Mutations in the GDF8 Gene in Double-Muscled Cattle: Mechanisms of Action and Relevance for Livestock Production
Myostatin (MSTN) has the function of limiting muscle growth, keeping satellite cells quiescent and acting on cell development, proliferation, differentiation, adhesion, migration and apoptosis. The mutation in the GDF8 gene proved to be advantageous as it presents some important attributes for the production of beef cattle, such as: gains in feed conversion and meat quality, making the meat leaner and more tender, resulting in intense selection for use in crossing programs in Brazil and around the world. We carried out a literature review on the mutation in the GDF8 gene in different breeds of Double Muscled cattle. We evaluated the frequency of polymorphisms present in research and observed that Indel c. 818 and MSTN-F94L were the markers most used in research. The MSTN-F94L polymorphism influenced musculature in the Jersey-Limousin backcross population. Furthermore, the polymorphism is strongly associated with the additive effect on birth weight, weaning weight and yearling weight, in addition to increasing the ease of direct birth in pure or mixed breeds. GDF8 gene polymorphisms can be used to improve performance traits in beef cattle. However, the influence of these markers needs to be determined in each herd.
Introduction
Beef livestock farming currently stands as one of the pivotal sectors within the Brazilian and global economies. Producers are increasingly prioritizing the restructuring of the beef cattle production chain to align with the stringent meat quality standards demanded by both domestic and international consumer markets Al-Samarai FR, et al. [1].
Consequently, there’s a pressing need to utilize animals exhibiting high productivity and suitability for breeding systems to foster the advancement of this activity on a global scale EMBRAPA [2]. Genetic improvement in beef cattle has been employed for decades as a method to enhance production efficiency and ensure the palatability of meat, a characteristic directly associated with tenderness Miller S [3]. Enhancing muscle mass in cattle stands as one of the primary objectives in genetic improvement programs for both researchers and producers Grigoletto L, et al. [4]. In this context, double muscle syndrome (MD) emerges as a mutation observed in various cattle breeds worldwide Konovalova E, et al. [5], Prihandini PW, et al. [6], first described in the 19th century by Culley Oliver WM, et al. [7], Karim L, et al. [8]. This syndrome is characterized by muscle hypertrophy, particularly in the hind region of cattle, resulting in a higher muscle-to-bone and muscle-to-fat ratio Menissier F [9], Shahin KA, et al. [10].
In various cattle breeds, the mutation results in the loss of function of the Myostatin protein gene, also referred to as Growth Differentiation Factor 8 (GDF8), which dictates the MD phenotype Haruna IL, et al. [11]. This mutation is typically triggered by a premature stop codon that renders the protein inactive and dysfunctional Karim L, et al. [8], Marchitelli C, et al. [12]. Hence, by defining parameters for genetic analysis of mutations in the GDF8 gene in animals with MD, it becomes feasible to establish the genomic profile associated with enhanced muscular development and productive performance Aiello D, et al. [13]. The mutation in the GDF8 gene has demonstrated advantages, including improved feed conversion efficiency and meat quality, leading to significant selection pressure for its integration into breeding programs both in Brazil and globally Arthur PF [14], Wheeler TL, et al. [15]. Thus, the present study aimed to conduct an integrative literature review spanning from 2014 to 2024, utilizing databases such as PubMed, Google Scholar, and Scielo. The focus was on investigating the mutation in the GDF8 gene across various breeds of double-muscled cattle. Assessing the role of this protein is crucial for determining optimal performance and potential in meat production. Moreover, it aids in the selection of superior beef cattle breeds for inclusion in breeding and genomic selection programs.
Double Musculature in Cattle
There is a persistent trend to improve carcass quality in specialty beef breeds Zheng Y, et al. [16]. As a result, a higher yield of lean meat is desirable for the consumer market Vanek Jk, et al. [17]. According to data from the Brazilian Association of Meat Exporting Industries-ABIEC [18], in 2019, the Brazilian herd accounted for an average of 213.68 million heads of cattle, with a record of slaughtering 43.3 million heads of cattle, being one of the main producers and exporters of beef in the world. In 2016, the national herd reached the mark of 218.23 million heads of cattle IBGE [19]. Across the globe, breeds selected for production and research prioritize both productive and reproductive potential. In certain cattle breeds exhibiting double musculature, various mutations leading to the loss of function of the myostatin gene can be observed Csurhes T, et al. [20]. The Belgian Blue breed, originating in Belgium in the 1970s, is renowned for its remarkable muscularity, attributed to a loss-of-function mutation in the myostatin gene Grobet LJR, et al. [21]. Charlier C, et al. [22] successfully mapped the GDF8 gene to bovine chromosome 2.
The Aberdeen Angus breed currently enjoys prominence in the market owing to the quality of its meat and its adaptability to the Brazilian tropical climate. Weber T, et al. [23] reported that among taurine beef breeds, Angus accounted for the highest number of semen doses sold in Brazil (16.4% of the total) and the highest number of imported semen doses (67.5%). These findings underscore the breed’s significant role in the Brazilian meat production chain. The Piemontese breed stands as the predominant breed within the Italian beef herd, currently accounting for an average of 350,000 head of cattle Rural C [24]. These cattle are characterized by their substantial muscle mass, low fat deposition, reduced skeletal weight, low feed consumption, and efficient feed conversion Fiems LO [25]. Primarily selected for its genetic superiority in meat production and favorable qualities for both direct and maternal birthing Albera A, et al. [26].
Another breed of significant importance in the beef market is the Marchigiana breed, originating from Italy Mancini G, et al. [27]. Known for its muscular hypertrophy, Marchigiana cattle yield carcasses with consistent high performance and minimal variation between individuals. With a notable emphasis on muscle mass and minimal fat and bone content, these attributes are highly valued economically Sarti FM, et al. [28], Sorbolini S, et al. [29]. The mutation responsible for the double musculature phenotype in this cattle population results from a G → T transversion at position 874, leading to a stop codon in the C-terminal bioactive domain, known as E291X Cappucio I, et al. [30], Marchitelli C, et al. [12]. Myostatin Protein (MSTN) MSTN, discovered in 1997 by Lee and McPherron (Rodgers BD, et al. [31]), spurred researchers to deepen their understanding of its function, particularly its potential for inhibition against conditions characterized by significant muscle mass loss. Concurrently, a team of researchers at the Johns Hopkins School of Medicine first described the protein as a negative regulator of muscle mass McPherron AC, et al. [32]. MSTN belongs to the transforming growth factor TGF-β supercytokine family, with its primary function being to restrict muscle growth by maintaining satellite cells in a quiescent state. It exerts influence over various cellular processes including development, proliferation, differentiation, adhesion, migration, and apoptosis Lee SJ [33]. Unlike other members of the TGF family, MSTN is predominantly expressed in the skeletal muscle of both adult and developing individuals Carlson CJ, et al. [34]. During the embryonic period of animal development, particularly during the myogenesis process (Figure 1a), the GDF8 gene plays a pivotal role in regulating myoblast proliferation through its interaction with the Activin type II2 receptor (ActIIb) (Figure
1b). This interaction inhibits somite activation by repressing the transcription factor Pax3, subsequently hindering myoblast proliferation by inhibiting the transcription factors MyoD and Myof5, and impeding myocyte differentiation into myotubes by suppressing myogenin and MRF4 transcription Wehling M, et al. [35].
Furthermore, during embryogenesis, myostatin is expressed by cells in the myotome and developing skeletal muscle, serving to regulate the final count of muscle fibers formed. In adulthood, myostatin exerts an inhibitory effect on skeletal muscle development, suppressing the expression of transcription factors crucial for the muscle formation cycle and preventing tissue hyperplasia Lee SJ [33].
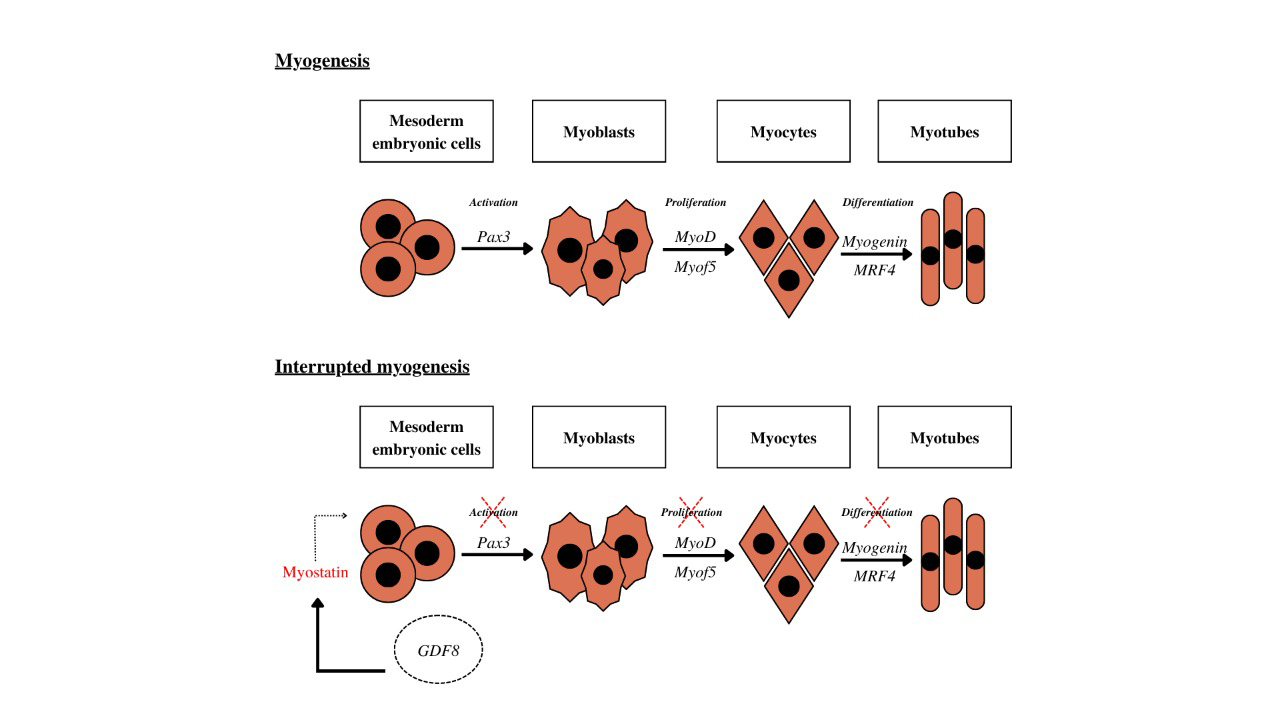
Figure 1: Stages of muscle tissue formation, or the myogenesis process, and mechanisms of gene action. a) Steps in muscle tissue formation, or the myogenesis process, without interference from gene expression regulation mechanisms. b) Stages of muscle tissue formation or the myogenesis process under the influence of myostatin and gene expression regulation mechanisms.
The mutation responsible for muscular hypertrophy (mh) is situated in the GDF8 gene, also known as the MSTN gene McPherron AC, et al. [32]. The myostatin gene has been precisely mapped to the distal end of chromosome 2 in cattle Grobet LJR, et al. [21]. The presence of the mutant mh allele in loss-of-function homozygotes consequently results in muscle hypertrophy. However, it’s worth noting that the mutation doesn’t necessarily have to be present in the homozygous condition for an effect to manifest Bellinge RH, et al. [36]. In a study conducted by Casas E, et al. [37], it was observed that animals inheriting a single copy of the mutant mh allele, either from a Belgian Blue or Piedmontese crossbreed, exhibited increased Longissimus muscle area and reduced external and intramuscular fat deposition compared to animals without any copy of the mutant mh allele. Bos taurus taurus was the first subspecies of cattle in which a mutation of the GDF8 gene was identified. A total of 20 different mutations have been identified in the GDF8 gene, with 14 located in the coding sequence and six in the intronic sequences of the gene Marchitelli C, et al. [12], Grobet LJR, et al. [21], Dunner S, et al. [38], Shibata M, et al. [39].
The first mutations of the myostatin gene in cattle were first documented in the Belgian Blue and Piedmontese breeds, both renowned for their substantial muscular development compared to conventional cattle. Mutations within the GDF8 gene have been identified across various species, including dogs Mosher DS, et al. [40], sheep Kijas JW, et al. [41], cattle Grobet LJR, et al. [21], pigs Stinckens A, et al. [42], and humans Schuelke M, et al. [43]. In this study, we examined the prevalence of polymorphisms reported in the literature and observed that Indel c. 818 Jakaria J, et al. [44] and MSTN-F94L Lee J, et al. [45] were among the most frequently utilized markers in research.
Association of Single Nucleotide Polymorphisms (SNPs) in the GDF8 Gene with the Productive Performance of Bovine Breeds Studies involving double-muscled animals have garnered significant attention from breeders and geneticists, particularly following the discovery of new scientific evidence linking the double-muscled phenotype to polymorphisms in the GDF8 gene in certain cattle. Jakaria J, et al. [44] investigated the presence of SNPs and 11 bp deletions in the coding region of the GDF8 gene, exploring their association with the double muscle phenotype in cattle crossbred with the Belgian Blue breed. Their study revealed the identification of four new SNPs and an 11 bp indel within the coding region of the GDF8 gene. The Belgian Blue cattle breed is renowned for its exceptional carcass yield. However, a correlation has been established between the mutation in the myostatin gene, which results in double musculature (mh/mh genotype) in Belgian Blue cattle, and susceptibility to psoroptic mange, a skin disease caused by the mite Psoroptes ovis Meyermans R, et al. [46], Vinet A, et al. [47] propose that the TG3811 polymorphism in the second intron of the myostatin gene accounts for the superior carcass and muscle yield observed in homozygous Blonde d’Aquitaine [G/G] breeds. Cattle carrying the TG3811 polymorphism exhibited lower intramuscular fat content compared to normal cattle. Hence, this mutation likely exerts a direct influence on lipid metabolism, alongside a notable impact on muscle fiber growth and differentiation.
Jakaria J, et al. [44] identified 8 SNPs within the coding region of the GDF8 gene, specifically: c.111G/C; rs523392653, c.267G/A; rs383271508, c.1077C/A; rs466598800; c.1083 T/C, and rs211583837. Additionally, they observed that crossbred individuals, Belgian Blue x Peranakan Ongole, predominantly exhibited point mutations in the GDF8 gene, followed by Peranakan Ongole cattle. In the same study, the authors detected a novel deletion at position 11 bp of exon 3, denoted as Indel c.818 (5’-ATGAACACTCC-3’; rs382669990), in Belgian Blue cattle. This deletion was not observed in Peranakan Ongole cattle, whereas in crossbred cattle (Belgian Blue x Peranakan Ongole), a heterozygous deletion (Indel 11bp) was noted. In the GDF8 gene, Sarti FM, et al. [28] identified a transversion from G to T in the Marchigiana breed. The “GG” homozygote exhibits a “normal” phenotype, while the “TT” homozygote displays a double musculature phenotype, albeit with a potential increase in mortality rate. Conversely, the heterozygous genotype can manifest double musculature without detrimental effects. Homozygous Marchigiana “TT” individuals are excluded from breeding programs, while heterozygous individuals are selected as breeders. Bimuscular carcasses typically exhibit a high carcass yield percentage and a low proportion of fat and bones Kobolak J, et al. [48]. In Ceccobelli S, et al. [49] findings, although no significant difference was observed between heterozygous and homozygous Marchigiana bulls, it was noted that heterozygous bulls achieved a final live weight at slaughter averaging 17 kg higher than monozygous bulls. Thus, concerning post-mortem carcass parameters, the observed average daily weight gain suggests an impact of the mutation. These findings align with the observed muscle mass growth attributed to the loss of function of the myostatin protein in Marchigiana beef cattle, resulting in increased muscle fiber count (hyperplasia), fiber enlargement (hypertrophy), and decreased subcutaneous and intermuscular fat deposition Bennett GL, et al. [50].
Cushman RA et al. [51] conducted a three-year assessment of the growth and reproductive indices of beef heifers to investigate the impact of the MSTN F94L polymorphism in the GDF8 gene on reproductive performance. Their findings indicated that the MSTN Leu allele was linked to reduced birth weight. Additionally, this allele was associated with a delayed onset of puberty, although it did not significantly affect the calving date. The MSTN F94L polymorphism shows promise for enhancing zootechnical performance related to weight gain in beef heifers without compromising fertility. However, further research is needed to ascertain its broader influence on cattle herd performance. Lee J, et al. [28] identified that individuals of the pure Limousin and mixed breed (1/2 Angus, 1/2 Limousin) possessing the MSTN F94L polymorphism in the 6 Mb region of BTA 2 (Bos taurus autosome) showed an association with additive effects on birth weight, weaning weight, and yearling weight, as well as an increased ease of direct birth. Furthermore, the study demonstrated the predictive accuracy achieved by incorporating the MSTN F94L polymorphism as either a random or fixed effect in genomic evaluations, compared to using other SNP markers in the BovineSNP50 BeadChip, thus enhancing prediction efficiency Lee SJ, et al. [52].
Conclusion
GDF8 gene polymorphisms hold potential for enhancing performance characteristics in beef cattle. Nonetheless, the impact of these markers must be assessed within each individual herd. Consequently, regarding the MSTN F94L marker, prior to its adoption for genetic selection decisions by beef producers, it is imperative to comprehend its association with reproductive performance within the cattle herd. In light of this, future studies are warranted to elucidate the mechanism of action underlying the MSTN F94L polymorphism, which remains to be fully understood.
References
-
Al-Samarai FR, Dawood TN, Luaibi OK, Al-Nedawi, AM (2021) Genetic Improvement of the Dairy Cattle in the Tropical Countries: A Review. International Journal of Science and Research Archive 3(2): 188-192.
-
Brazilian Agricultural Research Company (2018) Vision 2030: the future of Brazilian agriculture - EMBRAPA, Brasília-DF.
-
Miller S (2010) Genetic Improvement of Beef Cattle through Opportunities in Genomics. Revista Brasileira de Zootecnia 39: 247-55.
-
Grigoletto L, Ferraz JBS, Oliveira HR, Eler JP, Bussiman FO, et al. (2020) Genetic Architecture of Carcass and Meat Quality Traits in Montana Tropical® Composite Beef Cattle. Frontiers in Genetics 11: 123.
-
Konovalova E, Romanenkova O, Zimina A, Volkova V, Sermyagin A (2021) Genetic Variations and Haplotypic Diversity in the Myostatin Gene of Different Cattle Breeds in Russia. Animals (Basel) 11(10): 2810.
-
Prihandini PW, Primasari A, Aryogi A, Luthfi M, Hariyono DNH (2022) Genetic Polymorphisms of the 5’ Untranslated Regions of the HSP70 Gene in Indonesian Cattle Populations. Veterinary World 15(1): 168-172.
-
Oliver WM, Cartwright TC (1968) Double Muscling in Cattle: A Review of Expression, Genetics and Economic Implication. Agricultural Experiment Station Technical Report 12: 58.
-
Karim L, Coppieters W, Grobet L, Valentini A, Georges M (2020) Convenient Genotyping of Six Myostatin Mutations Causing Double-Muscling in Cattle using a Multiplex Oligonucleotide Ligation Assay. Animal genetics 31(6): 396-399.
-
Menissier F (1982) General Survey of the Effect of Double Muscling on Cattle Performance. Current Topics in Veterinary Medicine and Animal Science 16: 23-53.
-
Shahin KA, Berg RT (1985) Growth Patterns of Muscle, Fat and Bone, and Carcass Composition of Double Muscled and Normal Cattle. Canadian Journal of Animal Science 65(2): 279-293.
-
Haruna IL, Ekegbu UJ, Ullah F, Amirpour-Najafabadi H, Zhou H, et al. (2020) Genetic Variations and Haplotypic Diversity in the Myostatin Gene of New Zealand Cattle Breeds. Gene 740: 144400.
-
Marchitelli C, Savarese MC, Crisà A, Nardone A, Marsan PA, et al. (2003) Double Muscling in Marchigiana Beef Breed is caused by a Stop Codon in the Third Exon of Myostatin Gene. Mamm Genome 14(6): 392-395.
-
Aiello D, Patel K, Lasagna E (2018) The Myostatin Gene: An Overview of Mechanisms of Action and its Relevance to Livestock Animals. Animal Genetics 49(6): 505-519.
-
Arthur PF (1995) Double Muscling in Cattle: A Review. Australian Journal of Agricultural Research 46(8): 1493- 1515.
-
Wheeler TL, Shackelford SD, Casas E, Cundiff LV, Koohmaraie M (2001) The Effects of Piedmontese Inheritance and Myostatin Genotype on the Palatability of Longissimus Thoracis, Gluteus Medius, Semimembranosus, and Biceps Femoris. Journal of Animal Science 79(12): 3069-3074.
-
Zheng Y, Ma H, Zheng Y, Wang Y, Zhang B, et al. (2012) Site-Directed Mutagenesis of the Myostatin Gene in Ovine Fetal Myoblast Cells in Vitro. Research in Veterinary science 93(2): 763-769.
-
Vanek JK, Watts MJ, Brester GW (2008) Carcass Quality and Genetic Selection in the Beef Industry. Journal of Agricultural and Resource Economics 33(3): 349-363.
-
Brazilian Association of Meat Exporting Industries (2020) Beef Report: Profile of Livestock in Brazil. ABIEC, Brasilia-DF.
-
IBGE (2017) Municipal Livestock 2016: Central-West accounts for 34.4% of the country’s cattle herd.
-
Csurhes T, Szabo F, Hollo G, Miko E, Torok M, et al. (2023) Relationship between Some Myostatin Variants and Meat Production Related Calving, Weaning and Muscularity Traits in Charolais Cattle. Animals 13(12): 1895.
-
Grobet LJR, Martin D, Poncelet D, Pirottin B, Brouwers J, et al. (1997) A Deletion in the Bovine Myostatin Gene Causes the Double-Muscled Phenotype in Cattle. Nature Genetics 17: 71-74.
-
Charlier C, Coppieters W, Farnir F, Grobet F, Leroy PL, et al. (1995) The mh Gene Causing Double-Muscling in Cattle Maps to Bovine Chromosome 2. Mamm Genome 6(11): 788-792.
-
Weber T, Rorato PRN, Lopes JS, Comin JG, Dornelles MA, et al. (2009) Genetic Parameters and Genetic and Phenotypic Trends for Productive and Conformation Characteristics in the Pre-Weaning Phase in an Aberdeen Angus Breed Population. Brazilian Journal of Animal Science 38(5): 832-842.
-
Rural C (2021) Steers achieve carcass yield above 65%. São Paulo-Fernandópolis.
-
Fiems LO (2012) Double Muscling in Cattle: Genes, Husbandry, Carcasses and Meat. Animals 2(3): 472-506.
-
Albera A, Carnier P, Groen AF (2004) Definition of a Breeding Goal for the Piemontese Breed: Economic and Biological Values and Their Sensitivity to Production Circumstances. Livestock Production Science 89(1): 66- 77.
-
Mancini G, Gargani M, Chillemi G, Nicolazzi EL, Marsan PA, et al. (2014) Signatures of Selection In Five Italian Cattle Breeds Detected by a 54K SNP Panel. Mol Biol Rep 41(2): 957-965.
-
Sarti FM, Lasagna E, Ceccobelli S, Lorenzo PD, Filippini F, et al. (2014) Influence of Single Nucleotide Polymorphisms in the Myostatin and Myogenic Factor 5 Muscle Growth- Related Genes on rhe Performance Traits of Marchigiana Beef Cattle. Journal of Animal Science 92(9): 3804-3810.
-
Sorbolini S, Bongiorni S, Cellesi M, Gaspa G, Dimauro C, et al. (2017) Genome Wide Association Study on Beef Production Traits in Marchigiana Cattle Breed. Journal of Animal Breeding and Genetics 134(1): 43-48.
-
Cappucio I, Marchitelli C, Serracchioli A, Nardone A, Francesco F (1998) A G-T Transversion Introduces a Stop Codon at the mh Locus in Hypertrophy Marchigiana Beef Subjects. Animal Genetics 29(1): 51.
-
Rodgers BD, Garikipati DK (2008) Clinical, Agricultural, and Evolutionary Biology of Myostatin: A Comparative Review. Endocrine Reviews 29(5): 513-534.
-
McPherron AC, Lee SJ (1997) Double Muscling in Cattle Due to Mutations in the Myostatin Gene. Proc Natl Acad Sci U S A 94(23): 12457-12461.
-
Lee SJ (2004) Regulation of Muscle Mass by Myostatin. Annual Review of Cell and Developmental Biology 20: 61-86.
-
Carlson CJ, Booth FW, Gordon SE (1999) Skeletal Muscle Myostatin M-RNA Expression is Fi Ber-Type Specifi C and Increases during Hindlimb Unloading. American Physiological Society Journal 277(2 pt 2): R601-606.
-
Wehling M, Cai B, Tidball JG (2000) Modulation of Myostatin Expression During Modified Muscle Use. FASEB Journal. 14(1): 103-110.
-
Bellinge RH, Liberles DA, Iaschi SP, O’brien PA, Tay GK (2005) Myostatin and its Implications on Animal Breeding: A Review. Animal Genetics 36(1): 1-6.
-
Casas E, Keele JW, Shackelford SD, Koohmaraie M, Sonstegard TS, et al. (1998) Association of the Muscle Hypertrophy Locus with Carcass Traits in Beef Cattle. Journal of Animal Science 76(2): 468-473.
-
Dunner S, Miranda ME, Amigues Y, Canon J, Georges M, et al. (2003) Haplotype Diversity of the Myostatin Gene among Beef Cattle Breeds. Genetics Selection Evolution 35(1): 103-118.
-
Shibata M, Ohshima K, Kojima T, Muramoto T, Matsumoto K, et al. (2003) Nucleotide Sequence Ofmyostatingene and its Developmental Expression in Skeletal Muscles of Japanese Black Cattle. Animal Science Journal 74(5): 383-390.
-
Mosher DS, Quignon P, Bustamante CD, Sutter NB, Mellersh CS, et al. (2007) A Mutation in the Myostatin Gene Increases Muscle Mass and Enhances Racing Performance in Heterozygote Dogs. PLoS Genetics 3(5): e79.
-
Kijas JW, McCulloch R, Edwards JE, Oddy VH, Lee SH, et al. (2007) Evidence for Multiple Alleles Effecting Muscling and Fatness at the Ovine GDF8 Locus. BMC Genet 8: 80.
-
Stinckens A, Luyten T, Bijttebier J, Maagdenberg KV, Dieltiens D, et al. (2008) Characterization of the Complete Porcine MSTN Gene and Expression Levels in Pig Breeds Differing in Muscularity. Animal Genetics 39(6): 586-596.
-
Schuelke M, Wagner KR, Stolz LE, Hübner C, Riebel T, et al. (2004) Myostatin Mutation Associated with Gross Muscle Hypertrophy in a Child. New England Journal of Medicine 350(26): 2682-2688.
-
Jakaria J, Aliyya WLN, Ismail R, Siswanti SY, Ulum MF, et al. (2021) Discovery of SNPs and Indel 11-Bp of the Myostatin Gene and its Association with the Double- Muscled Phenotype in Belgian Blue Crossbred Cattle. Gene 784: 145598.
-
Lee J, Kim JM, Garrick DJ (2019) Increasing the Accuracy of Genomic Prediction in Pure-Bred Limousin Beef Cattle by Including Cross-Bred Limousin Data and Accounting for an F94L Variant in MSTN. Animal Genet 50(6): 621- 633.
-
Meyermans R, Janssens S, Cousse A, Gorssen W, Hubin X, et al. (2022) Myostatin Mutation causing Double Muscling could Affect Increased Psoroptic Mange Sensitivity in Dual Purpose Belgian Blue Cattle. Animal 16(3): 100460.
-
Vinet A, Bouyer C, Forestier L, Oulmouden A, Blanquet V, et al. (2021) The Blonde d’Aquitaine T3811>G3811 Mutation in the Myostatin Gene: Association with Growth, Carcass, and Muscle Phenotypes in Veal Calves. J Anim Sci 99(2): skab039.
-
Kobolak J, Gócza E (2002) The Role of the Myostatin Protein in Meat Quality-A Review. Archives Animal Breeding 45(2): 159-170.
-
Ceccobelli S, Perini F, Trombetta MF, Tavoletti S, Lasagna E, et al. (2022) Effect of Myostatin Gene Mutation on Slaughtering Performance and Meat Quality in Marchigiana Bulls. Animals 12(4): 518.
-
Bennett GL, Tait RG, Shackelford SD, Wheeler TL, King DA, et al. (2019) Enhanced Estimates of Carcass and Meat Quality Effects for Polymorphisms in Myostatin and µ-Calpain Genes. Journal of Animal Science 97(2): 569-577.
-
Cushman RA, Tait RG, McNeel AK, Forbes ED, Amundson OL, et al. (2015) A polymorphism in Myostatin Influences Puberty but Not Fertility in Beef Heifers, Whereas µ-Calpain Affects First Calf Birth Weight. Journal of Animal Science 93(1): 117-126.
-
Lee SJ, McPherron AC (2001) Regulation of Myostatin Activity and Muscle Growth. Proc Natl Acad Sci U S A 98(16): 9306-9311.
- In Situ Evaluation of the Anthelmintic Effect of the Aqueous Extract of Syzygium aromaticum (L) Merr and Perry on Bovine Strongyles
- Successful in Vitro Embryo Production with Oocytes Aspirated from Live White-Tailed Deer (Odocoileus Virginianus Texanus) Donors under Captivity in Northeast Mexico
- Bangladeshi Finches with their Evolutionary Thoughts (Aves: Passeriformes)
- Note on the Survival Status of Przewalski’s Horse, Equus ferus przewalskii (Perissodactyla: Equidae)
- In-Situ and Ex-Situ Protection of White-Breasted Waterhen (Amaurornis Phoenicurus) (Pennant, 1769) (Aves: Rallidae)
- Meat Examination in the Laboratory, the Acceptability and the Human Health