matched with (North Central and Central-East forest/savannah) African elephant haplotypes with 57% boot strap value and displayed 100% similarity (Figures 2 & 3,Table 4).](/fulltextimages/507/fig_2.png)
Figure 2: Haplotypes based Neighbor-joining tree representing the phylogenetic relationships and geographic distribution of Cyt b haplotypes of publicly available elephant sequences and sequences derived from elephant tail hairs apprehended in India (ETH-01 to ETH-07) and Nepal (ETH-13). Details of Haplotypes origin and accessions number given in (Table 2). Numbers near internal (%) derived from 1,000 bootstrap values. The taxonomic status of the African elephant (Loxodonta africana) has been debated since the turn of the 20th century [16], and up to 22 subspecies have been described [17, 18]. However, modern taxonomy and recent molecular studies refers to two types, with namely the savannah (Loxodonta africana africana) and the forest (Loxodonta africana cyclotis) African elephants. Their names were reflecting the habitat in which they are found. With the advances in molecular genetics, the emphasis has been in determining the source of origin of traded parts, so as likely impacts on population demography may be understood and appropriate measures may be taken for avoiding any inbreeding through establishing connectivity among the population in meta-population framework. Population assignment in molecular genetics has been a powerful tool in determining source of origin of traded parts based on spatial genetic information within the range of species by using nuclear or mtDNA markers. Wasser et al. (2004) demonstrated how elephant ivory may be tracked back to geographic region using nuclear markers (i.e. multi locus genotyping). However, use of this approach has precluded in the items made of hair lacking root part as this is source of nuclear DNA in such articles. Alternatively, mtDNA haplogroups have also been used in population assignment for the parts reported in the trade. We compare the haplotypes of seized samples for Cyt b sequences with reported haplogroups available for same gene data of different populations of African elephants of known geographic origin [19] (Table 2). It was possible to concluded that 7 samples of elephant tail hair examined from the seizure in India, (ETH-02,ETH-03), (ETH-05, ETH-06) and (ETH-01,ETH-04) matched with (North- Central and Central-East forest/savannah), (Central-West forest) and (Central Africa forest) African elephant haplotypes with (57%), (64%) and (43%) bootstrap value and displayed 99.09% to 100% similarity. Whereas, remaining one sample (ETH-07) matched with (South- Eastern and Central Savannah) African elephants haplotypes with 53% boot strap values and displayed 100% similarity. In a case of sample seized from Nepal(ETH-13), it matched with (North Central and Central-East forest/savannah) African elephant haplotypes with 57% boot strap value and displayed 100% similarity (Figures 2 & 3,Table 4).
![Figure 3: Geographical assignment of elephant tail hair seizure. Out of 8 samples undertaken for analysis, 3 samples originated from North-Central Africa {(02 [ETH-02,03] seized in India and 01 [ETH-08] seized in Nepal) }, 02 samples originated from Central-west forest Africa {(All 2 [ETH-05&06] seized in India) },02 sample originated from Central forest Africa {(All 2(ETH-01 &04) seized in India)} and Last 01 originated from Central Savannah {01 sample(ETH-07) Seized in India)}.](/fulltextimages/507/fig_3.jpeg)
Figure 3: Geographical assignment of elephant tail hair seizure. Out of 8 samples undertaken for analysis, 3 samples originated from North-Central Africa {(02 [ETH-02,03] seized in India and 01 [ETH-08] seized in Nepal) }, 02 samples originated from Central-west forest Africa {(All 2 [ETH-05&06] seized in India) },02 sample originated from Central forest Africa {(All 2(ETH-01 &04) seized in India)} and Last 01 originated from Central Savannah {01 sample(ETH-07) Seized in India)}.
Taxon with the highest
Query coverage Similarity Haplotype Geographic origin similarity (%) (%)
Sample
Democratic Republic of Congo (DRC), Central African Republic (CAR),Republic of Congo (RC) ETH-04 L. a cyclotis 100 100 ETH-02 L. a cyclotis 100 100 ETH-01 L. a cyclotis 100 99.09 Hap-05 DR Congo,Uganda ETH-03 L. a cyclotis 100 100 Cameroon, ETH-13 L. a cyclotis 100 100 Chad Hap-04
| L. a cyclotis | 100 | 100 | Hap-11 | Republic of Congo (RC)-Gabon, Gabon | |
| ETH-05 | L. a cyclotis | 100 | Hap-11 | Republic of Congo (RC)-Gabon, Gabon | 100 |
| ETH-06 | L. a cyclotis | 100 | 100 | Hap-02 | Zimbabwe, Uganda |
| ETH-07 | L. a africana | 100 | 100 | Hap-02 | Namibia, Botswana |
Table 5: Assigning the geographical origin of elephant seizure based on sequences similarity with published Cyt b locus Based on
Table 4: Assigning the geographical origin of elephant seizure based on sequences similarity with published Cyt b locus Based on the haplotypes similarity, it was thus possible to track back the approximate geographic location of seized tail hairs from elephants inhabiting in African regions using molecular and DNA forensic techniques. Conclusion Literature indicate that, the menace of illegal wildlife trafficking can be controlled by identifying the hot-spots of poaching using DNA-based molecular techniques of using mtDNA [9, 10] and nuclear markers [20]. Once the information on these aspects is known, it would help in rendering the better enforcement services for the protection of wildlife globally. The results from the present study demonstrate the use of molecular biology techniques along with spatial genetics and geographical data in identifying species and obtaining insight into the molecular tracking to the geographic region of origin using published information on mtDNA haplogroups for the samples where obtaining nuclear DNA is difficult. Findings of the present study and other studies using, haplotypes of mtDNA genome, may be useful in understanding likely impacts of population demography of the species due to illegal poaching. Our study shows the importance of genetic analysis in wildlife forensics and the utility of DNA-based analysis in the implementation of CITES (Convention on International Trade in Endangered Species). Hence, we suggest it is necessary to establish and promote the use of such DNA profile database of mitochondrial DNA across species’ range which is of global conservation importance and reported in wildlife trade. Such information can easily be applied to use across different labs, as comparing results of multi locus data across different labs are of limited use at present in wildlife due to lack of allelic ladder for species.
Acknowledgments
The authors are grateful to the Director, Dean and Research Coordinator, Wildlife Institute of India, Dehradun, for their strong support and facilitation. The authors acknowledge the support provided by the Nodal Officer and all the researchers and staff of the Wildlife Forensic and Conservation Genetic Cell of the Wildlife Institute of India for carrying out this study.
Competing interest
The authors declare that they have no competing interest.
Ethical Statement
Ethical approval was not required for this study because all samples were collected from wildlife forensic and conservation genetic cell.
Funding
The study funded by WII grant in aid, Ministry of Environment, Forest and Climate Change, New Delhi, India.
References
-
Arumugam T (2015) Endangered wildlife species on the plate. New straits time.
-
WWF (2014) Illegal Wildlife Trade in India Special Issue.
-
(2014) Traffic Bulletin is vested in Traffic International © Traffic International.
-
Mondol S, Celia R Mailand, Samuel K Wasser (2014) Male biased sex ratio of poached elephants is negatively related to poaching intensity over time. Conservation Genetics 15(5): 1259-1263.
-
Amorocho DF, Abreu-Grobois FA, Dutton PH, Reina RD (2012) Multiple Distant Origins for Green Sea Turtles Aggregating off Gorgona Island in the Colombian Eastern Pacific. Plos One.
-
Barbuto M, Galimberti A, Ferri E, Labra M, Malandra R, et al. (2010) DNA barcoding reveals fraudulent substitutions in shark seafood products: The Italian case of ‘‘palombo” (Mustelus spp.) Food Research International 43(1): 376-381.
-
Martin E, Stiles D (2000) The Ivory Markets of Africa Save the Elephants, Kenya.
-
Blanc J (2008) Loxodonta africana. In: IUCN 2012. IUCN Red List of Threatened Species Version 2012.
-
Lee SJ, Jung G, Min MS, Kim CK, Lee H, et al. (2011) Genetic origin identification of Siberian chipmunks (_Tamias sibericus_) in pet shop of South Korea, Animal Cells and System 15(2): 161-168.
-
Koh HS, Wang J, Lee BK, Yang BG, Heo SW, et al. (2009) A phylogroup of the Siberian chipmunk from Korea (_Tamias sibericus barberi_) revealed from the mitochondrial DNA cytochrome b gene. Biochem Genet 47(1-2): 1-7.
-
Wasser SK, Shedlock AM, Comstock KE, Ostrander EA, Mutayoba B (2004) Assigning African elephant DNA to geographic region of origin: applications to the ivory trade. _PNAS_ 101(41): 14847-14852.
-
Meyer R, Hofelein C, Luthy J, Candrian U (1995) Polymerase chain reaction-restriction fragment length polymorphism analysis: a simple method for species identification in food. J AOAC Int 78: 1542- 1551.
-
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41: 95-98.
-
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, et al. (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28(10): 2731-2739.
-
Berger JW, Rubin PA, Jakobiec FA (1996) Pediatric orbital pseudotumor: case report and review of the literature. Int Ophthalmol Clin Winter 36(1): 161- 177.
-
Barriel V, Thuet E, Tassy P (1999) Molecular phylogeny of Elephantidae. Extreme divergence of the extant forest African elephant. C R Acad Sci III 322(6): 447-454.
-
Roca AL, Georgiadis N, O'Brien SJ (2007) Cyto-nuclear genomic dissociation and the African elephant species question. Quat Int 169-170: 4-16.
-
Rohland N, Reich D, Mallick S, Meyer M, Green RE, et al. (2010) Genomic DNA Sequences from Mastodon and Woolly Mammoth Reveal Deep Speciation of Forest and Savanna Elephants. PLOS BIOLOGY 8(12).
-
Johnson MB, Clifford SL, Goossens B, Nyakaana S, Curran C, et al. (2007) Complex phylogeographic history of central African forest elephants and its implications for taxonomy. BMC Evolutionary Biology 7: 244.
-
Wasser SK, Brown L, Mailand C, Modol S, Clark W, et al. (2015) Weir BS: Genetic assignment of large seizures of elephant ivory reveals Africa’s major poaching hotspots. Science 349(6243): 84-87.
- Forensic Implications of Adverse Drug Reactions in Schizophrenia A Case Series
- Narcotics and Digital Forensics: Bridging Crimes in the Digital Age
- Ethics in Forensic Psychiatry: Principles, Dilemmas, and Human Rights
- Impact of Acute Stress on Attentional Orienting to Social Cues
- Head Injury and Intracranial Hemorrhage in Western Region of Libya
- A Forensic Study on Handedness: Examination of Handwriting Features in Right and Left Handed Writers