Enhancement of Tolerance to Sheath Blight in Indica Rice through Incorporation of Chitinase Genes
Rice sheath blight, caused by Rhizoctonia solani, a polyphagous fungus, is a major disease on rice and causes up to 50% yield losses. Genetic enhancement of host plant disease resistance is an attractive solution to address diseases like sheath blight. The major problem associated with enhancement of host (rice) resistance to sheath blight through conventional approaches is the lack of resistant donors in the rice gene pool and only moderate levels of resistance is available. In this study, an attempt was made to enhance host plant resistance through incorporation of a chitinase gene employing biolistic approach. The incorporation of the transgene was confirmed through molecular analyses like PCR and Southern assays. The results of the bioassay studies against the pathogen Rhizoctonia solani did indicate higher levels of resistance moreover the inheritance pattern of the transgene was Mendelian. The transgenics generated did show high promise and can provide a practical solution to tackle sheath blight, a major disease on rice.
Introduction
Sheath blight disease, caused by fungus Rhizoctonia solani, is the most widely spread disease in all the rice growing countries [1]. This disease has been a constant major biotic threat, lowers the productivity of superior rice cultivars by causing yield losses in the range of 5.2 - 50 % [2] and pathogen populations change to overcome the incorporated resistance [3]. Despite many years of continuous and sustained efforts by conventional breeders, consistent progress has not been made in the development of suitable cultivars to effectively combat the rice sheath blight due to lack of truly resistant donors in the rice gene pool [4] although genes conferring only partial resistance have recently been identified [5]. With shrinking land resources and ever increasing demands of food production, the productivity levels has to be raised substantially to increase the overall rice production in the country. Through genetic improvement especially that geared towards the development of varieties resistant/highly tolerant to sheath blight, the production levels of rice will increase through stabilization of yields.
Genetic engineering techniques can augment the conventional approaches through incorporation of novel genes from different sources into rice thereby widening the genetic base of rice. A major goal of genetic engineering is the development of improved natural defense capabilities of the host plants in cases like sheath blight where use of chemical fungicides in controlling this disease has been ineffective, inefficient, expensive and non ecofriendly. The present work is an effort in this direction and the studies were focused on development of transgenic rice employing biolistic approach through over expression of a Chitinase-11 gene in Pusa Basamti1, a popular aromatic indica rice cultivar.
Materials and Methods
Plasmid Construct
The transformation vector pNO1, harboring a hygromycin (hpt) selectable marker gene and Chitinase- 11 (gene of interest) under control of cauliflower mosaic virus (CaMV) 35S promoter was employed.
Transformation and Development of Putative Transgenics
Scutellum derived embryogenic calli obtained from the sterilized kernels of Oryza sativa L. indica variety cv. Pusa Basmati1 (PB1) were bombarded at 1100 psi helium pressure with the plasmid vector pNO1 coated on gold micro carriers using the particle gun PDC-1000/He system (BIORAD) following manufacturer’s instructions. Plasmid extraction, explant preparation for bombardment, transformation and selection were done as described earlier [6].
Actively dividing calli were transferred from selection medium to regeneration medium [7]. (i.e. MS medium supplemented with BAP (1.5 mg l-1) + Kn (0.5 mg l-1) + NAA (0.5mgl-1). The cultures were incubated under cool fluorescent light (2000 lux) for one month. The regenerated green plants were transferred to MS semisolid rooting medium supplemented with NAA (1.0 mg l-1), Kinetin (0.1 mg l-1), maltose (50 gm l-1) and agar (8g l-1). The well-established plants were transferred from the rooting medium to pots and were grown in the transgenic green house under controlled conditions (temperature -27-30°C; light intensity - ~Ca 2000 lux) and relative humidity (55-60%).
PCR Analysis
DNA was isolated from the fresh leaves [8]. Isolated DNA was examined for the presence of transgene with the help of specific primers and amplified by polymerase chain reaction (PCR) as follows; 1µl of plant DNA(20ng) along with 0.8µl of 2.5mMdNTPs (Fermentas), 1.0 µl of 10X PCR buffer (Sigma), 0.2µl of Taq DNA Polymerase(5U/µl Sigma),1 µl each of both forward & reverse primers (5 pico moles/µl Sigma), 5 µl of autoclaved sterile distilled water were added (total volume of 10 µl reaction) and amplified in a thermal cycler (Eppendorf Vapo protect) with the following experimental conditions: initial denaturation at 94ºC for 3min final denaturation at 94ºC for 1min, Annealing at 60ºC for 1min, extension at 72ºC for 1min and final extension at 72ºC for 10min. Using specific primers, amplification of the specific portion of the chitinase-11 gene was attempted through PCR using the DNA isolated from the putative transgenic plants as the template. The PCR products, after separation on a 1.5% agarose gel, were analysed on a gel documentation system (Alpha innotech).
Southern Hybridization
Ten micrograms of genomic DNA was digested with PstI restriction endonuclease enzyme PstI overnight at 37°C in order to release the pGL2 (CaMV-Chi11–Ubi terminator) cassette (~ 1.1 kb). The digested samples were separated by electrophoresis in a 0.8% agarose in 1X TBE buffer [9]. The DNA was transferred by capillary blotting to a nylon membrane (Micro Separations Inc., Westboro, MA) and was cross linked by UV light (Bioslink, Bios Corporation, CT). The 1.1 kb chitinase-11 DNA isolated from pCGN1589 was used as probe, and labeled with 32P-dCTP (NEN, Du Pont) by random primer labeling following the manufacturer’s instructions (Life Techonologies, Gaithersburg, MD). The hybridizations were performed according to the manufacturer’s specifications (Micro Separations Inc., Westboro, MA). The blot was washed in 5X SSC, 0.5% SDS for 30 minutes at room temperature; in 1X SSC, 0.5% SDS for 30 minutes at 65°C; followed by a final wash in 0.1X SSC, 1% SDS at 65°C. The blot was kept in radio cassette for overnight and visualized in phosphoimager (GE Health care).
Bioassay Against Rhizoctonia Solani
Bioassays were conducted with the homozygous transgenic rice lines of the T2 generation to assess their level of resistance to sheath blight using RS7, a highly virulant isolate. The assays were conducted on ~75 day aged plants in pots placed in transgenic greenhouse. Agar plugs (5-mm thick, 9.5-mm diameter) with the R. solani isolate were placed between the culm and the sheath of a tiller. Three healthy tillers of each plant were inoculated and wrapped with wet cotton and parafilm individually
and nine plants of each transgenic line constitute a
replication. Cotton was wetted with water twice a day.
The score based on the sheath blight symptoms was
recorded at 7 and 14 days intervals after inoculation [10].
The spread of symptom to a distance covering 15 cm
above and 15 cm below the site of inoculation (total
distance 30 cm) was measured, and grade points of 0, 1, 2,
3, 4 and 5 were assigned, i.e., for no symptom (0),
symptom development up to 6-cm (1), 12-cm (2), 18-cm
(3), 24-cm (4), and above 30 cm (5), respectively [11].
After assigning the grade points for each plant (mean of
three tillers), disease index of each plant was determined
using the formula:
$$ D i s e s e \quad i n d e x = \mathrm {M e a n} \quad g r a d e \quad p o i n t \quad p e r \quad p l a n t $$
x100/Maximum grade point (5)
Results
After transformation and selection over four cycles, fifteen plantlets were generated. Through PCR analysis, 4 positive plants were identified among the 15 putative transgenics (Figure 1). In the four positive plants, an amplified product of the expected size of chitinase-11 gene, was similar in size to that of the product obtained with the positive control (plasmid). PCR assays conducted on these four positive plants also revealed the presence of hpt gene in these plants as evidenced by the amplification product of a 0.95kb region of hpt gene (Figure 2).
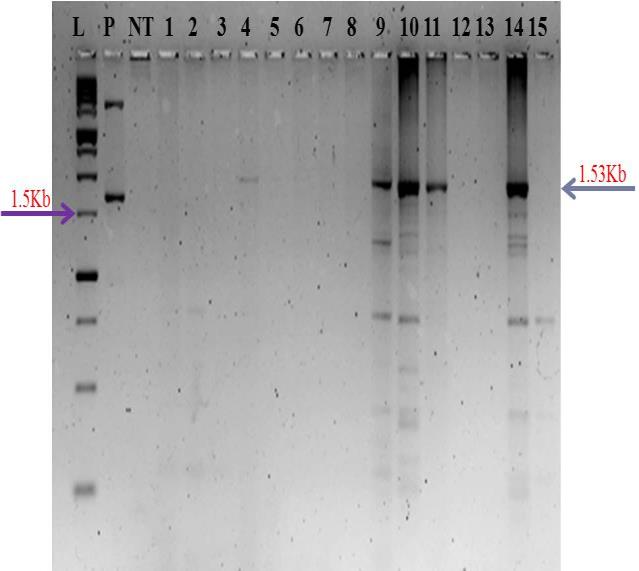
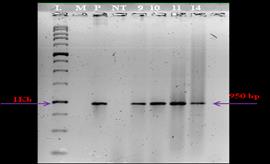
L-1Kb ladder, M-PCR control, P-plasmid, NT-Wild type, Nos. 9,10,11,14 –samples.
When the genomic DNA isolated from the PCR positive putative transgenic plants was digested with Hind III restriction enzyme, transferred on to a membrane and hybridized with the 1.53-kb Hind III fragment of pGL2CaMV35SCHI11, carrying the chitinase-11 gene coding sequence along with the promoter, the signal was observed at the expected 1.1-kb level in all the four transgenics that were identified by PCR analysis while the signal was not observed in the non transformed (wild type) plants (Figure 3).
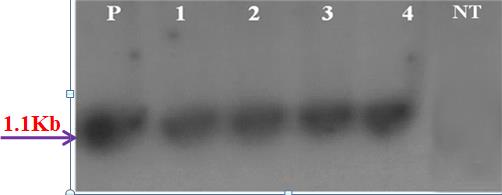
P-plasmid, Nos. 1,2,3,4 - Plant samples.
NT-Non Transformant (wild type); Nos. 1-4 corresponds to samples 9,10,11,14 respectively of Figure 1.
- Inheritance of the Transgene
- The fertility status of the T0 generation transgenics of
- Pusa Basmati1 was normal and showed full fertility. The first generation transgenics were grown to maturity in the transgenic green house and T1 generation was raised. A total of 27 plants were raised from PC 9 and PC10, two analyzed
- No. of Progenies chi 11+ No. of Progenies chi11–
- 2 fitness for
- 3:1 ratio (df 1)
- T1 Progeny line
- No. of T1 progenies
- PC 9
- 12
- 11
- 1
- 1.77ns
- PC 10
- 15
- 13
- 2
- 1.36ns
Table 1: Segregation of chitinase transgene in the T1 generation.
Bioassays
Bioassay of the T2 generation lines derived from the positive plants (PC9 and PC10) was conducted against a virulent isolate of sheath blight under the transgenic green house conditions using established protocols. The results based on the relative integrated infection index showed enhanced resistance in the chitinase 11 containing transgenic lines as compared to the controls (Figure 4). The PC9 progeny lines showed higher levels of resistance by exhibiting low levels of disease spread. Disease index values from five individual plants recorded were: 20.00, 20.00, 13.34, 20.00 and 13.34. In the progeny lines derived from PC10, the values from five individual plants were 26.67, 20.00, 26.67, 20.00 and 26.67 and the values of the non transformed control plants was 46.00 while the disease index value of the rice cultivar Jogen, the susceptible control for sheath blight, was 52.00 (Table 2 & Figure 4).
| Scoring | |||||||||||||||||
| Plant No. | Mean Grade point | Disease index scores | |||||||||||||||
| Tiller1 | Tiller2 | Tiller3 | |||||||||||||||
| PC9-1 | 1 | 1 | 1 | 1.0 | 20.0 | ||||||||||||
| PC9-2 | 1 | 1 | 1 | 1.0 | 20.0 | ||||||||||||
| PC9-3 | 1 | 1 | 0 | 0.6 | 13.3 | ||||||||||||
| PC9-4 | 1 | 1 | 1 | 1.0 | 20.0 | ||||||||||||
| PC9-5 | 0 | 1 | 1 | 0.6 | 13.3 | ||||||||||||
| PC10-1 | 2 | 1 | 1 | 1.3 | 26.6 | ||||||||||||
| PC10-2 | 1 | 1 | 1 | 1.0 | 20.0 | ||||||||||||
| PC10-3 | 1 | 2 | 1 | 1.3 | 26.6 | ||||||||||||
| PC10-4 | 1 | 1 | 1 | 1.0 | 20.0 | ||||||||||||
| PC10-5 | 1 | 1 | 2 | 1.3 | 26.6 | ||||||||||||
| NT | 2 | 2 | 3 | 2.3 | 46.0 | ||||||||||||
| Jogen | 2 | 2 | 4 | 2.6 | 52.0 |
Table 2: Disease index values of the transgenic plants against sheath blight disease.

Discussion
Particle bombardment approach of gene transfer has several applications and has unique advantages when compared with other methods of gene transfer. Biolistic system has become the method of choice because it alleviates the need for preparing protoplasts, reduces the time needed to regenerate transgenic plants and results in transgenic plants with higher fertility [12]. Employing this approach, pure, linear DNA sequences can be transferred without any vector backbone interference. To achieve higher levels of transformation efficiency, biolistics method is preferable as large number of cells can be treated and transformed at a single time [13]. In addition, particle-mediated gene delivery is the only method reported so far to introduce foreign genes into cell organelles such as chloroplasts [14]. Transgenic plants generated by this method have been reported for Monocotyledonous species such as Kentucky bluegrass [15], jute [16], rice [17, 18], and dicotyledonous species such as mothbean [19], cowpea [20], sunflower [21], soybean [22], and wheat [23].
The in vitro antifungal potential of chitinases have been reported [24, 25]. Chitinases play an important role in host plant defense as pathogenesis-related proteins expressed in response to pathogen attack and their constitutive expression is well documented. However, the inducible defense mechanisms of plants are often too week, appear too late to be effective for protection [26]. In order to incorporate resistance against sheath blight disease in Pusa Basmati1, an elite indica rice cultivar, in the present study, an attempt was made to constitutively overexpress a chitinase gene. The PCR and Southern assays confirm the presence and integration of the introduced chitinase gene in four plants out of the fifteen plants studied so far. With the analysis of more positive putative transgenic lines, different independent events are expected to be identified and plants with different events are possible.
The results suggest that the transgenic lines displayed enhanced levels of resistance to sheath blight in comparison to the parent (non transformant) and susceptible varieties and disease levels recorded on the transgenics is much low. Though a total protection against Rhizoctonia solani was not observed in the study, the enhanced levels of resistance will be of great use in containing the disease caused by this polyphagus fungus for which no known donor is available in both primary and secondary gene pool of rice. Thus it is evident that the major objective of the study i.e., development of transgenics with enhanced resistance to sheath blight was achieved and with characterization and bioassay of more putative transgenic plants, it is expected to have lines with different integration patterns and the events may show even higher levels of resistance against this deadly disease on rice.
Acknowledgements
The authors are thankful to Director, NRRI for the facilities and encouragement. The authors Sai Krishna Repalli and Chaitanya Kumar Geda are thankful to ICAR (NPTC) for providing them Fellowship. Compliance with Ethical Standards: This manuscript is in compliance with Ethical Standards
References
-
Dasgupta M (1991) Rice sheath blight: problems and challenges In: Integrated control of diseases with minimum use of agrochemicals to obtain maximum economic yield of rice. ICAR final technical report, Visva Bharati, Palli Siksha Bavana, India.
-
Mathur SC (1983) Fungal diseases of rice in India. In: Hussain A, Singh K, Singh BP, Agnihotri VP (Eds.), Recent advances in plant pathology. Print House, Lucknow, pp: 368.
-
Peever TL, Zeigler RS, Dorrance AE, Correa-Victori FJ, St. Martin S, et al. (2000) Pathogen Population Genetics and Breeding for Disease Resistance.
-
Bonman JM, Khush GS, Nelson RJ (1992) Breeding Rice for Resistance to Pests. Annu Rev Phytopathol 30: 507-528.
-
Pan XB, Rush MC, Sha XY, Xie SD, Linscombe SR, et al. (1999) Major gene, non-allelic sheath blight resistance from the rice cultivars Jasmine 85 and Tequing. Crop Sci 39: 338-346.
-
Saikrishna R, Chaitanya KG, Rao GJN (2013) Efficacy of different transformation methods in rice (Oryza sativa L.). JEBAS 1: 106-110.
-
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15: 473-497.
-
Tai TH, Tanksley SD (1990) A rapid and inexpensive method for isolation of total DNA from dehydrated plant tissue. Plant Mol Biol Rep 8(4): 297-303.
-
Southern EM (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98(3): 503-517.
-
Sridevi G, Sabapathi N, Meena P, Nandakumar R, Samiyappan R, et al. (2003) Transgenic indica rice variety Pusa Basmati1 constitutively expressing a rice chitinase gene exhibits enhanced resistance to Rhizoctonia solani. Jr Plant Biochem Biotechnol 12(2): 93-101.
-
Sriram S, Raguchander T, Vidhyasekaran P, Muthukrishnan S, Samiyappan R (1997) Genetic relatedness with special reference to virulence among the isolates of Rhizoctonia solani causing sheath blight in rice. Jr Plant Dis Protect 104(3): 260-271.
-
Rakszegi M, Tamas C, Szucs P, Tamas T, Bedo Z, et al. (2001) Current status of wheat transformation. Plant Biotech J 3: 67-81.
-
Dalton SJ, Bettany AJE, Timms E, Morris P (1999) Co- transformed, diploid Lolium perenne (perennial ryegrass) Lolium multiflorum (Italian ryegrass) and Lolium temulentum (darnel) plants produced by microprojectile bombardment. Plant Cell Rep 18(9): 721-726.
-
Grevich JJ, Daniell H (2005) Chloroplast genetic engineering: recent advances and future perspectives. Crit Rev Plant Sci 24(2): 83-107.
-
Gao C, Jiang L, Folling M, Han L, Nielsen KK, et al. (2005) Generation of large numbers of transgenic Kentucky bluegrass (Pao pratensis L.) plants following biolistic gene transfer. Plant Cell Rep 25(1): 19-25.
-
Ghosh M, Saha T, Nayak P, Sen SK (2002) Genetic transformation by particle bombardment of cultivar jute Corchorus capsularis L. Plant Cell Rep 20(10): 936-942.
-
Li L, Qu R, Kochko A, Fauquet C, Beachy RN, et al. (1993) An improved rice transformation system using the biolistic method. Plant Cell Rep 12(5): 250- 255.
-
Jain RK, Jain S, Wang B, Wu R (1996) Optimization of biolistic method for transient gene expression and production of agronomically useful transgenic Basmati rice plants. Plant Cell Rep 15(12): 963-968.
-
Kamble S, Misra HS, Mahajan SK, Eapen S (2003) A protocol for efficient biolistic transformation of mothbean Vigna aconitifolia L. Jacq. Marechal. Jr Plant Mol Biol 21(4): 457a-457j.
-
Ikea J, Inglebrecht I, Uwaifo A, Thottappily G (2003) Stable gene transformation in cowpea (Vigna unguiculata L.walp) using particle gun method. Afr Jr Biotechnol 2(8): 211-218.
-
Molinier J, Thomas C, Brignou M, Hahne G (2002) Transient expression of ipt gene enhances regeneration and transformation rates of sunflower shoot apices (Helianthus annuus L.) Plant Cell Rep 21(3): 251-256.
-
Moore PJ, Moore AJ, Collins GB (1994) Genotypic and developmental regulation of transient expression of a reporter gene in soybean zygotic cotyledons. Plant Cell Rep 13(10): 556-560.
-
Zhang L, Rybczynski JJ, Langenberg WG, Mitra A, French R, et al. (2000) An efficient wheat transformation procedure: transformed calli with long-term morphogenic potential for plant regeneration. Plant Cell Rep 19(3): 241-250
-
Ji C, Kuc J (1996) Antifungal activity of cucumber β- 1,3-glucanase and chitinase. Physiol Mol Plant Path 49(4): 257-265.
-
Collinge DB, Kragh KM, Mikkelsen JD, Nielsen KK, Rasmussen U, et al. (1993) Plant chitinases. Plant J 3(1): 31-40.
-
Chen L, Marmey P, Taylor NJ, Brizard JP, Espinoza C (1998) Expression and inheritance of multiple transgenes in rice plants. Nat Biotechnol 16(11): 1060-1064.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note