Effects of High Temperature on the Physiological and Antioxidation Abilities of Cucumber Seedlings
When cucumber plants (Cucumis sativus L.) are subjected to high-temperature environments (>35°C), their physiological structures are disrupted, affecting male–female flower inflorescence and fruit shapes, which are critical to cucumber quality. Accordingly, selecting heat-resistant cultivars is imperative in cucumber cultivation. This study tested three cucumber cultivars with varying resistance levels to high temperature: Wen nong 210, CU-127, and Ks No. 3. When the seedlings of these cultivars grew their primary leaves, leaf discs were collected and treated at varying temperatures; subsequently, relative injury (RI) was calculated to determine the cultivars’ cell membrane stability at high temperatures. The results revealed that Wen nong 210 sustained 50% RI at 54.1°C, exhibiting the greatest cell membrane stability, followed by CU-127, which sustained 50% RI at 53.9°C, and then Ks No. 3, which sustained 50% RI at 52.8°C. Analyzing the cultivars’ physiological indices indicated that, at 45°C, Ks No. 3 had the highest alondialdehyde (MDA) content but the lowest chlorophyll content, whereas Wen nong 210 had low MDA content but high chlorophyll content. Antioxidation analysis showed that at 45°C, Ks No. 3 and Wen nong 210 exhibited the lowest and highest ascorbate peroxidase (APX) activity, respectively. Furthermore, at 35°C, Ks No. 3 exhibited high proline content. In summary, physiological indices (H2O2, MDA, and chlorophyll content), proline content, and APX activity can constitute the basis of heat resistance screening standards for cucumber plants grown under high-temperature conditions.
Introduction
High temperatures can alter crop’s structure, physiological mechanisms, and biochemical reactions, hence affecting crops’ yield and quality. Therefore, a high environmental temperature is a major limiting factor to the growth of crops. The term “high temperature stress” refers to a situation in which the observed temperature exceeds a specified threshold; when a temperature of environment exceeds an optimal growth temperature of plant by 10°C–15°C, the plant’s growth and development are irreversibly impaired [1]. Cucumber (Cucumis sativus L.) is an annual creeping plant belonging to the Cucurbitaceae family and Cucumis genus, and its favorable growth temperature is in the range 20°C–30°C. When the environmental temperature exceeds 35°C, the physiological functions of cucumber plants are impaired, affecting the inflorescence of male and female flowers and the shape and quality of fruits [2]. With the strengthening of global warming and climate change, cucumber plants are often subjected to high temperature stress levels in summer, rendering summer heat a factor that inhibits their production [3, 4].
When plants experience biotic or abiotic stresses such as drought, high temperature, flooding, ozone, saline, heavy metal, high-intensity light, and pest stresses, their metabolism generates numerous types of reactive oxygen species (ROS), including the superoxide radical (O2-∙), hydroxyl radical (HO∙), and hydrogen peroxide (H2O2). These ROS accumulate in plant cells and cause oxidative stress, damaging the structure and affecting the permeability of the cell membrane system. In severe situations, they can result in membrane lipid disintegration, disrupting cell membrane stability and causing plants to perish in stages [5, 6, 7, 8, 9]. Among the ROS, H2O2 is a cell metabolism product and constitutes a powerful oxidizer. Low-concentration H2O2 can serve as an intracellular signaling molecule. However, when cells contain iron or copper ions, the Haber–Weiss reaction occurs between H2O2 and O2 -‧, generating toxic hydroxyl radicals (OH‧) that can cause severe damage to cells. Therefore, high- concentration H2O2 has been recognized as being harmful to cells [10, 11]. When plants grow, their antioxidation mechanisms regulate their H2O2 content to promote growth and development [5, 6, 7].
Antioxidant enzymes, including ascorbate peroxidase (APX), catalase (CAT), glutathione reductase (GR), peroxidase (POX), and superoxide dismutase (SOD), are among the antioxidation mechanisms of plants. SOD is the first line of defense of plants against oxidation, converting toxic O 2 -‧ into H2O2, which is subsequently reduced into water by APX, GR, and CAT, thus mitigating free-radical-induced damage to plant cells [5, 6, 7].
Plants accumulate peroxides in high-temperature environments, engendering peroxidation in the lipids and membranes of numerous organelles, such as chloroplasts and mitochondrions [12, 13]. Lipid peroxidation can induce membrane system degradation, increasing membrane permeability [14]. Therefore, the content of a lipid peroxidation product, malondialdehyde (MDA), can be used as an indicator of the degree of lipid peroxidation [15, 16]. When Asian rice (Oryza sativa) encounters the heavy metal arsenic, the rice stalks shorten as the arsenic concentration increases. Additionally, the H2O2 content in rice increases with the arsenic concentration, and this is accompanied by an increase in MDA content and decrease in chlorophyll content [17].
Numerous researchers have investigated cucumber plants’ heat resistance properties from ecological, physiological, and biochemical perspectives or applied pretreatment to enhance heat resistance of cucumber plant [18, 19, 20]. These studies have suggested that during vegetative growth, the activity of antioxidant enzymes and the proline content in cucumber leaves are associated with heat resistance of cucumber plants [18, 21, 22]; moreover, stability of plant cell membranes in a high-temperature environment can be considered an indicator of heat resistance [23, 24, 25]. The present study placed cucumber cultivars with different levels of heat resistance in high-temperature environments to investigate the morphometric and physiological indices of cucumber seedlings. This study analyzed changes in physiological characteristics and antioxidant enzymes to establish a seedling screening model for the selection of heat-resistant cultivars.
Materials and Methods
Material and High-Temperature Treatment
Wen nong 210, CU-127, and Ks No.3 served as the cucumber cultivars used for the experiment in this study. Wen nong 210 and CU-127 are primarily grown in summer and autumn, whereas Ks No.3 is resistant to downy mildew and is suitable for summer and autumn [26].
Cucumber seeds were planted in plug trays (54.5 × 27.5 cm2; cell size 5.5 × 3.3 cm2) filled with unamended peat (pH 7.0) as the substrate. Seedlings growing their primary leaves were moved to growth chambers in which the temperature was set to 28°C, 35°C, 43°C, or 48°C for a 24-hour dark period and the relative humidity was 60%–70%. Each of the growth chambers received four seedlings per session, and a total of four sessions were conducted. Subsequently, the seedlings were moved to a nursery chamber to acclimatize for 2 days, after which various physiological readings and antioxidation properties (i.e., chlorophyll, H2O2, MDA, CAT, APX, and proline contents) were recorded. Cell membrane thermostability (CMT) analysis was performed in 28°C, 38°C, and 43°C environments, and each of these environments contained eight seedlings.
Analytical Methods
After being subjected to treatment at different temperatures, the cucumber seedlings were measured for their morphometric and physiological indices, physiological readings, chlorophyll content [27], H2O2 content [28], and MDA content [15]. Antioxidant enzymes were also analyzed in this study, with a modified version of the method [29] applied to analyze CAT activity. Similarly, a modified version of the method proposed by Nakano and Asada (1981) [30] was used to measure APX activity. The method proposed by Bates, et al. (1973) [31] was used to measure proline content.
CMT analysis [25, 32] was performed according to the method proposed by Martineau (1979) [32]. Cucumber seedlings growing their primary leaves were moved into growth chambers in which the temperature was set to 28°C, 38°C, or 43°C for a 24-hour dark period; subsequently, leaf discs were collected and subjected to treatment at high temperatures of 35°C, 40°C, 43°C, 45°C, 47°C, 49°C, 51°C, 53°C, and 58°C for 15 minutes. Subsequently, the leaf discs were used in the measurement of cell membrane relative injury (RI) by applying the following formula: RI (%) = {1− [1−(Ti/Tf)]/ [1−(Ci/Cf)]} × 100.
Statistical Analysis
A completely randomized design was applied for the treatments in this study. An analysis of variance was conducted using the general linear model procedure implemented in SAS software. The mean values of cultivars and temperature treatments were compared using the least significance difference procedure.
Results
Effects Of High Temperature on Cucumber Plant CMT
After the leaf discs of Wen nong 210, CU-127, and Ks No.3 were treated in water baths of various temperatures, the correlation coefficient (r2) between CMT and treatment temperature ranged between 0.80 and 0.86, indicating that the degree of heat-based RI inflicted on the cell membrane increased with the treatment temperature (Figure 1A). The Ks No. 3 cultivar consistently exhibited an RI degree of 20% or lower when it was treated at temperatures lower than 50°C. However, when the treatment temperature was increased to 50°C, the RI degree exceeded 50%. Wen nong 210 and CU-127 treated at 51°C had RI degrees of 40%–60% and less than 40%, respectively, indicating that CU-127 had high CMT. When the treatment temperature was higher than 51°C, the RI degree of Ks No. 3 exceeded 80%, whereas that of the other two cultivars remained at approximately 80% (Figure 1). According to the temperatures that induced RI degrees of 50% in the three cultivars, Wen nong 210 (54.1°C) exhibited the greatest CMT, followed by CU-127 (53.9°C) and Ks No. 3 (52.8°C). Ks No. 3 had the lowest CMT, rendering it the most sensitive to heat.
The experimental results revealed that when the Wen nong 210 seedlings were subjected to treatment temperatures higher than 51°C, seedlings that underwent treatment in the dark at 28°C sustained the greatest degrees of RI (81%–82%). By contrast, seedlings that were treated in the dark at 38°C and 43°C exhibited an RI degree of approximately 64%. This suggests that heat acclimatization helped reduce heat-based damage to Wen nong 210 (Figure 1B). When the CU-127 seedlings were subjected to treatment temperatures higher than 51°C, the RI degrees were in the range of 70%–80%. This suggests that CU-127 had high heat resistance (Figure 1C).
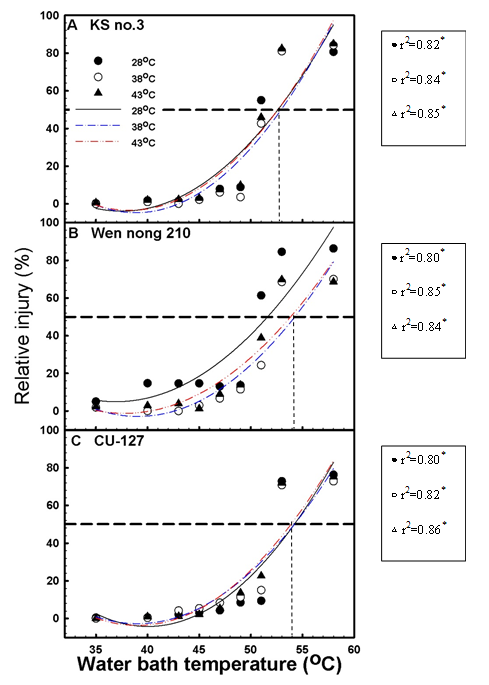
To obtain the thermo-response curve and relative injury (%).
Effects of High Temperature on Cucumber Plant’s Physiological Indices
Because the three cucumber cultivars were determined to have different heat resistance levels, this study further explored the effects of treatment at various temperatures on the physiological indices of the cultivars. Treatment at 28°C and 35°C engendered no notable difference in the chlorophyll content of the three cultivars. However, as the treatment temperature increased, the chlorophyll content of all three cultivars decreased significantly. Among the three cultivars, Ks No. 3 showed the lowest chlorophyll content, which was observed at 43°C and 45°C (Figure 2B). Regarding H2O2 content, at 28°C–43°C, Ks No. 3 exhibited higher H2O2 content levels than did the other two cultivars. However, at 43°C, the three cultivars exhibited no significant difference in H2O2 content levels, all of which were within 24–26 μmol (Figure 2A). Regarding MDA content, at 28°C, all three cultivars had MDA content levels of 8–13 μmol. However, the MDA content of Ks No. 3 increased significantly with processing time and peaked at 42.4 nmol at 45°C, approximately twice as high as those of the other two cultivars (Figure 2C).
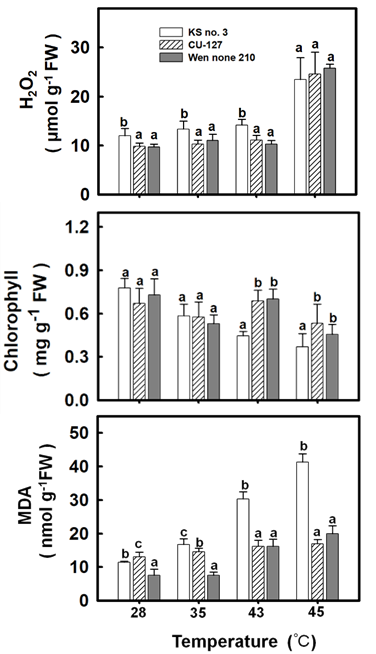
Effects of High Temperature on Cucumber Plants’ Antioxidant Enzyme Activity
Regarding physiological indices, both CU-127 and Wen nong 210 had high chlorophyll content levels but low MDA content levels. This might be associated with their antioxidation abilities. Accordingly, this study further examined the effects of high-temperature treatment on antioxidant enzyme activity. At 43°C, the CAT activity of Ks No. 3 was significantly weaker than that of Wen nong 210; however, the three cultivars showed little difference in CAT activity at other temperatures (Figure 3A). Furthermore, at 43°C, the APX activity of Ks No. 3 was approximately 0.82 units, which was significantly lower than that of the other cultivars. Additionally, at 45°C, the APX activity of Ks No. 3 was 1.1 units, which was still significantly lower than that of the other cultivars. Notably, at 45°C, the APX activity of Wen nong 210 was 3.42 units, which was 3.3 times higher than that of Ks No. 3 (Figure 3B).
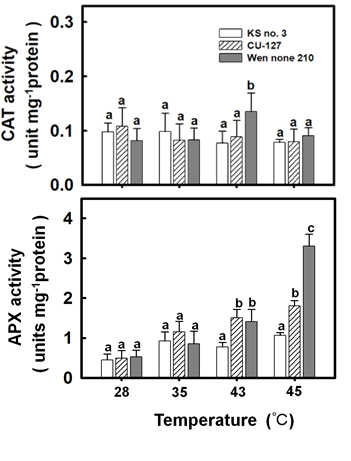
Effects of High Temperature on the Morphometric and Physiological Indices of Cucumber Plant
This study probed the effects of high-temperature treatment on the three cultivars’ morphometric and physiological indices, and the results are presented in Table 1. As the treatment temperature increased, the height, weight, and dry weight of seedlings (representing above- ground morphometric and physiological indices) decreased. However, for Wen nong 210 and CU-127, which exhibited greater heat resistance than the other cultivar, the cultivars’ height, weight, and dry weight observed at 45°C did not differ significantly compared with those observed at 28°C. Conversely, for the heat-sensitive Ks No. 3 cultivar, the cultivar height, weight, and dry weight decreased significantly at 45°C. This study also included the the root length, root weight, and root dry weight of plants (representing underground morphometric and physiological indices). The results revealed that at all treatment temperatures, the root length, root weight, and root dry weight of Ks No. 3 were higher than those of the other two cultivars. These main effect results indicate that temperature exerted a significant effect on the above-ground morphometric and physiological indices of seedling as well as their underground agronomic trait (i.e., root dry weight); moreover, the different cultivars differed significantly in above-ground and underground morphometric and physiological indices. However, this study revealed no significant interplay between temperature and species difference. Nevertheless, these results indicate that temperature and species difference had a significant effect on seedlings’ underground root dry weight (Table 1).
| Temperature | Variety | Shoot | Root | ||||
|---|---|---|---|---|---|---|---|
| Plant height -cm- | Seedling weight -g- | Dry weight -g- | Root length -cm- | Root weight -g- | Root dry weight -g- | ||
| 28℃ | KS no.3 | 7.2 | 0.69 | 0.09 | 18.15 | 0.660 | 0.041 |
| CU-127 | 6.9 | 0.99 | 0.14 | 15.35 | 0.650 | 0.045 | |
| Wen nong 210 | 8.1 | 1.09 | 0.13 | 14.70 | 0.523 | 0.042 | |
| 35℃ | KS no.3 | 6.9 | 0.92 | 0.11 | 27.85 | 0.783 | 0.049 |
| CU-127 | 6.3 | 0.91 | 0.11 | 16.63 | 0.633 | 0.048 | |
| Wen nong 210 | 7.5 | 1.18 | 0.15 | 15.38 | 0.665 | 0.050 | |
| 43℃ | KS no.3 | 5.9 | 0.75 | 0.09 | 22.40 | 0.690 | 0.041 |
| CU-127 | 5.9 | 0.75 | 0.11 | 17.83 | 0.668 | 0.040 | |
| Wen nong 210 | 5.9 | 0.61 | 0.09 | 14.18 | 0.578 | 0.034 | |
| 45℃ | KS no.3 | 5.9 | 0.56 | 0.07 | 19.15 | 0.728 | 0.052 |
| CU-127 | 5.8 | 0.88 | 0.11 | 17.68 | 0.723 | 0.034 | |
| Wen nong 210 | 7.2 | 0.85 | 0.12 | 14.33 | 0.458 | 0.030 | |
| Temperature | **z | ** | ** | ns | ns | ** | |
| Variety | ** | ** | ** | ** | * | * | |
| T vs. V | nsy | ns | * | ns | ns | ns |
Table 1: Responses of different morphometric and physiological indices of different varieties of cucumber seedlings at differe
z * and significant at P < 0.05 and 0.01, respectively. y ns not-significant at P < 0.05 Table 1:** Responses of different morphometric and physiological indices of different varieties of cucumber seedlings at different temperatures.
Effects of High Temperature on Proline Content of Cucumber Plants
When plants are subjected to environmental stress, they accumulate proline to enhance their resistance against such stress. This study thus assessed the effects of high temperature on the proline content of cultivars. According to Table 2, the proline content levels in the three cucumber cultivars increased progressively with the treatment temperature. At 28°C, the three cultivars showed no significant difference in proline content. At 35°C, the proline content of the heat- sensitive Ks No. 3 cultivar was 0.134 μmol/g, approximately 1.2 times higher than those of the other two cultivars. At 43°C, the proline content of Ks No. 3 was 0.172 μmol/g, still approximately 1.2 times higher than those of the other two cultivars. These results thus suggest that when Ks No. 3 encountered high temperature during its early growth period, it rapidly increased its proline content to enhance its heat resistance. When the treatment temperature was 45°C, the proline content levels of the three cultivars showed no significant difference, but the levels were all 1.73–1.9 times higher than those observed at 28°C (Table 2).
| Variety | Temperature | |||
|---|---|---|---|---|
| 28°C | 35°C | 43°C | 45°C | |
| KS no. 3 | 0.095a C | 0.134a B | 0.172a AB | 0.165a A z |
| CU-127 | 0.109a C | 0.119a C | 0.154a B | 0.188a A |
| Wen nong 210 | 0.097a C | 0.112a C | 0.149a B | 0.185a A |
| LSD 5% | 0.021ns y | 0.039ns | 0.034ns | 0.052ns |
Table 2: Proline content (μmol/g) in leaf after pre-treatment at 28, 35, 43 and 45℃ in cucumber ‘Ks no.3’, ‘CU-127’ and ‘Wen n
z. different capital letter means significant in temperature by F test at P<0.05. y. ns not-significant at P < 0.05 Table 2: Proline content (μmol/g) in leaf after pre-treatment at 28, 35, 43 and 45℃ in cucumber ‘Ks no.3’, ‘CU-127’ and ‘Wen nong 210’.
Evaluation of Temperature Resistance Test Parameters in Cucumber Seedlings
This study analyzed the mean values of chlorophyll, H2O2, MDA, CAT, APX, and proline content levels of the cucumber seedlings to determine the significance of their differences at different temperatures. Table 3 reveals that except for CAT content, all other parameters exhibited significant differences.
| Temperature | Chlorophyll -mg/g- | H O 2 2 -μmol/g- | MDA -nmol/g- | CAT Units/mg protein | APX Units/mg protein | Proline -μmol/g- |
|---|---|---|---|---|---|---|
| 28°C | 0.73 | 10.49 | 11.96 | 0.10 | 0.50 | 0.10 |
| 35°C | 0.56 | 11.54 | 12.95 | 0.09 | 0.98 | 0.12 |
| 43°C | 0.61 | 11.86 | 20.88 | 0.10 | 1.23 | 0.16 |
| 45°C | 0.45 | 23.83 | 26.05 | 0.08 | 2.05 | 0.18 |
| LSD 5% | 0.084** | 2.102** | 5.98** | nsy | 0.454** | 0.02** |
Table 3: Analysis of physiology factor levels, antioxidative enzyme activity and proline content of cucumber seedlings under d
z significant at P< 0.01. y ns, not significant at P< 0.01 Table 3:** Analysis of physiology factor levels, antioxidative enzyme activity and proline content of cucumber seedlings under different temperatures.
Discussion
When plants encounter high temperature or low temperature while growing, their chlorophyll synthesis is affected [33, 34]. Arun and Baishnab (1998) [35] observed that temperature-related stress affects the activity of chlorophyll synthase in cucumber and wheat plants, which in turn affects chlorophyll content. In the present study, the chlorophyll content of all three cucumber cultivars decreased as the treatment temperature increased. In particular, Ks No. 3 had a significantly lower chlorophyll content level but a higher MDA content level than did the other two cultivars. This suggests that high temperature promoted lipid peroxidation, thus reducing the chlorophyll content. Furtana and Tipidamaz (2010) [36] applied high concentrations of saline to salt-tolerant and salt-sensitive cucumber cultivars, and they revealed that the salt-tolerant cultivars exhibited a slower degradation of chlorophyll. Furthermore, they noted that the MDA content increased by 53% in the salt-tolerant cultivars and by 182% in the salt-sensitive cultivars. These observations suggest that plants’ cell membrane stability affects their resistance to environmental stress. Yeh and Lin (2003) [25] adopted CMT as the screening indicator of heat- resistant African daisy species; they discovered that seedlings of the heat-resistant Monte Crysto treated in 45°C–48°C water bath for 15 minutes sustained 50% RI at 54.1°C, which is considerably superior to those of other species. Moreover, the Fv/Fm values of Monte Cristo were superior to those of other species. Wen nong 210 used in the present study is one of the major cucumber cultivars cultivated in summer in Taiwan; this is because it has excellent resistance to high temperatures. The results of CMT analysis reveal that Wen nong 210 sustained 50% RI at 54.1°C and that CU- 127 sustained 50% RI at 53.9°C. By contrast, Ks No. 3 was sensitive to heat and sustained 50% RI at 52.8°C. These observations indicate that cucumber seedlings with high CMT are more resistant to high temperatures.
When plants encounter environmental stress, the content levels of free radicals such as O2 -, OH-, and H2O2 increase. Free radicals are ions in an unstable state, which drives them to grab electrons from the cell membrane, thereby affecting the membrane structure. This leads to lipid peroxidation, which in turn results in cell membrane disintegration [17]. In the present study, when the Ks No. 3 cultivar was treated at 28°C–43°C, its H2O2 content did not differ significantly from those of the other two cultivars, although its MDA content was significantly higher. This study inferred that the ability of antioxidant enzymes in clearing free radicals might have played a role in such variation. Accordingly, the activity of CAT and PAX was further analyzed. The results indicate that both Wen nong 210 and CU-127 had higher APX activity than did Ks No. 3, implying that in high-temperature environments, the APX activity of Wen nong 210 and CU-127 increased to reduce the H2O2 content, increase CMT, and alleviate chlorophyll degradation. This consequently increased the Wen nong 210 and CU-127 seedlings’ height, fresh weight, and dry weight relative to those of the Ks No. 3 seedlings. Previous research on cucumber plants under saline stress noted that when treated with high concentrations of saline, the CAT and APX activity of salt-tolerant cultivars could be rapidly increased to remove harmful free radicals, alleviating the reduction in dry weight and the degradation of chlorophyll [36]. This observation is in agreement with the results of the present study, suggesting that when cucumber plants encounter environmental stress, their stress resistance ability is affected by changes in chlorophyll, H2O2, and MDA content levels and APX activity.
When plants encounter environmental stress, the activity of antioxidant enzymes is increased to alleviate the damage caused by free radicals; the plants also generate osmoregulation substances to maintain cell membrane stability [37]. Osmoregulation substances, such as proline, are low-molecular-weight, nontoxic, and highly soluble compounds that regulate cellular osmosis, alleviate ROS- induced damage, and maintain cell membrane integrity under conditions of environmental stress [37]. Harsh, et al. (2016) [38] grew 37 species of cowpea in growth chambers for 1 week and then treated the sprouts at a temperature of 42°C for 1 hour. They reported that in 30 species, the activity of the antioxidant enzymes CAT and SOD increased significantly, and the proline content levels increased significantly. Similarly, authors have documented that the proline content and antioxidant enzyme (CAT, POD, and SOD) activity were previously found to have significantly increased in sorghum seedlings that had undergone high-temperature treatment (41°C) for 6 hours [39]. In the present study, the proline content of all three cucumber cultivars increased with temperature. In particular, at 35°C and 43°C, the accumulated proline content of Ks No. 3 was 1.2 times higher than that of the other two cultivars, signifying that the heat-sensitive Ks No. 3 rapidly increased its proline accumulation as soon as the environmental temperature started to increase. Mantovanini, et al. (2019) [40] observed that the proline content in the leaves of sugarcane plants in the CTC-2 strain increased by 58%; in addition, the biomass of the plants’ underground roots was increased to mitigate the poisoning of high-concentration aluminum. Authors have showed that providing proline supplementation to growing Arabidopsis thaliana plants accelerated root growth [41]. The findings of the aforementioned studies indicate that increased proline content in the above-ground portions of a plant can promote the growth of its underground roots. In the present study, at 28°C, the Ks No. 3 cultivar did not exhibit a significant difference in root length and root fresh weight when compared with the other two cucumber cultivars. However, at 35°C and 43°C, the proline content of Ks No. 3 was 1.2 times higher than that of the other two cultivars. Moreover, at these temperatures, Ks No. 3 exhibited significantly greater root length and root fresh weight than did the other two cultivars (Table 1); temperature and species difference appeared to have significantly affected the dry weight of underground roots. These observations suggest that proline is rapidly accumulated in the above-ground portion of the heat- sensitive Ks No. 3 cultivar in the event of high temperature, which also promotes the growth of its underground portion. Nevertheless, because of its weak CMT, Ks No. 3 cannot achieve rapid enhancement of antioxidant enzyme activity to remove harmful free radicals promptly. Extensive research is required to verify this inference.
Conclusion
This study selected three cucumber cultivars with varying heat resistance levels as raw materials and used their seedlings for heat resistance screening. CMT analysis was conducted to test the stability of their cell membrane systems when subjected to high temperatures. Numerous physiological indices and antioxidant enzyme activity levels were subsequently measured. There is a significant difference between the test parameters and the processing temperature. These results indicate that the optimal timing for cucumber plants to receive high-temperature treatment is when a seedling grows its primary leaf. Furthermore, the content levels of chlorophyll, H2O2, MDA, and proline, as well as the activity of the antioxidant enzyme APX, were found to be suitable indicators of cucumber plant heat resistance.
References
-
Wahid A, Gelani S, Ashraf M, Foolad MR (2007) Heat tolerance in plants: An overview. Environmental and Experimental Botany 61(3):199-223.
-
Meng LB, Li SM (2003) On the influence of high temperature on cucumber’s physical and chemical development. Journal of Harbin University 24: 121-125.
-
Han, YK, Du SL, Zhang GH, Wei AM, Liu N, et al. (2009) Screening of heat-tolerance of cucumber at seedling Stage. China Cucurbits Vegetables 22: 1-4.
-
Wang YH, Lin TK, Lin JN (2011) Breeding and cultural technologies of cucurbits in the changing agricultural environment. Proceedings of the Workshop on Crop Breeding and Management of Agricultural Environment for Coping with Climate Change Taiwan Agriculture. 156:153-161.
-
Apel, K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review Plant Biology 55: 373-399.
-
Bhattacharjee S (2005) Reactive oxygen species and oxidative burst: Roles in stress, senescence and signal transduction in plants. Current in Science 89(7): 1113- 1121.
-
Bolwell GP, Wojtaszek P (1997) Mechanisms for the generation of reactive oxygen species in plant defence–a broad perspective. Physiological and Molecular Plant Pathology 51(6): 347-366.
-
Gechev TS, Van Breusegem F, Stone JM, Denev I, Laloi C (2006) Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 28(11): 1091-1101.
-
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of Botany, pp: 26.
-
Hodges DM, Forne CF (2000) The effects of ethylene, depressed oxygen and elevated carbon dioxide on antioxidant profiles of senescing spinach leaves. Journal of Experiment Botany 51(344): 645-655.
-
Salin ML (1988) Toxic oxygen species and protective systems of the chloroplast. Physiologia Plantarum 72(3): 681-689.
-
Liu X, Huang B (2000) Heat stress injury in relation to membrane lipid peroxidation in creeping bentgrass. Crop Science 40(2): 503-510.
-
Ya’acov YL (1988) Plant senescence processes and free radicals. Free Radical Biology Medicine 5(1): 39-49.
-
Dhindsa RS, Plumb-Dhindsa P, Thorpe TA (1981) Leaf senescence: correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. Journal of Experiment Botany 32(1): 93-101.
-
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Archives of Biochemistry and Biophysics 125(1): 189-198.
-
Hodges DM, Andrews CJ, Johnson DA, Hamilton RI (1996) Antioxidant compound responses to chilling stress in differentially sensitive inbred maize lines. Physiologia Plantarum 98(4): 685-692.
-
Choudhury B, Chowdhury S, Biswas AK (2011) Regulation of growth and metabolism in rice (_Oryza sativa_ L.) by arsenic and its possible reversal by phosphate. Journal of Plant Internatioal 6(1): 15-24.
-
He XM, Lin YE, Chen QH, Deng JM (2002) Effects of heat stress on seedling growthproline contents and SOD activity in cucumbers. J. Shanghai Jiaotong Univsity 20(1): 30-33.
-
Shi Q, Bao Z, Zhu Z, Ying Q, Qian Q (2006) Effects of different treatments of salicylic acid on heat tolerance, chlorophyll fluorescence, and antioxidant enzyme activity in seedlings of _Cucumis sativa_ L. Plant growth regulation 48: 127-135.
-
Xu XY, Yu JH, Xie JM (2007) Effects of high temperature stress on chlorophyll fluorescence quenching in cucumber leaves. Journal of Gansu Agricultural University 42: 60-62.
-
Ngaklunchon R, Huang WL, Liu ML, Tai SF, Chi HS (2010) Change of antioxidant enzyme activities during heat stress in cucumber (_Cucumis sativus_ L.). Journal of Agriculture and Forestry, National Chiayi University 7: 118-134.
-
Zhang, HM, Jin HJ, Ding XT, Yu JZ (2011) Physiological reaction and chlorophyll fluorescence characteristics of cucumber seedlings under hot stress. Journal of Shanghai Jiaotong University. (Agric. Sci.) 29: 61-66.
-
Chen WL, Yang WJ, Lo HF, Yeh DM (2014) Physiology, anatomy, and cell membrane thermostability selection of leafy radish (_Raphanus sativus var. oleiformis_ Pers.) with different tolerance under heat stress. Science Horticultural 179: 367-375.
-
Shih PY, Shih JC, Chang LC, Lo HF (2013) Physiological index for tolerance to high temperature and waterlogging in cauliflower_._ Journal of Taiwan Social Horticulture Science 59:191-204.
-
Yeh DM, Lin HF (2003) Thermostability of cell membranes as a measure of heat tolerance and relationship to flowering delay in chrysanthemum. Journal of the American Society for Horticultural Science 128(5): 656-660.
-
Liu ML, Hsieh CH, Chao YY (2017) Kaohsiung No. 3 cucumber: an early flowering variety tolerant to heat and moisture. Horticultural Science 52(10): 1435-1437.
-
Wintermans J, De Mots A (1965) Spectrophotometric characteristics of chlorophylls a and b and their phenophytins in ethanol. Biochimica et Biophysica Acta 109(2): 448-453.
-
Jana S, Choudhuri MA (1981) Glycolate metabolism of three submersed aquatic angiosperms: effect of heavy metals. Aquatic Botany 11: 67-77.
-
Kato M, Shimizu S (1985) Chlorophyll metabolism in higher plants VI. Involvement of peroxidase in chlorophyll degradation. Plant Cell Physiology 26(7): 1291-1301.
-
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant cell physiology 22(5): 867-880.
-
Bates L, Waldren R, Teare I (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39: 205-207.
-
Martineau J, Specht J, Williams J, Sullivan C (1979) Temperature tolerance in soybeans. I. evaluation of a technique for assessing cellular membrane thermostability. Crop Science 19(1): 75-78.
-
Feierabend J (1977) Capacity for chlorophyll synthesis in heat-bleached 70S ribosome-deficient rye leaves. Planta 135: 83-88.
-
Hasselt PR, Strikwerda J (1976) Pigment degradation in discs of the thermophilic _Cucumis sativus_ as affected by light, temperature, sugar application and inhibitors. Physiologia Plantarum 37(4): 253-257.
-
Arun KT, Baishnab CT (1998) Temperature-stress- induced impairment of chlorophyll biosynthetic reactions in cucumber and wheat. Plant Physiology 117(3):851-858.
-
Furtana GB, Tipirdamaz R (2010) Physiological and antioxidant response of three cultivars of cucumber (_Cucumis sativus_ L.) to salinity. Turkish Journal of Biology 34: 287-296.
-
Ashraf M, Foolad MR (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environment Experiment Botany 59(2): 206-216.
-
Harsh A, harma YKS, Joshi U, Rampuria S, Singh G, et al. (2016) Effect of short-term heat stress on total sugars, proline and some antioxidant enzymes in moth bean (_Vigna aconitifolia_). Annals of Agricultural Science 61(1): 57-64.
-
Gosavi GU, Jadhav AS, Kale AA, Gadakh SR, Pawar BD, et al. (2014) Effect of heat stress on proline, chlorophyll content, heat shock proteins and antioxidant enzyme activity in sorghum (_Sorghum bicolor_) at seedlings stage. Indian Journal of Biotechnology 13(3): 356-363.
-
Mantovanini LJ, Silva RG da, Silva JDOL, Rosa TM, dos Santos DMMD, et al. (2019) Root system development and proline accumulation in sugarcane leaves under aluminum (Al3+) stress. Australia Journal of Crop Scice 13(2): 208-213.
-
Biancucci M, Mattioli R, Moubayidin L, Sabatini S, Costantino P, et al. (2015) Proline affects the size of the root meristematic zone in Arabidopsis. BMC Plant Biology 15: 263-277.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note