Surviving the Cold: A Case Study in Competition between Native and Invasive Fish for Overwinter Habitat Use
Invasive species management can be challenging or even impossible, especially in aquatic environments. To determine if the tropical origins of an invasive species affected where it could be targeted, we constructed a study to compare winter behavior of a tropical invasive to a temperate weather native species in Louisiana. The winter activity and cold tolerance of ten invasive Rio Grande Cichlids, Herichthys cyanoguttatus and ten native bluegill, Lepomis macrochirus was observed using an outdoor pond with a dual antenna PIT tag array and an artificial culvert created inside of the pond. All fishes had PIT tags injected into their lateral musculature. Both species were active at temperatures below 10°C and all twenty fish survived temperatures as low as 7°C. Even though the invasive H. cyanoguttatus is a member of a tropical family of fishes, activity and even feeding behavior of both species was similar up to 13°C, but H. cyanoguttatus individuals were more active than bluegill at higher temperatures. Both species appeared to use the culvert during cold fronts, demonstrating that the behavior of both species during cold weather were remarkably similar. This study illustrates the thermal tolerance abilities of H. cyanoguttatus in New Orleans and also demonstrates their potential for invading other cold-water habitats.
Introduction
Invasive species are damaging to ecosystems and cost over $100 billion annually [1]. Invasive species control is typically a challenging task, but there are many success stories involving elimination or great reduction in numbers of invasive species [2]. Understanding the biology of the invasive species is key to many of these success stories. Sea lamprey pheromones and sterile male insects are two examples where knowledge of the invasive species’ biology aided in control or elimination of the species [3, 4].
The Rio Grande Cichlid (Herichthys cyanoguttatus) is an invasive fish species that has been present in New Orleans for decades [5, 6]. The species is native to northeast Mexico and southern Texas from the Rio Conchos to southern drainages of the Rio Grande River [7]. In addition to Louisiana, this species has been introduced beyond its native range into drainages in both Texas and Florida (Fuller et al., 1999). Concerns over the continued spread of H. cyanoguttatus throughout these invaded habitats are driven by its salinity tolerance and its relatively rapid spread throughout the city and adjacent areas, including new parishes in the last decade [8, 9].
One potential limiting factor to the further spread of H. cyanoguttatus is that cichlids have been shown to have difficulty with cold tolerance [10, 11]. Most of the cichlids tested under laboratory conditions fare poorly, exhibiting extreme responses, loss of equilibrium and death, when the temperatures approach the 10-degree Celsius mark. However, H. cyanoguttatus is the northern-most species of naturally occurring cichlid in North America, therefore the cold tolerance of this species is of interest as it is the most likely of any cichlid species to find success as invader in more temperate latitudes, especially if climate change creates more suitable environments for invasions [12]. Siemien and Stauffer (1989) found that H. cyanoguttatus could survive at unusually low temperatures (7 degrees Celsius) for cichlids [13].
If it can be determined that there are thermal refuges being used, these refuges could be managed in a way to further the impact of cold winters. This and other cichlid species have been more successful in disturbed habitats versus natural ones. This may be due to lower relative temperatures experienced outside of urban environments, either because of heat generated in the city or thermal refuges not available in natural habitats. Buildings in general and concrete in the canals specifically consist of materials known to retain heat [14]. Overpasses in New Orleans and areas with more concrete held heat better than natural bayous nearby, as observed when temperature data loggers were deployed in both areas (Lorenz, pers obs).
Some behavior by H. cyanoguttatus in the fall months also indicates a shift in habitat use. Herichthys cyanoguttatus is easily observed grazing and breeding in late spring and summer months. This is represented clearly by the gut contents of canal dwelling cichlids in particular, which almost entirely consist of algae (Lorenz pers obs). In the fall the cichlids start to form loose shoals that move about the canals until late fall when no cichlids are seen at all. This sort of behavior (grazing, shoaling) is also observed in a close relative, H. tamasopoensis [15]. The virtual disappearance of the cichlids in the canals has created the hypothesis that cichlids are shifting habitat use to culverts and overpasses and these invasive fish could be targeted at these locations if they use them during known times and/or temperatures and if they use these areas differently than native fishes.
Our experiment examined H. cyanoguttatus activity compared to native fish of similar size (Lepomis macrochirus), at different overwintering temperatures in an artificial outdoor mesocosm with a culvert. We constructed the mesocosm and added tagged fish and a PIT tag array to answer the two following questions: 1) do activity patterns of invasive and native species vary at different temperatures? and 2) do invasive cichlids differ from native bluegill in activity level at low temperatures?
Materials and Methods
Mesocosm Construction
The mesocosm was constructed using a frame of cinder blocks and mortar. The dimensions of the pond were 4.5 m long and 1.8 m wide and 0.6 m high. With a pond liner installed, the water filled to a height of approximately 0.4 to 0.5 m in depth. 2.5 centimeters of sand was used as substrate and 6 cinderblocks were distributed evenly and on end to provide cover in the two thirds of the pond that were open. The other third consisted of the culvert, which was created by balancing six 61cm X 61 cm X 3 cm pavement stones. Three of these stones covered the entire width of the pond with less than 5 cm total overlap of the stones and two rows of these stones then created a covered area of approximately 1.6 m by 1.2 m. These pavement stones were supported to a height of 0.5 to 0.6 meters by cinder blocks and bricks underneath (Figure 1).
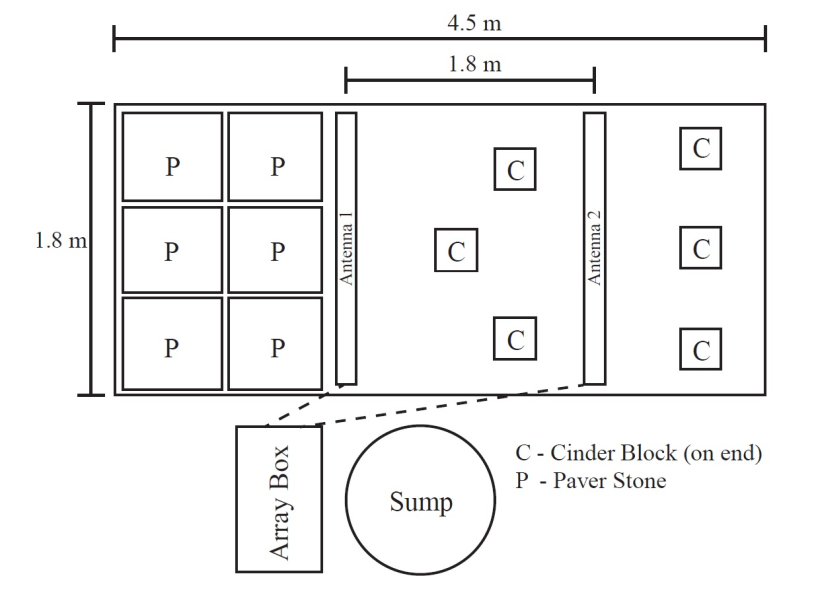
Filtration was performed in a sump created by a 416 liter stock tank, connected by 2.5 cm diameter hoses to the pond. A UV filter (Tetrapond UVC-9) and sump pump (Rio Plus Powerhead 1700) treated and recirculated the water back to the pond from the sump. Fish were fed Hikari food sticks ad libitum three times a week.
Tagging
Eleven H. cyanoguttatus and eleven L. macrochirus were collected from City Park in New Orleans using either hook- and-line, electrofishing, or cast net. Each fish was given one week of acclimation in a 416 liter tank with an air driven sponge filter. Individual adult fish were tagged with half- duplex (HDX) passive integrated transponder (PIT) tags while sedated by clove oil at 4mg/L concentration (2 ml added to 5 liters of water) and an airstone was added to aid in respiration during the process [16, 17]. Fish were tagged intramuscularly after fish were sedated (always in less than 2 minutes) and placed in fresh water less than 5 minutes after sedation (less than 7 minutes after initial sedation) [18]. Recovery appeared to take less than 10 minutes for all individuals and all individuals survived the duration of the experiment.
Array
To investigate the activity of tagged L. macrochirus and H. cyanoguttatus, a PIT tag array was constructed to record any movement activity [19, 20]. This array is a monitoring station that records half-duplex (HDX) passive integrated transponder (PIT) tags using a multiplexer self-contained Oregon RFID monitoring station (Multi-Antenna HDX reader, Oregon RFID) powered by one 12V deep cycle marine battery enclosed in a field box to for protection from the climate. The reader is connected to two coiled antennas encased in 5.08 cm diameter PVC sheaths anchored in the mesocosm. Each antenna consisted of a continuous length of 14 AWG stranded copper wire wrapped twice inside of a PVC sheath. Inductance of each antenna was measured with the Extech LCR Meter (Model 380193) in order to calibrate within the 8-80 uH range. Each antenna achieved a reading of 20.2 uH which is well within the ideal inductance range needed for calibration with the Oregon RFID components used. After calibration was complete the antennas were sealed, waterproofed and anchored to the bottom of the artificial culvert. The PVC sheathed antennas were placed in two locations, one next to the culvert and one 1.8 meters away from the first antenna. For the purposes of this study, the antenna by the culvert was labeled “Antenna 1” and the antenna farther away was labeled “Antenna 2”. Both antennas were 1.35 meters from the ends of the mesocosm.
Throughout the experiment our antennas held amperage levels adequate for maintenance of a complete electromagnetic field throughout our test field but weekly checks of the voltage and reader levels ensured stable voltage levels and tuning range. Batteries were exchanged every other day to maintain a voltage above 11V. The data in the array was periodically uploaded onto a laptop during one of the battery exchanges. Hobo temperature data loggers were deployed in shade and sun and within the culvert to record average water temperatures for analyses.
Fish Activity
Measuring activity directly is difficult when location data is limited to when the animal passes a specific location. We had two stations (antenna 1 and antenna 2) that measured the entire cross section (1.8 m apart) of mescosm and felt that we could use movement as a proxy for activity (Figure 1). For this study, a movement was defined as when a fish was detected at a different location than its previous detection within one hour. To test for differences in movement between species and with respect to temperature, we employed two different generalized linear mixed models (GLMM) using R and the lme4 package [21]. For both models, temperature, species and an interaction between temperature and species were treated as fixed effects, while individual and date were treated as random effects. To better understand the relationship between whether or not a fish moved in a given hour, movement or non-movement was treated as a binary response for a binomial GLMM. To better understand the relationship of hourly movements, hourly counts were used as a response variable in a Poisson GLMM. For both models, Bayesian Information Criteria (BIC) was used to select the most parsimonious model [22].
Results
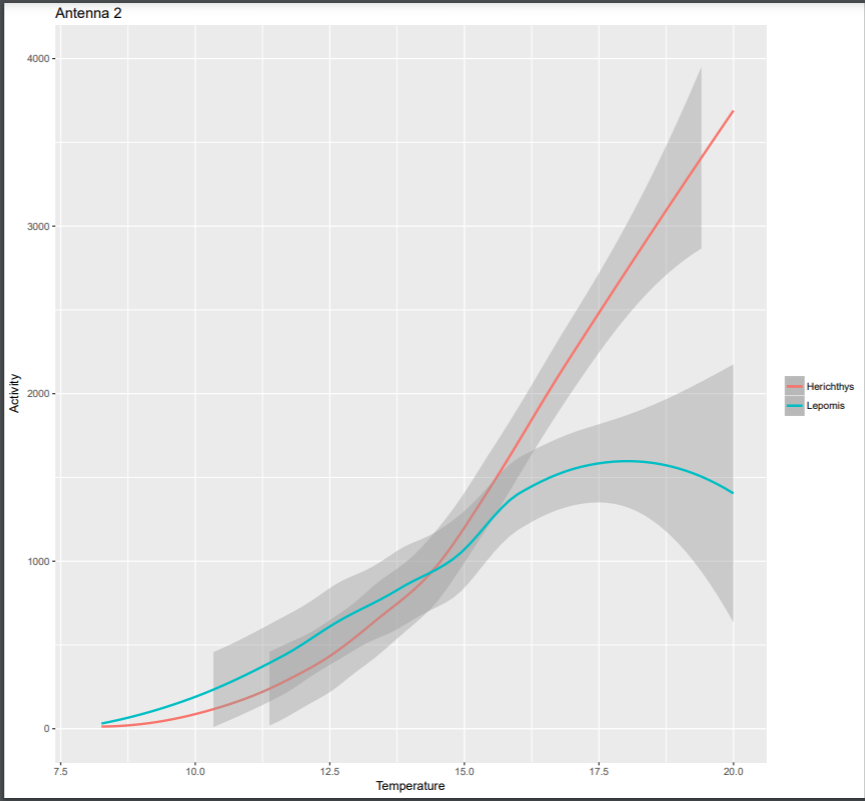
All fish survived the duration of the experiment with no loss to temperatures as cold as 7 degrees Celsius and no loss to predation due to being outdoors. Growth was minimal (not measureable) for all individuals, despite feeding even during the coldest months. The null model was found to be the most parsimonious for both the binomial and the Poisson GLMMs. We found a significant positive correlation between movements and temperature between species. There was a significantly larger slope between movements and temperature for H. cyanoguttatus (Figure 2). There was also a stronger positive correlation between movements and temperature for H. cyanogutatus than L. macrochirus. Individual “pings” of activity rarely exceeded 2000 at any temperature for L. macrochirus, while it approached 4000 pings at higher temperatures (20 degrees Celsius) for H. cyanoguttatus. There was no significant difference in overall mean movements between species, however when controlling for the effect of temperature, mean hourly movements were higher for H. cynagutattus (Tables 1 & 2). Activity did not appear to differ at the lowest temperatures between the two species.
| Estimate | SE | z | p | |
|---|---|---|---|---|
| Intercept | -1.74495 | 0.13759 | -12.683 | <0.001 |
| Species | 0.34822 | 0.17797 | 1.957 | 0.0504 |
| Temperature | 0.36665 | 0.02575 | 14.237 | <0.001 |
| Species:Temperature | 0.22499 | 0.02245 | 10.024 | <0.001 |
Table 1: Binomial BIC suggested that the null model was the most parsimonious Binomial GLMM and those results are presented here.
| Estimate | SE | z | p | |
|---|---|---|---|---|
| Intercept | -0.88054 | 0.20549 | -4.285 | <0.001 |
| Species | 0.40645 | 0.25933 | 1.567 | 0.117 |
| Temperature | 0.30262 | 0.01638 | 18.478 | <0.001 |
| Species: Temperature | 0.29485 | 0.01205 | 24.459 | <0.001 |
Table 2: Poisson BIC suggested that the null model was the most parsimonious Poisson GLMM and those results are presented here. A
Conclusion
The results of the movement analyses suggest that H. cyanoguttatus had similar movements with respect to temperature as L. macrochirus, despite differences in natural history and range. L. macrochirus is a more temperate fish and its current distribution extends much higher latitudinally than H. cyanoguttatus, a more tropical cichlid. One significant difference between species observed in this study is a significantly larger slope between temperature and movements was observed for H. cyanoguttatus, but there was not a significant difference in mean movements between species. This indicates that H. cyanoguttatus maintained a similar activity level as bluegill at low temperatures but became more active than bluegill at warmer temperatures.
While there was a difference between activity between H. cyanoguttatus and L. macrochirus, it was not at lower temperatures. Both species stayed active at low temperatures and both species used the culvert and were not observed outside of the culvert during the coldest day of every cold front that passed through. Cold tolerance for both species clearly goes as low as 7 degrees Celsius, which is lower than what is thought possible for most cichlids, and was reported as the lower lethal temperature for this species [13]. This has serious implications on the survivorship of this species at low temperatures and the possible places that could be invaded. It is possible that negative effects were incurred but unobserved, however these cichlids were used in additional experimental studies suggesting this is not the case (Ron Oldfield pers obs).
Activity at Antenna 2 for higher temperatures (above 16 degrees Celsius) was greater for H. cyanoguttatus than for L. macrochirus (Figure 2), and this has implications as well. This increased activity of the invasive versus the native can mean fewer foraging or spawning habitat for bluegill. This supports previous observations that H. cyanoguttatus is successful at invading new territories, and defending those territories, while L. macrochirus is not struggles to invade cichlid territory [23]. The timing of the dominance of Antenna 2 is also disconcerting for L. macrochirus breeding success because nests of bluegill are observed at higher temperatures in the city and early breeding to avoid cichlid dominance could mean lower survival of sunfish larvae [24].
Few studies have used PIT tag arrays in mesocosms to observe fish movement [25]. Even fewer have observed seasonal movement [26]. This study is a first to observe overwintering behavior of an invasive species in comparison to native species to the climate being examined. Interestingly, the cold appeared to have no negative effects on the invasive, raising the question of how to manage such a species.
In addition to low temperatures, salinity has also been shown to not be a barrier [8]. It is possible that such extremes limit growth and reproduction or that other factors limit their spread, such as unsuitable feeding or breeding habitats outside of the urban canals. Management of Rio Grande cichlids in general is difficult with their high rate of reproduction and parental care typical of cichlids. Targeting culverts in the winter for poisoning with rotenone or electrofishing may still be effective, even though natives will likely also be using these habitats. Native species can be stocked or not removed (in the case of electrofishing). Future field studies are needed to examine if these mesocosm observations are also observed in wild fish overwintering behavior.
Acknowledgements
The research was supported by the Barataria-Terrebonne National Estuary Program and the Nekton Research Laboratory at the University of New Orleans. Special thanks to T. Zimmerman for support with the electronics. All aspects of the experiment were approved by the University of New Orleans IACUC, Protocol # UNO-IACUC 10-003.
References
-
Pimentel D, Zuniga R, Morrison D (2005) Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ 52(3): 273-288.
-
Simberloff D (2008) We can eliminate invasions or live with them. Successful management projects. Ecological Impacts of Non-Native Invertebrates and Fungi on Terrestrial Ecosystems, pp: 149-157.
-
Sorensen PW, Hoye TR (2007) A critical review of the discovery and application of a migratory pheromone in an invasive fish, the sea lamprey Petromyzon marinus L. J Fish Bio 71: 100-114.
-
Klassen W, Curtis CF (2005) History of the sterile insect technique. Sterile insect technique, pp: 3-36.
-
Fuentes GN, Cashner RC (2002) Rio Grande cichlid established in the Lake Pontchartrain drainage, Louisiana. The Southwestern Naturalist 47(3): 456-459.
-
O’Connell MT, Cashner RC, Fuentes GN (2002) Application of a diffusion model to describe a recent invasion; observations and insights concerning early stages of expansion for the introduced Rio Grande cichlid, Cichlasoma cyanoguttatum, in southeastern Louisiana. Aquat. Invaders 13(4): 13-21.
-
Baird SF, Girard C (1853) Descriptions of new species of fishes collected by Mr. John H. Clark, on the US and Mexican boundary Survey, under Lt. Col. Jas. D. Graham. Proc Acad Nat Sci Phil 4: 387-390.
-
Lorenz OT, Riccobono SA, Smith P (2016) Effects of salinity on the survival and aggression of the invasive Rio Grande cichlid (Herichthys cyanoguttatus). Marine and Freshwater Behaviour and Physiology 49(1): 1-8.
-
Lorenz OT, O’Connell MT (2011) Establishment and post- hurricane survival of the non-native Rio Grande cichlid (Herichthys cyanoguttatus) in the greater New Orleans metropolitan area. Southeastern Naturalist 10(4): 673- 686.
-
Shafland PL, Pestrak JM (1982) Lower lethal temperatures for fourteen non-native fishes in Florida. Environ Biol Fishes 7(2): 149-156.
-
Schofield PJ, Loftus WF, Kobza RM, Cook MI, Slone DH (2010) Tolerance of nonindigenous cichlid fishes (Cichlasoma urophthalmus, Hemichromis letourneuxi) to low temperature: laboratory and field experiments in south Florida. Biological Invasions 12(8): 2441-2457.
-
Hellmann JJ, Byers JE, Bierwagen BG, Dukes JS (2008) Five potential consequences of climate change for invasive species. Conserv Biol 22(3): 534-543.
-
Siemien MJ, Stauffer JR (1989) Temperature preference and tolerance of the spotted tilapia and Rio Grande cichlid. Archiv fur Hydrobiologie 115(2): 287-303.
-
Lee T, Hawes DW, Banu DT, Feldman D (2000) Control aspects of latent heat storage and recovery in concrete. Solar Energy Materials & Solar Cells 62(3): 217-237.
-
Azas A, Juan M (1993) Herichthys tamasopoensis n. sp., a new cichlid from México (Pisces, Cichlidae). Cichlids Yearbook 3: 65-70.
-
Peake S (1998) Sodium bicarbonate and clove oil as potential anesthetics for nonsalmonid fishes. North American Journal of Fisheries Management 18(4): 919- 924.
-
Javahery S, Nekoubin H, Moradlu AH (2012) Effect of anaesthesia with clove oil in fish. Fish Physiol. Biochem 38(6): 1545-1552.
-
Hopko M, Zakes Z, Kowalska A, Partyka K (2010) Impact of intraperitoneal and intramuscular PIT tags on survival, growth, and tag retention in juvenile pikeperch, Sander lucioperca (L.). Fisheries & Aquatic Life 18(2): 85-92.
-
Barbour AB, Adams AJ, Behringer DC, Yess TANNER, Wolfe RK (2011) PIT Tag Antennae Arrays as fishery monitoring tools in tropical environments pp: 1-7.
-
Barbour AB, Adams AJ, Yess T, Behringer DC, Wolfe RK (2012) Comparison and cost-benefit analysis of PIT tag antennae resighting and seine-net recapture techniques for survival analysis of an estuarine-dependent fish. Fisheries Research 121: 153-160.
-
Bates D, Machler M, Bolker BM, Walker SC (2014) Fitting linear mixed-effects models using lme4. Journal of Statistical Software, pp: 1-51.
-
Schwarz G (1978) Estimating the dimension of a model. Ann Statist 6(2): 461-464.
-
Lorenz OT, O’Connell MT, Schofield PJ (2010) Aggressive interactions between the invasive Rio Grande cichlid (Herichthys cyanoguttatus) and native bluegill (Lepomis macrochirus), with notes on redspotted sunfish (Lepomis miniatus). Journal of Ethology 29(1): 39-46.
-
Garvey JE, Herra TP, Leggett WC (2002) Protracted reproduction in sunfish: the temporal dimension in fish recruitment revisited. Ecol App 12(1): 194-205.
-
Nagrodski A, Murchie KJ, Stamplecoskie KM, Suski CD, Cooke SJ (2013) Effects of an experimental short‐term cortisol challenge on the behaviour of wild creek chub Semotilus atromaculatus in mesocosm and stream environments. J Fish Biol 82(4): 1138-1158.
-
Clark SR, Schaefer JF (2016) Ecological influences on the local movement dynamics of the blackspotted topminnow, Fundulus olivaceus. Behav Ecol Sociobiol 70(4): 557-567.
- In Situ Evaluation of the Anthelmintic Effect of the Aqueous Extract of Syzygium aromaticum (L) Merr and Perry on Bovine Strongyles
- Successful in Vitro Embryo Production with Oocytes Aspirated from Live White-Tailed Deer (Odocoileus Virginianus Texanus) Donors under Captivity in Northeast Mexico
- Bangladeshi Finches with their Evolutionary Thoughts (Aves: Passeriformes)
- Note on the Survival Status of Przewalski’s Horse, Equus ferus przewalskii (Perissodactyla: Equidae)
- In-Situ and Ex-Situ Protection of White-Breasted Waterhen (Amaurornis Phoenicurus) (Pennant, 1769) (Aves: Rallidae)
- Meat Examination in the Laboratory, the Acceptability and the Human Health