Do Red Foxes (Vulpes Vulpes) Increase the Detectability of Scent Marks by Selecting Highly Conspicuous Substrates
In mammals, especially those that are nocturnal or crepuscular, chemical marks usually play a significant role as environmental labels. Scent marks in mammals can have many functions, including territorial defense and communication signals in the mating season. Furthermore, animals can increase the detectability of marks by selecting highly conspicuous locations and substrates on which to mark, such as stones and sticks, on or around faeces, other animal carcasses, anthropogenic features, and plants. Human-carnivore interactions can result in a conflict where perceived damage to livelihoods occurs in socio-economically poor areas. In the Shigar Valley, Karakorum Range, Pakistan, subsistence mixed farming is the predominant land use and red foxes (Vulpes vulpes) are widely persecuted. Understanding the effects of human activity and habitat factors on fox behaviors in the region are lacking. We used line transect surveys aided by a domestic dog (Canis familiaris) to detect fox faeces locations and characterize fox den form 2015-16-17. We encountered 467 fox scats by dog assistance. We tested for associations between scent deposition, and environmental variables including vegetation and substrate type, distribution of livestock carcasses and dung, roads and agricultural land at micro-habitat. We found more fox scats on aromatic plants, shrubs, and on or near livestock feaces and carcasses. Fox scent marking site selection varied for forested and shrub areas relative to open grassland or barren ground and vegetation cover and tree density at the microhabitat scale. The findings suggest that foxes do not avoid human activity per se and anthropogenic benefits (road kill, livestock carrion, crop food sources) may outweigh the costs of persecution. Foxes may also provide ecosystem benefits via seed dispersal, carrion removal and regulation of prey populations.
Introduction
In mammals, especially those that are nocturnal or crepuscular, chemical marks usually play a significant role as environmental labels [1]. Scent marks in mammals can play many functions [2], including territorial defense and communication signals in the mating season. Furthermore, animals can increase the detectability of marks by selecting highly conspicuous locations and substrates on which to mark, such as stones and sticks, on or around feces, other animal carcasses, anthropogenic features, and plants that may strengthen signaling potential [3]. However, associations between fecal marking site selection and vegetation type are infrequently explored or reported [4].
Barja, et al. [5], determined that red foxes (Vulpes vulpes) in Galizia, northern Spain, marked with the feacal matter around woody plants to a greater extent than herbaceous plants. Plant selection for scent marks appears to be important in the case of rubbing behavior by black bear (Ursus americanus, Pallas, Burst and Pelton [6], as well as for urine marking by Barbary lions (Panthera leo leo L.) and Siberian tigers (Panthera tigris altaica, Temminck) in Madrid Zoo [7]. Both white-tailed deer (Odocoileus virginianus) and Alaskan moose (Alces alces, L.) chose aromatic species of trees for rubbing Bowyer, et al. [8], and Barja, et al. [5] found that Iberian wolves (Canis lupus signatus, Cabrera) urinate mostly on trees with a thicker trunk.
Chemical marking structures play an influential role in communicating with sympatric mammals [1, 9]. Moreover, foxes and lagomorphs are known to use clearings and landmarks for scent-marking behaviors [10, 11]. Previous studies indicate that foxes defend their food resources from conspecific and heterospecific competitors using markings [10].
The Canidae comprise a principal group of predators [12], which typically shelter in dens dug into the ground, rock cavities, crevices, caves or hollows [13, 14]. The red fox utilizes dens for birthing and rearing offspring (‘breeding dens’) and as resting sites outside the breeding periods (‘non-breeding dens’) [15]. Lactating females spend prolonged periods within the breeding den with their cubs during the initial few weeks of life [16].
The availability and quality of resources influence the size of the animal home range [17], such that, in canids, for example, a home range in a resource-rich area may be smaller than in a resource-poor area (e.g., 0.4 km2 and >40 km2 respectively [14]).
In northern Pakistan, Central Karakoram National Park (CKNP) is inhabited by major wildlife species of global importance such as snow leopard(Panthera uncia), some of which prey on or competitively displace red foxes [18, 19]. Locally, the red fox is also a primary predator of small mammals (e.g, Royle’s Pika or Indian Pika (Ochotona roylei) [20]. Major factors affecting red fox behavior and populations include roads and varying agricultural practice, whilst retaliatory killing for crop damage and prey depletion, local perception of the red fox as a pest species [21, 22]. Although animals may use any of their diverse sensorial channels to obtain this information, most mammals are nocturnal and live in grassland. As a result, olfactory communication plays a principal role [3]. However, resource selection by red foxes in the region has not previously been subject to study and the effect of human disturbance on fox behavior and densities are unknown [23]. Here we hypothesize that, Do domestic dog can assistance to improve the detection of scats, red foxes choose specific substrates and aromatic or medicinal plants as signal posts in different habitats and also helping to the dispersal of seeds. Does fox use olfactory communication to interactions with opposite sexes?
Material and Methods
Study Location
This study was conducted in the Shigar Valley in the Karakorum range located along the north bank of the river Indus. It lies at 25° 25′32″ N latitude and 75° 42′59″E longitude and covers an area of 4373 sq. km with altitudinal amplitudes of 2, 260 to 8611 m above sea level including K2 (8611 m), Broad Peak (8047 m), Angel Peak (6858 m) and Skil Brum (7360 m) [24] see (Figure 1). We conducted this observational and non-invasive study according to the regulations for animal welfare and conservation under the Gilgit- Baltistan Wildlife Preservation Act 1975 and the North-east Forestry University Guidelines for the Use of Animals in Research.
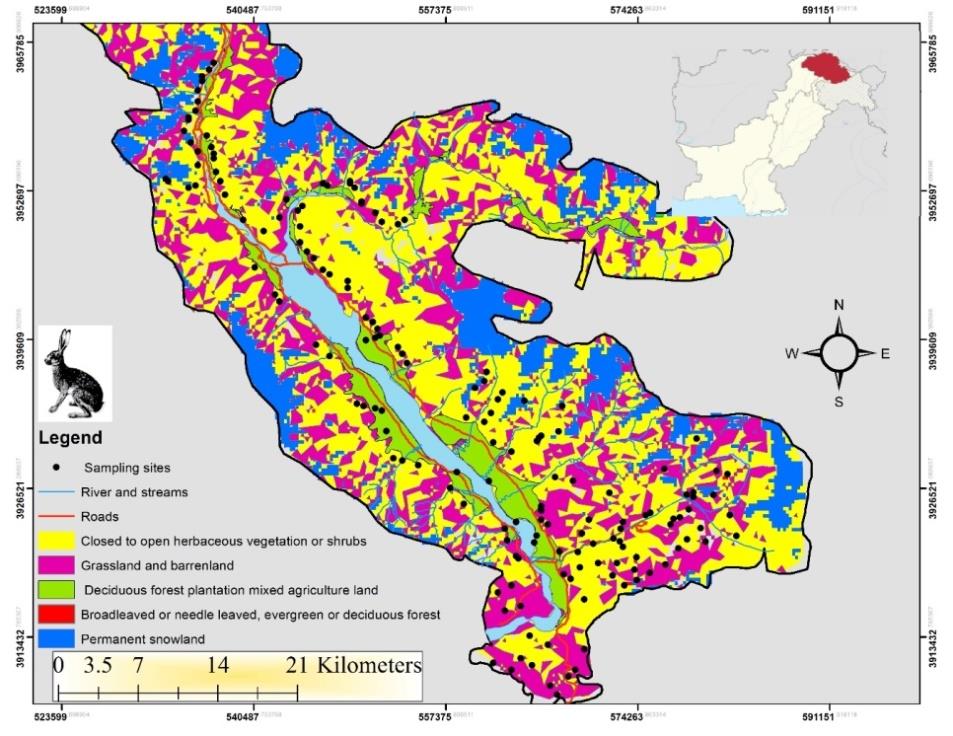
Collection of Territory Marking Data
We used line transect and domestic dog (Canis familiaris) to detect feces throughout the 24h diel period, at night aided by torchlight [25, 26]. We established a temporary 200 sample plot (10 m ×10 m) . We searched for fox faeces in each plot, and for every faecal sample detected we recorded associations with plant species following the technique described by de Miguel, et al. [27]. We considered an association to have occurred when feces were deposited adjacent to a plant (≤ 0.5 m) or on top of it. We also noted the classification and species of plant: wooden/herbaceous plants or shrubs, including aromatic herbaceous plants and shrubs within each plot (measured at 1 m above ground level). We additionally recorded carcasses, trails, livestock dung and rocks within ~2 - 5 m of the faeces where they occurred within the plot, and recorded the association as deposited on or near these substrates. The following plant species were considered: tree (woody) species (Prunus armeniaca , Morus nigra, Salix alba , Elaeagnus angustifolia), and herbaceous species (Datura fastuosa, Mentha royleana Benth, Rheum webbianum Royle) and shrubs (Artemisia absinthium, Astragalus psilocentros Fisch, Capparis spinosa Linn, Hippophe rhamnoides, Ephedra gerardiana Wall, Rosa webbiana). The seeds of food plants such as Hippophe rhamnoids, Prunus armeniaca, Morus nigra, Elaeagnus angustifolia and Rosa webbiana were found in fox feaces and were, therefore, also considered.
Data Analysis
Regarding feces as territory marks, we calculated the frequency of feaces associated with different substrates in different habitats, taking into consideration the usual distribution of fox feces within the plots. Our data did not follow a normal distribution and independence of the data was not assured as we did not perform genetic analyses to identify individual, and therefore the identity of the feces was unknown. Thus we used a Wilcoxon test [28]. And For each plot, we classified vegetation cover according to protocols described by Wang et al. [29], as well as the relative density, frequency, cover, and importance value index (IVI) of plant species, as described by Mahmood, et al. [30]. We also did not include 170, feaces in the final analysis due to bias. The Bonferroni Z-statistics were performed in SPSS 20.00 (SPSS Inc., and Chicago, Illinois, USA).
Results
Territory Marks
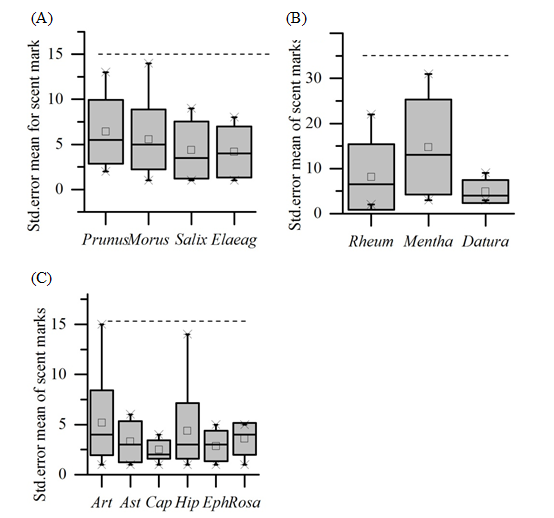
We identified in total of 467 fox scats by dog assistance and direct observation. In total, 44.4 % were associated with aromatic shrubs including Artemisia absinthium and Capparis spinosa in both barren areas and forest (Figure 2c). In the barren and open grassland, 18.4 % of the feces were associated with aromatic herbs, mostly Mentha royleana Benth and to a lesser extent Datura fastuosa (Figure 2b). Foxes favored shrub species over herbaceous species (Z = -2.497; P = 0.013). Of the woody plants, 35.1 % of fox feces were related to fruit trees in the forest, with the greatest frequency of scat linked to Prunus armeniaca, and lowest associated with Elaeagnus angustifolia (Figure 2a). Scat deposition was not significantly different between shrubs and trees (Z = - 0.537; P = 0.591). Furthermore, fox feaces were significantly associated with livestock feaces and animal carcasses in the open grassland than barren areas (Z = - 2.106; P = 0.02). The trend was observed where fox feaces were associated with rocks and animal trails in both the barren and open grassland habitats, but this was not statistically significant (Z = 0.161; P = 0.872).
Discussion
We found that foxes selected specific substrates or fruits trees and aromatic medicinal plants for territorial signal marking sites. Ecological factors such as temperature, substrate, and air currents, as well as experience levels, are known to influences the abilities of scat detection dogs [31]. The transmission of information by means of chemical signals is fundamental for many animals, playing a key function in the biological success of the individuals [32]. Foxes select plants as signal posts, for this reason, marking in places where they can be easily received by conspecifics.
As we predicted, the association of feaces with plants was notably higher in barren land than a forest, possibly because it was more detectable in the former. Polunin described foxes depositing scats on oaks and rockrose plants; hence they found these plants to have an attractive fragrance and sticky gums. Punjabi, et al. [33] found that wild fruit (viz. Zizyphus mauritiana and Cássia sp.) are significant components of Indian fox diet. Therefore, we expected wild fruit to positively influence the selection of signal posts at a smaller scale. In our results, woody plants were marked less than shrubs and herbaceous species, which contrasts with the findings of Barja, et al. [5] in Galizia, Spain. Carnivore scent profiles consist of sex-specific information on the reproductive state as well as individual identity cues and social organization [34].
Rocky substrates and animal trails were not preferred scent marking sites for red foxes, unlike in felids, especially solitary snow leopard, where scrape marking and scent marking and disproportionately focused on these areas [35]. The distribution of scats in different habitats may be used to infer dispersal distances, such as the fox movements reported by Walton, et al. [36], ranging from 132 to 1036 km. Furthermore, it is also possible that there is a competition of signal post resources with conspecifics (opposite sexes) or heterospecifics (prey species) [37].
Although animals may use any of their diverse sensorial channels to obtain this information, most mammals are nocturnal and live in grassland. As a result, olfactory communication plays a principal role [3]. The seeds of food plants such as Hippophe rhamnoids, Prunus armeniaca, Morus nigra, Elaeagnus angustifolia and Rosa webbiana were found in fox faeces and were, therefore, also considered. We also recorded that fox commonly ingests fleshy fruits and are thus likely to be important seed dispersers.
This study forms a baseline from which to explore predator-prey interactions and fox resource selection at different spatial scales. Foxes focus their activities near den sites throughout the breeding season and frequently visit dens at other times of the year, hence den location is an essential consideration in managing human-fox conflict and protecting natural prey species.
Acknowledgments
We thank, Jinzhe Qi, Xin Liang, Henry and Nathan Roberts, friends in Feline Research Center of National Forestry and Grassland Administration for their useful comments. This work was supported by Fundamental Research Funds for the Central Universities (2572017PZ14) and NSFC (31872241; 31572285).
References
-
Johnson RP (1973) Scent marking in mammals. Animal Behaviour 21(3): 521-535.
-
Kavaliers M, Choleris E, Pfaff DW (2005) Recognition and avoidance of the odors of parasitized conspecifics and predators: differential genomic correlates. Neurosci Biobehav Rev 29(8): 1347-1359.
-
Monclús R, Arroyo M, Valencia A, De Miguel FJ (2009) Red foxes (_Vulpes vulpes_) use rabbit (_Oryctolagus_ _cuniculus_) scent marks as territorial marking sites. Journal of ethology 27(1): 153-156.
-
Banks PB, Daly A, Bytheway JP (2016) Predator odours attract other predators, creating an olfactory web of information. Biol lett 12 (5): 1-4.
-
Barja I, De Miguel F, Bárcena F (2001) Distribución espacial de los excrementos de zorro rojo (_Vulpes_ _vulpes_ Linnaeus, 1758) en los Montes do Invernadeiro (Ourense). Galemys 13: 171-178.
-
Burst TL, Pelton MR (1983) Black bear mark trees in the Smoky Mountains. Bears: Their Biology and Management 5: 45-53.
-
Barja I (1996) Estudio comparado del comportamiento en cautividad de leones del Atlas (Panthera leo leo) y tigres siberianos (Panthera tigris altaica)-Memoria de licenciatura. Universidad Autónoma de Madrid (in Spanish).
-
Bowyer RT, Ballenberghe VV, Rock KR (1994) Scent marking by Alaskan moose: characteristics and spatial distribution of rubbed trees. Canadian Journal of Zoology 72(12): 2186-2192.
-
Ralls K (1971) Mammalian scent marking. Science 171(3970): 443-449.
-
Monclús R, Miguel F (2003a) Senalización y respuesta a intrusos en el zorro rojo (Vulpes vulpes). Ediciones de la Universidad Autónoma de Madrid, Madrid.
-
Monclús R, De Miguel F (2003b) Distribución espacial de las letrinas de conejo (Oryctolagus cuniculus) en el monte de Valdelatas (Madrid). Galemys 15: 157-165.
-
Nowak RM, Walker EP (1999) Walker's Mammals of the World. JHU Press.
-
Ewer RF (1998) The carnivores. Cornell University Press.
-
Macdonald DW (2004) Dramatis personae. Wild Canids-an introduction and dramatis personae. The biology and conservation of wild canids pp: 4-35.
-
Meia J, Weber J (1992) Characteristics and distribution of breeding dens of the red fox (Vulpes vulpes) in a mountainous habitat. Zeitschrift für Saugetierkunde 57: 137-143.
-
Saunders G, Coman B, Kinnear J, Braysher M (1995) Managing vertebrate pests: foxes. Australian Government Publ Service.
-
Macdonald DW (1983) The ecology of carnivore social behaviour. Nature 301(5899): 379-384.
-
Raza G, Mirza SN, Anwar M, Hussain I, Kha, SW, et al. (2015) Population and Distribution of Himalayan Ibex, Capra ibex sibrica, in Hushe Valley, Central Karakoram National Park, Pakistan. Pakistan Journal of Zoolog, 47(4): 1025-1030.
-
Roberts TJ (2005) Field guide to the large and medium-sized mammals of Pakistan. Oxford University Press Karachi.
-
Schneider MF (2001) Habitat loss, fragmentation and predator impact: spatial implications for prey conservation. Journal of Applied Ecology 38 (4): 720- 735.
-
Maheshwari A (2018) Foraging habits of the Red Fox Vulpes vulpes (Mammalia: Carnivora: Canidae) in the Himalaya, India. Journal of Threatened Taxa 10(10): 12418-12421.
-
Zaman M, Tolhurst BA, Zhu M, Jiang G 2019 Increased Flight Initiation Distance (FID) in Golden Marmots (Marmota caudata aurea) Responding to Domestic Dogs in A Landscape of Human Disturbance. Animals 9(9): 605.
-
Weston M, McLeod EM, Blumstein D, Guay PJ (2012) A review of flight-initiation distances and their application to managing disturbance to Australian birds. Emu-Austral Ornithology 112(4): 269-286.
-
Abbas Z, Khan SM, Alam J, Khan SW, Abbasi AM (2017) Medicinal plants used by inhabitants of the Shigar Valley, Baltistan region of Karakorum range- Pakistan. Journal of ethnobiology and ethnomedicine 13(1): 53.
-
Ralls K, Cypher BL (2002) Use of Night Vision Technology to Study Kit Fox Behavior. Transactions of the Western Section of the Wildlife Society 38: 27- 28.
-
Smith DA, Ralls K, Hurt A, Adams B, Parker M, et al. (2003) Detection and accuracy rates of dogs trained to find scats of San Joaquin kit foxes (Vulpes macrotis mutica). Animal Conservation 6(4): 339-346.
-
de Migule FJ, Valencia A, Arroyo M, Monclus R (2009) Spatial distribution of scent marks in the red fox (Vulpes vulpes L.): do red foxes select certain plants as signal posts. Pol J Ecol 57 (3): 605-609.
-
Vogt K, Zimmermann F, Kölliker M, Breitenmoser U (2014) Scent-marking behaviour and social dynamics in a wild population of Eurasian lynx Lynx lynx. Behavioural processes 106: 98-106.
-
Wang ZH, Wang XM, Chmura A (2008) Den Habitat Characteristics of Tibetan Foxes (Vulpes ferrilata) in Shiqu County, Sichuan Province, China. Zoological Studies 47(4): 445-454.
-
Mahmood S, Mahmood T, Rais M, Qureshi I, Nadeem M (2010) A comparative study on the populations and habitats of the grey francolin Francolinus pondicerianus and the black francolin Francolinus francolinus in Lehri Nature Park, Punjab, Pakistan. Podoces 5(1): 42-53.
-
Schoon G (1996) Scent identification lineups by dogs (Canis familiaris): experimental design and forensic application. Applied Animal Behaviour Science 49(3): 257-267.
-
Dawley EM (1984) Recognition of individual, sex and species odours by salamanders of the Plethodon glutinosus-P. jordani complex. Animal Behaviour 32(2): 353-361.
-
Punjabi GA, Chellam R, Vanak AT (2013) Importance of native grassland habitat for den-site selection of Indian foxes in a fragmented landscape. PloS One 8 (10): e76410.
-
Vogt K, Boos S, Breitenmoser U, Kölliker M (2016) Chemical composition of Eurasian lynx urine conveys information on reproductive state, individual identity, and urine age. Chemoecology 26(6): 205-217.
-
Macri AM, Patterson-Kane E (2011) Behavioural analysis of solitary versus socially housed snow leopards (Panthera uncia), with the provision of simulated social contact. Applied Animal Behaviour Science 130(3-4): 115-123.
-
Walton Z, Samelius G, Odden M, Willebrand T (2018) Long-distance dispersal in red foxes Vulpes vulpes revealed by GPS tracking. European Journal of Wildlife Research 64(6): 64.
-
Zeus VM, Puechmaille SJ, Kerth G (2017) Conspecific and heterospecific social groups affect each other's resource use: a study on roost sharing among bat colonies. Animal behaviour 123: 329-338.
- In Situ Evaluation of the Anthelmintic Effect of the Aqueous Extract of Syzygium aromaticum (L) Merr and Perry on Bovine Strongyles
- Successful in Vitro Embryo Production with Oocytes Aspirated from Live White-Tailed Deer (Odocoileus Virginianus Texanus) Donors under Captivity in Northeast Mexico
- Bangladeshi Finches with their Evolutionary Thoughts (Aves: Passeriformes)
- Note on the Survival Status of Przewalski’s Horse, Equus ferus przewalskii (Perissodactyla: Equidae)
- In-Situ and Ex-Situ Protection of White-Breasted Waterhen (Amaurornis Phoenicurus) (Pennant, 1769) (Aves: Rallidae)
- Meat Examination in the Laboratory, the Acceptability and the Human Health