Potential of Synthetic Hexapod Wheat for Stripe Rust Resistance in Ethiopia
Stripe rust caused by Puccinia striiformis f.sp. tritici is one of the major constraints of wheat production worldwide. The most recent epidemics was occurred in 2010 in major wheat growing regions of central, west Asia, north and sub-Saharan Africa causing significant yield losses because of breakdown of resistance in predominantly cultivated wheat varieties (e.g. Kubsa/Attila and Galama in Ethiopia and Cham-8 in Syria). The major cause might be the narrow genetic base on which the breeding for resistance has been founded. Many control measures have been used to minimize the losses incurred by yellow rust but use of resistant cultivars remains the most economical, efficient and environment and farmer friendly strategy. To broaden the genetic basis of wheat cultivars, it is important to collect, evaluate and document new source of resistance genes from wild relatives of wheat including Triticum and Aegilops species. Synthetic hexaploid wheat (SHW) is a valuable genetic resource for resistance to a range of biotic stresses. A total of 653 SHWs derived from Aegilops tauschii and Triticum turgidum subsp. durum were evaluated for resistance to yellow rust in Meraro and Kulumsa, Ethiopia, at the adult plant growth stage. Of these, 644 entries were further tested on wheat cultivars carrying Yr2, Yr6, Yr7, Yr9, YrA, Yr25 and Yr27 against stripe rust isolates virulent on these genes at the seedling growth stage of 116 exhibited resistant to moderately resistant reaction under field conditions in both locations. Of these, 40 and 76 SHWs showed susceptible and resistant reactions at the seedling stage, respectively. The resistant SHWs identified could be useful in broadening the genetic bases of stripe resistance and further characterized to uncover potentially new resistance gene(s) in SHWs effective against prevalent races currently attacking wheat in Ethiopia and other stripe rust countries in the region.
Introduction
Stripe (yellow) rust caused by Puccinia striiformis f.sp. tritici, is one of the major diseases of wheat in temperate regions as well as in the highlands of the tropics and subtropics. It is reported to be one of the major wheat diseases in Ethiopia, Kenya, Tanzania and Uganda, Syria and Lebanon, and Yemen [1, 2, 3, 4, 5]. This is accentuated by the re-occurring breakdown in resistance of genes conferring resistance to prevailing races of the pathogen in major wheat growing regions of the world. Stripe rust infects leaves, leaf sheath and spikes of a wheat plant, and infection of spikes at a higher altitude often leads to significant yield losses including at higher altitudes regions of Ethiopia. In Ethiopia, the yield loss due to stripe rust could be as high as 96% depending on the susceptibility of the host and environmental conditions [6]. On average, a yield loss in the most recent outbreaks in Ethiopia is estimated at 30-40%.
New races may develop through mutation or recombination of nuclei via a para-sexual process [7]. The fungus was considered to have only a hemi-form of life cycle comprising uredial and telial states, but recently Berberis sp. has been reported as an alternate host, and most likely played an active role in generating new races in Pst as well. In earlier studies, the stripe rust races identified in East African were found to be virulent on most of the known seedling resistance genes [3, 8]. The overlapping and or continues planting dates and favorable environmental conditions present in different eco-zones as well as the presence of volunteers in wheat fields provide continuous sources of inoculums within or between countries in East Africa. Further, the mutation rate of the pathogen could be high in the higher elevations probably due to intensive UV light. Previous studies indicated that the mutability of yellow rust isolates increased when exposed to UV light [9].
Due to similarities in the distribution of cereal rust races among the East African countries, a common epidemiological zone was proposed [10, 11]. Rust spores are carried by wind among the East African countries and the Middle East [12]. The stripe rust virulence for Yr9 detected in Kenya in 1986 took 10-12 years to reach Southeast Asia. Similarly, the yellow rust virulence on cv. Attila ’S’ was detected in Uganda in 1994, and four years later was detected in Ethiopia in 1998 [13]. Recently, similar race virulent on Atilla “S” has been detected in Syria, North Africa and other Middle east [14].
Wellings provided a historical review of the major stripe rust epidemics including the extent of severity and losses [15]. It continues to pose a major threat to wheat production and food security in many parts of the world. In 2010, 2014 and 2015 stripe rust outbreaks occurred in Ethiopia most of the high yielding bread wheat cultivars succumbed to stripe rust. To cope with the ever-changing races, the resistance of wheat cultivars should be broadened and diversified. The most recent cultivated wheat varieties in CWANA including Ethiopia are susceptible to stripe rust. In the absence of resistance in cultivated wheat varieties new sources are sought from related species [16].
Significant genetic diversity for a wide range of biotic stresses has been reported in synthetic hexaploid wheat and their derived wheat lines including stripe rust [17, 18, 19, 20]. The diploid ‘goat grass’, Aegilops tauschii (Coss.) Schmal. (= Aegilops squarrosa L., syn. T. tauschii Coss.) (DD, 2n=2x=14) as well as the tetraploid, Triticum turgidum (2n=4x=28, AABB) species has been reported to be valuable sources of resistance to several pests and diseases [21, 22, 23]. There are several reports on techniques used to transfer rust resistance from putative progenitors into bread wheat [16]. The production of amphiploids, so called synthetic hexaploid wheat, from durum wheat, T. durum and Ae. tauschii and their subsequent crossing with bread wheat promotes full transfer of genetic material from both parents [24, 25, 26]. Despite the potential rust resistance sources in the wild progenitors of common wheat, relatively few stripe rust resistance genes have been identified and catalogued from these [27, 28]. The yellow rust genes, Yr8, Yr15, Yr17, Yr28 and Yr36 are the only genes known to originate from wild progenitors of wheat such as Aegilops comosa, wild emmer wheat, Ae. ventricosa, Ae. tauschii W-219, Turgidum ssp. diccocoides, and Ae. variabilis [29, 30, 31, 32, 33, 34]. This implies that there exist untapped genetic resources, which could be useful to broaden the resistance of the currently grown bread wheat cultivars. It is thus essential to identify additional sources of resistance.
The objective of this study was to identify stripe rust resistance in synthetic hexaploid wheat derived from different Ae. tauschii and T. durum crosses to the prevailing races of yellow rust races in Ethiopia which could be used in breeding to improve YR resistance in elite locally adapted wheat cultivars.
Materials and methods
Seedling Tests
A total of 644 SHWs derived Ae. tauschii and Triticum durum were evaluated for seedling response in Ethiopia in 2013-14. The SHWs genotypes are listed in Table 1 (see Supplement Table 1). Seedling assessment was carried out at ICARDA according to standard procedure using a local race of Pst virulent for Yr2, Yr6, Yr7, Yr8, Yr9, YrA, Yr25, Yr27, and YrSd genes [27]. Seedlings were grown at 20-25°C in a greenhouse. The 9-day old seedlings were inoculated using fresh urediniospores suspended in Saltrol 170®, followed by incubation at 10°C and relative humidity close to 100% for 24 hours. The seedlings were then transferred to a glasshouse at 18°C. Infection types were recorded 15 and 17 days after inoculation using 0-4 scale [27]. Infection types 0; (fleck), 1, and 2 were considered as resistant (low infection types; LIT) and 3 to 4 were considered as susceptible (high infection types; HIT). In all seedling assessments the susceptible variety Morocco was used as susceptible check.
Field Adult-Plant Screening
A total of 653 SWHs (see Supplement Table 1) were evaluated against stripe rust at adult plant growth stage at two locations in Ethiopia namely, Meraro (extreme highland, ca. 2920 masl) and Kulumsa (mid altitude, ca. 2230 masl). Each entry was planted in two rows of 1- meter length and 30 cm row space. Artificial inoculation was carried out three times during seedling (2 leaf stage), tillering and booting stages using the same isolate used for seedling test. Meraro is a major hot spot site for stripe rust development and is a designated stripe rust screening site in Ethiopia. Fertilizer and other agronomic practices were applied according to the recommendation for each location.
The field responses were recorded according to Roelfs et al for main rust infection types (R, MR, MS and S) and for disease severity (0-100%) according to modified Cobb’s scale when field response of the susceptible check Morocco reached to 100S [35, 36]. Field scoring was recorded three times at early booting, flag leaf, and heading stages, final scoring on flag leaf was used in data analysis. The field severity data were converted to coefficient of Infection (CI) by multiplying with a constant value of 0.2, 0.4, 0.6, 0.8, and 1 for R, MR, MS and S type reactions, respectively according Stubbs [11]. The SHWs were clustered into different resistance groups based on the CI values of the susceptible (Kubsa and Galama), Moderately susceptible (ET-13A2 and K6295-4A), and Resistant to Moderately Resistant cultivars (Meraro and Kenya Kudu); i.e., Resistant (CIs= 0-20), Moderately Resistant, (CIs=20-30), Moderately Susceptible, (CIs= 30- 40), Moderately Susceptible to Susceptible, (ICs= 40-60), and Susceptible (CIs= 60-100).
Results
The SHWs displayed a wide range of reaction to field infections in the trial sites. Frequency of adult-plant responses of SHWs to stripe rust at Kulumsa and Meraro is shown in Figure 1.
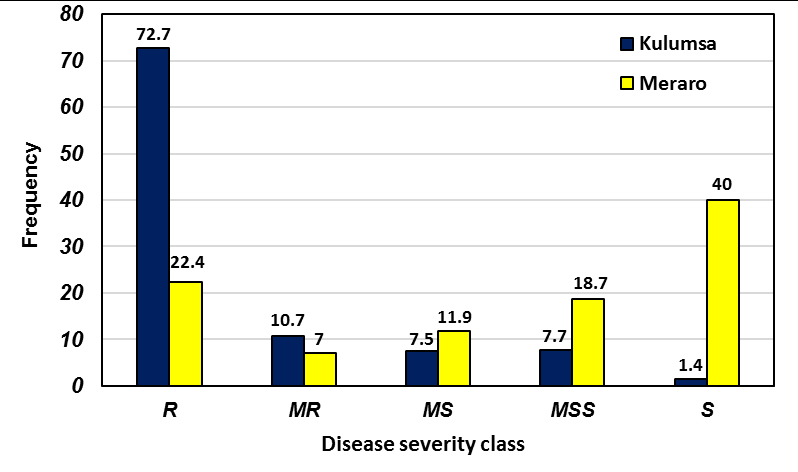
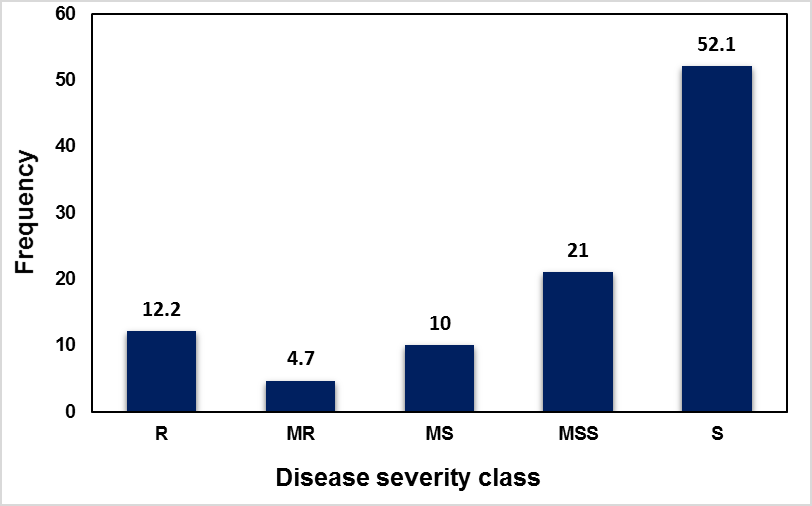
Out of 653 SHW genotypes evaluated, 72.7%, 10.7% and 7.5% showed resistant, moderately resistant and moderately susceptible reactions, respectively at Kulumsa. At Meraro, 22.4%, 7.0% and 11.9% of the SWHs exhibited resistant, moderately resistant, and moderately susceptible reaction, respectively (Fig. 1). The highest incidence of stripe rust was observed at Meraro where the some of the dominant wheat varieties, such as Galama and Kubsa showed up to 100% severity. The severity of 10MS was observed on the Kenyan old variety, Kenya Kudu and the severity of stripe rust on the two old Ethiopian wheat cultivars, K62954A and ET13-A2 was 20- 30% (data not presented). The reaction of SHWs to stripe rust was compared across locations. Out of 644 SHW genotypes common across the two locations in Ethiopia, about 12.2 %, 4.7 % and 10% exhibited resistant, moderately resistant, and moderately susceptible reaction, respectively (Figure 2).
The SHWs were compared for their reaction to stripe rust at seedling and adult plant growth stages. Out of 644 SHWs tested at both growth stages, 116 exhibited resistances to moderately resistance reactions under field conditions across locations. Of these, 40 and 76 SHWs exhibited seedling susceptible and resistance reactions, respectively. This indicating the presence of adult plant and seedling resistances in the SHWs.
Discussion
There was an outbreak of stripe rust epidemics in Ethiopia and other CWANA countries 2010, 2014, 2015 and 2016 season in almost all major wheat growing areas. Countries previously experienced yellow rust epidemics resulting in significant yield losses to farmers. In Ethiopia, stripe rust often cause substantial yield loss in higher elevation (>2400 masl), however, in 2010, 2014, and 2015 the disease was wide spread reaching even to the lower elevations as a result of virulence to Yr27 present in the most widely grown cultivar, ‘Kubsa’. Current management strategies to reduce the impact of yellow rust incidence include chemical control and use of resistance cultivars. In countries such as Ethiopia where Yr resistance breakdown is frequent, chemical control is not sustainable due to costs and the risk to environment. It is imperative to broaden the genetic base of resistance by incorporating both major and minor genes in locally adapted wheat varieties. The main objective of this study was to identify SHWs that possess resistance to stripe rust, which could then be used for crosses with adapted wheat cultivars to develop resistant varieties.
Previous studies have evaluated collections of SHW for resistance to yellow rust [17, 18, 37]. In the current study, we evaluated a relatively large number of SHWs aimed at identifying resistance to Yr27 virulent race that devastated wheat crops in Ethiopia. Screening of 644 SHW lines in Ethiopia resulted in the identification of a considerable number of lines that exhibit varying degrees of resistance to prevailing yellow rust isolates and/or races in Ethiopia virulent to Yr27, the gene present in the mega-cultivars Kubsa and Glama that occupies large wheat acreages in the country. SHWs classified as exhibiting resistant and moderately resistant phenotypes represents collectively 29% and 84% in Meraro and Kulumsa, Ethiopia respectively. Some of the SHW lines showed field resistance under field conditions at Meraro and Kulumsa, Ethiopia. This suggests that the resistance effective to field isolates in the highland and lowlands of Ethiopia, also confers resistance to many isolates in the region. Additionally, this may imply that these SHW possess multiple resistance genes that confer resistance to a broad range of isolates. Ogbonnaya, et al. reported the identification of SHW that confer multiple disease resistance including rusts [20]. This is also consistent with previous studies which reported SHWs to be a reservoir of useful genes for disease resistance in wheat [38, 39, 40]
With the frequent breakdown of resistance in wheat cultivars possessing major gene resistance to stripe rust, it has been proposed that the use and accumulation of durable rust resistance will improve the shelf-life of resistance genes and as such durable. Rust resistance genes in wheat fall under two broad categories and are referred to as seedling and adult plant resistance (APR) genes. Seedling resistance genes are detected during both the seedling and adult plant stages and as such constitute an all stage resistance phenotype. APR is commonly detected at the post-seedling stage and often as field resistance, although some APR genes can be induced to express in seedlings by varying the growth temperature and light conditions (reference for example Lr24/Yr18). A large proportion of seedling resistance genes exhibit phenotypes of major effect and with varying infection types whereas most of the APR genes are partial in effect with varying levels of disease severity [41]. Lines that possess durable rust resistance have been described as those that are susceptible at the seedling stage but resistant at the adult plant stage [42, 43]. About 40% the SHW evaluated displayed Adult plant resistance (APR) phenotype at the two sites, Meraro and Kulumsa in Ethiopia. These constitute valuable genetic resources that could be used in wheat breeding programs.
Of the more than 70 yellow rust resistance genes/QTL designated to date, only a few of them are classified as adult-plant resistance to stripe rust of which very limited number such as Lr34/Yr18 and Lr46/Yr29 are well characterized and cloned. This raises some questions, do the SHWs with APR phenotypes possess the same gene as those currently available in elite cultivars. Therefore, further studies would be needed to determine if the potentially new APR genes identified in this study are the same genes to those previously identified. The availability of molecular markers linked to some of the designated APR genes would facilitate the screening of the SHWs to determine if these are different. If so, the SHW provide potentially new sources of APR genes that could be used for crosses in developing locally adapted cultivars resistant to prevailing Yr isolates/or races virulent to Yr27.
Acknowledgments
We gratefully acknowledge the financial support from Ethiopian Institute of Agricultural Research (EIAR) and the technical support from Kulumsa wheat pathology team.
References
-
Badebo A, Stubbs RW (1995) Valuable sources of resistance of wheat to the East African yellow rust isolates. Breeding for disease resistance with emphasis on durability. Proceedings of a regional workshop for Eastern, Central, and Southern Africa. D L Danial (eds) pp: 206-214.
-
Danial DL, Stubbs RW (1992) Virulence of yellow rust races and types of resistance in wheat cultivars in Kenya. In: Seventh Regional Wheat Workshop for Eastern, Central, and Southern Africa. Nakuru, Kenya: CIMMYT, Tanner DG, Mwangi W (eds) pp: 165-175.
-
Louwers JM, van Silfhout CH, Stubbs RW (1992) Race analysis of yellow rust in wheat in developing countries report 1990-1992. Wageningen, The Netherlands pp: 23.
-
Hakim MS, Mamluk OF (1996) Virulences of wheat yellow rust pathogen in Syria and Lebanon. Proceedings of the 9th European and Mediterranean Cereal Rust and Powdery Mildew Conference. 2-6, September 1996. Lunteren, The Netherlands pp: 141.
-
Bahmish HS (1996) Preliminary results on wheat rusts in Yemen. Paper presented during the 16th NVRP meeting. ICARDA. Sanaa, Yemen.
-
Bekele E (1986) A review of research on diseases of barley, tef, and wheat in Ethiopia. A review of crop protection research in Ethiopia. Proceedings of the first Ethiopian Crop Protection Symposium. 4-7 February, 1986. Addis Ababa, Ethiopia, Abate T (eds), pp: 79-108.
-
Little R, Manners JG (1969) Somatic recombination in yellow rust of wheat (_Puccinia striiformis_). I. The production and possible origin of two new physiologic races. Trans of the British Mycol Soc 53(2): 251-258.
-
Jin Y (2011) Role of Berberis spp. as alternate hosts in generating new races of _Puccini graminis_ and _P._ _striiformis_. Euphytica 179(1): 105-108.
-
Johnson R, Priestley RH, Taylor EC (1978) Occurrence of virulence in _Puccinia striiformis_ for Compair wheat in England. Cereal Rusts Bull 6: 11-13.
-
Saari EE, Prescott JM (1985) World distribution in relation to economic losses. Diseases, distribution, epidemiology, and control pp: 259-298.
-
Stubbs RW (1988) Pathogenicity analysis of yellow (stripe) rust of wheat and its significance in a global context. Breeding strategies for resistance to the rusts of wheat pp: 23-38.
-
Dubin HJ, Rajaram S (1996) Breeding disease-resistant wheat for tropical highlands and lowlands. Ann Rev Phytopathol 34: 503-526.
-
Badebo A, Fehrmann H, Yahyaoui A (2008) Status of wheat yellow rust (_Puccina_ _striiformis_) races and their virulence in major wheat growing areas of Ethiopia. Pest Mgt J Eth 12: 1-7.
-
Nazari K (2011) Yellow rust in CWANA in 2010 and 2011: The situation and measures to control it. In: BGRI 2011 technical workshop. St. Paul, Minnesota, USA.
-
Wellings C (2010) Global status of stripe rust. BGRI 2010 Technical Workshop, St Petersburg, Russia.
-
Knott DR (1989) The wheat rusts: Breeding for resistance. In: Monograph on theoretical and applied genetics 12. Frankel R,. Grossman M, Linskens HF, Maliga P, Riley R (eds.). Springer-Verlag, Berlin, pp: 201.
-
Ma H, Singh RP, Mujeeb Kazi A (1995) Resistance to stripe rust in _Triticum turgidum_, _T_. _tauschii_ and their synthetic hexaploid. Euphytica 82(2): 117-124.
-
Yang WY, Lu BR, Yu Y, Hu XR (2001) Genetic evaluation of synthetic hexaploid wheat for resistance to the physiological strain CYR30 and CYR31 of wheat stripe rust in China. Jour Genet Mol Genet 12: 190- 198.
-
Badebo A, Fehrmann H (2005) Resistance to yellow rust in _Aegilops tauschii (_Coss.), _Triticum_ _durum_ and their synthetic amphiploids_._ Ethiopian Jour of Agric Sci (EJAS) 18(2): 129-135.
-
Ogbonnaya FC, Imtiaz M, Bariana HS_,_ McLean M, Shankar M, et al. (2008) Mining synthetic hexaploids for multiple disease resistance to improve bread wheat. Aust Jour of Agric Res 59(5): 421-443.
-
Kimber G, ZhaoYH (1983) The D genome of _Triticeae_. Can J Genet Cytol 25(6): 581-589.
-
Cox TS, Raupp WJ, Wilson DL, Gill BS, Leath S, et al. (1992) Resistance to foliar diseases in collection of _Triticum tauschii_ germ plasm. Plant Disease 76: 1061- 1064.
-
Halloran GM, Ogbonnaya FC, Lagudah ES (2008) _Triticum_ (_Aegilops_) _tauschii_ in the natural and artificial synthesis of hexaploid wheat. Aust Jour Agric Res 59(5): 475-490.
-
Gill BS, Raupp WJ (1986) Direct genetic transfers from _Aegilops squarrosa_ L. to hexaploid wheat. Crop Sci 27(3): 445-450.
-
Lange W, Jochemsen G (1992) Use of the gene pools of _Triticum turgidum ssp. dicoccoides_ and _Aegilops_ _squarrosa_ for breeding of common wheat (_Triticum_ _aestivum_) through chromosome-doubled hybrids. Euphytica 59(2-3): 213-220.
-
Mujeeb-Kazi A (1993) Interspecific and intergeneric hybridization in the _Triticeae_ for wheat improvement. Biodiversity and wheat improvement pp: 95-102.
-
McIntosh RA, Wellings CR, and Park RF (1995) Wheat Rusts: An Atlas of Resistance Genes. Kluwer Academic Publishers, London.
-
McIntosh RA, Yamazaki Y, Dubcovsky J, Rogers J, Morris C, et al. (2008) Catalogue of Gene Symbols for Wheat- morphological and physiological traits. 11th International Wheat Genetics Symposium 24-29 August, Brisbane Qld Australia.
-
Riley R, Chapman V, Johnson R (1968) Introduction of yellow rust resistance of _Aegilops comosa_ into wheat by genetically induced homoeologous recombination. Nature 217: 383-384.
-
Gerechter-Amitai ZK, van Silfhout CH, Adriana G, Kleitman F (1989) _Yr_15-a new gene for resistance to _Puccinia striiformis_ in _Triticum dicoccoides_ Sel.G-25. Euphytica 43(1-2): 187-190.
-
Bariana HS, McIntosh RA (1994) Characterization and origin of rust and powdery mildew resistance genes in VPM1 wheat. Euphytica 76(1-2): 53-61.
-
Singh RP, Nelson JC, Sorrells ME (2000) Mapping _Yr_28 and other genes for resistance to stripe rust in wheat. Crop Sci 40(4): 1148-1155.
-
Uauy C, Brevis JC, Chen X, Khan I, Jackson L, et al. (2005) High-temperature adult-plant (HTAP) stripe rust resistance gene _Yr36_ from _Triticum turgidum_ ssp. _dicoccoides_ is closely linked to the grain protein content locus Gpc-B1. Theor Applied Genet 112(1): 97-105..
-
Dhaliwal HS, Singh H, Gill KS, Randhawa HS (1993) Evaluation and cataloguing of wheat germplasm for resistance and quality. A Wiley-Sayce Publications, The Netherlands pp: 123-140.
-
Roelfs AP, Singh RP, Saari EE (1992) Rust diseases of wheat: Concepts and methods of disease management. CIMMYT pp: 81.
-
Peterson RF, Campell AB, Hannah AE (1948) A diagrammatic scale of estimating rust intensity on leaves and stems of cereals. Can Jour Rec 26(5): 496- 500.
-
Ogbonnaya FC (2011) Development, management and utilization of synthetic hexaploids in wheat improvement. The World Wheat Book-A history of Wheat Breeding 2: 823-843.
-
Siedler H, Obst A, Hsam SLK, Zeller FJ (1994) Evaluation for resistance to _Pyrenophora tritici-_ _repentis_ in _Aegilops tauschii_ Coss and synthetic hexaploid wheat amphiploids. Genet Resour Crop Evol 41(1): 27-34.
-
Mujeeb-Kazi A, Delgado R (2001) A second, elite set of synthetic hexaploid wheats based upon multiple disease resistance. Ann Wheat Newsl 47: 114-116.
-
Xu SS, Friesen TL, Mujeeb-Kazi A (2004) Seedling resistance to tan spot and _Stagonospora_ leaf blotch in synthetic hexaploid wheats. Crop Sci 44: 2238-2245.
-
Lagudah ES (2011) Molecular genetics of race non- specific rust resistance in wheat. Euphytica 179(1): 81-91.
-
Singh RP, Hodson DP, Huerta Espino J, Jin Y, Njau P, et al. (2008) Will stem rust destroy the world’s wheat crop?. Adv Agron 98: 271-309.
-
Badebo A**,** Assefa S, Fehrmann H (2008) Yellow rust resistance in advanced lines and commercial cultivars of bread wheat from Ethiopia. East African Jour of Sci 2(1): 29-34.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note