Recovery of Cryopreserved Buds of Chinese Yam (Dioscorea Polystachya Turcz.) Observed under Scanning Electron Microscope (Sem)
Early development of cryopreserved Chinese yams’ (Dioscorea polystachya Turcz) explants were studied under scanning electron microscope (SEM). A combination of vitrification and droplet protocol was implemented with 3 weeks cold acclimation in an alternating temperature of 28°Cday/5°C night and 15% sucrose in the pre-culture medium. Survival in the form of shoot and swelling at the axil or at the near apex of original leaf bud was observed at 3 days after re-culture (rewarming). At 7 days after re-culture, development of new axillary shoot, elongation and enlargement of the surviving green structures as well as addition of new tissues to the previously developed callus at terminal bud were observed. Shoot differentiation from green structures was observed at 16 days after re-culture.
Introduction
Along with the implementation of cryopreservation method in routine conserva- tion of various crop species [1], studies on protocol development have been continued. A good cryopreservation protocol should not only increase the quantity but also the quality of surviving explants [2] through direct recovery of shoot instead of via callus.
SEM has been very instrumental for plant morphologists in investigating the surface features of numerous organisms and their organs, due to its properties of great depth of field and the ability to image large, topographically complex specimens in three dimensions at a range of magnifications [3]. Successful use of SEM in supporting the cryopreservation related study has been demonstrated for some crop species including potato [4], chrysanthemum [5]
and carrot [6].
Using a combination of vitrivication and droplet methods, a cryopreservation study on D. polystachya was able to obtain about 70% survival and 40% shoot/plant regeneration [7]. Apical buds explants produced by this genotype are also relatively simple to prepare for SEM and their number per plantlets could be readily estimated (with low standard error) prior to cultivation. This genotype is therefore practical to be used as an experimental material.
This study intends to observe the early stages of development and describe morphological features of surviving buds of D. polystacya under SEM folowing cryopreservation, in comparison to that of PVS2 treated and plain in vitro culture at their early stages of development.
Materials and Methods
Cryopreservation Procedure
A combination of vitrivication and droplet cryopreservation protocol was imple- mented in this study. The detail protocol was described in Leunufna S, et al [8]. In vitro plantlets of Yam 21 (Dioscorea polystachya) cultivated for 2 months in Murashige and Skoog (MS) [9] medium supplemented with 2.0 µM Benzylaminopurine (BAP), 0.1 µM Naphtaleneacetic acid (NAA) (M2 medium) with 3% sucrose, 0.2 mg/l activated charcoal (AC) and 1% agar, in a culture room of 25/27°C, 60-80 µmol cm-2s-1 light intensity and 16h/8h light/dark periods using fluorescent lamps. Selected explants were further cold acclimated in a growth chamber at an alternating 28°C/5°C and 12h thermo-photo-periods with light intensity of 60-80 µmol cm-2s-1 provided by fluorescent lamps. Following cold acclimation for 3 weeks period, the plantlets’ apical buds were dissected under a light microscope (binocular) to 2-4 mm in length and further preculture in M2 medium with 15% sucrose.
The explants were further loaded for 20 min in loading solution followed by dehydration in plant vitrification solution (PVS) 2 for 20 min and immersion into liquid nitrogen (LN) in droplets of 7.5 μl PVS2. After rewarming for 3 min in MS, 3% sucrose solution and unloaded in M2, 40% sucrose for 5 min, they were cultivated in M2 medium followed by cultivation in MS medium without hormone one week after.
Sample Preparation for SEM Observation
Total samples used for the main experiment were 50 explants including 20 cryo- preserved (+LN), 10 PVS2 treated (-LN), and 10 normal in vitro (control) explants.
Survival rates obtained were 60% for cryopreserved, 90% for PVS2 treated and 100% for control explants. In addition to the main experiment, other small experiments were conducted to obtained additional samples. Overall, 3 to 6 samples were observed for each treatment at each stage of development. Observation was conducted at 3 days, 7 days and 16 days after re-culture. Before preparation for the SEM, general conditions of the explants were recorded under the light microscope (binocular).
Samples for SEM were trimmed to about 2 mm and fixed overnight in a fridge (4°C) in 0.05 M cacodylate buffer containing 2% glutaraldehyde and 2% formal- dehyde at pH 7.2. Fixed samples were than stored in a fridge in 0.05 M cacodylate buffer until further use. Samples were than degassed before washed twice, each for 15 min with fresh 0.05 M cacodylate buffer followed by dehydration in a series of solution comprising 30%, 50%, 70%, 90% and 100% ethanol. Dehydration process was conducted 2 times and each treatment lasted 15 min. Samples were then transferred into a Bal-Tec CPD030 critical point dryer (Bat-Tec Ag, Liech- tenstein) and critical point dried. Dried samples were gold coated in an Edwards S150B sputter coater (Edwards high vacuum inc., Crawley, West Sussex, UK) and examined in a Hitachi S4100 SEM (Hisco Europe, Ratingen, Germany).
Results
Natural Shoots and Buds Morphology, and Developmental Pattern
Visual observation of morphological features of yams natural shoot development revealed that beside the terminal shoot, a number of buds can arise from the axillary node (Figure 3).

Figure 1: In vivo (from glass house grown plants) shoot developmental pattern of the four yams species and in vitro explants used for cryopreservation. Terminal shoot (a-e), main and serial buds (f, g, i, j), aerial tuber’s primordium (arrow head - h) and in vitro buds to be cryopreserved (k, l). D. bulbifera (a, f, h, k), D. polystachya (b, g, l), D. alata (c, i) and D. cayenensis (d, e, j). Bars indicate 1 mm.
These buds have been designated as main bud (one) and more than one serial buds [10] (Figure 2), which include the buds developed to form an aerial tuber and at the aerial tuber. The development of new shoots after cryopreservation resembles this system since the explants used were buds at the upper most node (Figure 3 k,l).
![Figure 2: Illustrative developmental pattern of serial buds on D. polystachya. Primary and serial buds (a), aerial tuber containing serial buds (b) [10], primary and serial buds (c) primordia as observed under light microscope. B leaf/leaf primordium. H main stem, J primary/main bud, S, S1 and S2 serial buds.](/fulltextimages/6071/fig_2.jpeg)
Morphological Features at Three Days After Re-culture
The new emergence of the cryopreserved explants appeared at two sites: at the axil between the young leaf (leaf primordium) and the petiole (inner core of the bud), and at the center or near apex of the leaf primordium (Figure 3).

Figure 3: Morphological features of D. polystachya explants three days after re-culture. Surviving cryopreserved explants showing structures developed at the axil (main bud – a, b), swelling of the leaf primordium (c), and green (under light microscopic observation) original leaf primordium with a small swelling at the side, which eventually turn brown in colour and die (d). Surviving PVS2 treated explant (green in colour under light microscope - e), and dead explant (f - brown in colour under light microscope). sp shoot pri- mordium, bp bump (green structure under light microscope), s swelling. Bars indicate 300 µm (a), 400 µm (b-e), 100 µm (f).
These new growths took the form of shoot at the axil (Figure 3a), bump at the axil (Figure 3b) or swelling of the original leaf primordium (Figure 3c) and could already be observed at three days after re-culture. Most of the original young leaf did not survive, show- ing brown in colour. In some cases, the original leaf promordium remained green with or without a small swelling (Figure 3d) but eventually turned brown. PVS2 treated explants mostly remained green at 3 days after re-culture and exhibited similar features as that before treatment (Figure 3e), however, some explants showed suppressive growth resulting in the appearance of a swelling near the apex of leaf bud similar to that shown by survival cryopreserved explant (picture not shown).
Although relatively very low in number in comparison to those of cryopreserved ones, dead explants were also observed after PVS2 treatment (Figure 3f). There were no noticeable changes on the in vitro (control) explants at three days after culture (they remained green in colour).
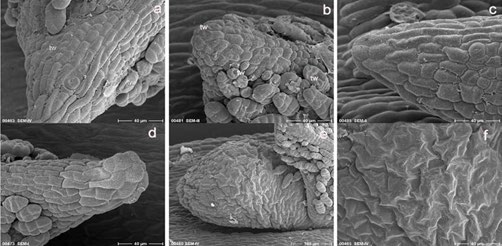
Figure 4: Morphological features of surviving and dead explants at three days after recul- ture. Dead cryopreserved (a) and PVS2 treated (b) leaf primordium (explants) show small wrinkles at various sites at the tissue’s surface and at trichomes, whilst surviving PVS2 treated (c) and in vitro culture (d) explants exhibit turgened (normal) cells at surface (epidermis) of the tissues. Newly developed axillary structure of cryopreserved explants exhibit highly wrinkled surface (d, f); tw, wrinkle. Bars indicate 100µm (e), 40µm (the rest).
Remarkable morphological differences were not observed on the surface tissue of original leaf buds (explants) between the survived and the dead explants. However, closer look indicated small wrinkles appeared at various sites of the tissue surface and at trichomes of the death cryopreserved (Figure 4a) and PVS2 treated (Figure 4b) leaf buds. These features did not or very little appeared on the surviving leaf buds of PVS2 treated (Figure 4c) and on in vitro cultured leaf buds (Figure 4d). On the tissues’ surface of the new axillary developed structures of cryopreserved explant, however, highly wrinkled tissues’ surface were present (Figure 4e, f).
Development at 7 Days after Re-culture
The emergence of new axillary buds was one of the features observed at seven days after re-culture on cryopreserved explants (Figure 5a). The previously developed axillary structure showed further elongation and enlargement (increase in size – Figure 5b). Newly developed fresh and smooth tissues structures were added to the previously developed callus from near apex of the original leaf bud (Figure 5c). Surviving old tissues of cryopreserved leaf buds seven days after rewarming also showed a disappearance of wrinkles in comparison to that of three days after re- warming (Figure 5d, e). Although the leaf buds of PVS2 treated (-LN) explants were still alive seven days after re-culture, their development was slow (Figure 5f). Rapid growth was shown at the base part and later took over (replaced) the slowly developed old leaf buds (Figure 5g). Other explants showed normal growth and development. In vitro cultured explants showed normal enlargement of the original leaf buds followed by initiation of new leaf buds primordium from the axil after seven days of culture (Figure 5g,h).
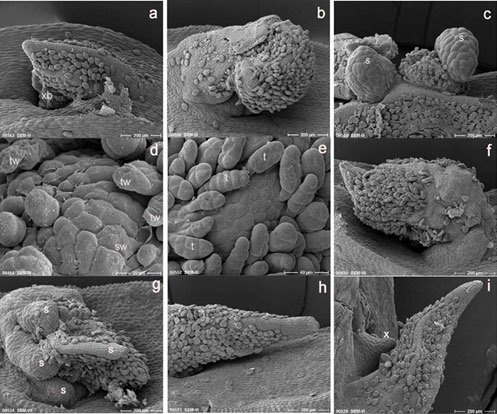
Figure 5: Morphological features one week after re-culture (a-i). An axillary leaf bud developed from the dead original leaf primordium (a), enlargement of the axillary developed structure (b) and development of new tissues on the previously developed tissue structures from the near apex of the original leaf primordium (c) of the surviving cryopreserved explants. Fresh and smooth epidermal tissues observed on the seven days re-cultured surviving cryopreserved explants (e) in comparison to several wrinkled and broken trichomes on the surface of three days old surviving explants (d). Slowly developed surviving original leaf primordium (f) and rapidly elongated tissue from the base part of the leaf primordium (g) after PVS2 treatment. Elongation and enlargement of the original leaf primordium (h) and newly developed axillary bud primordium (i) exhibit by in vitro cultured explants; xb, axillary bud, s, newly developed tissue structures, tw, wrinkled trichomes, sw, slightly rough (wrinkled) tissue surface, t, trichome, x, axilary bud pri- mordium. Bars indicate 20 μm (d), 40 μm (e) and 200 μm (the rest).
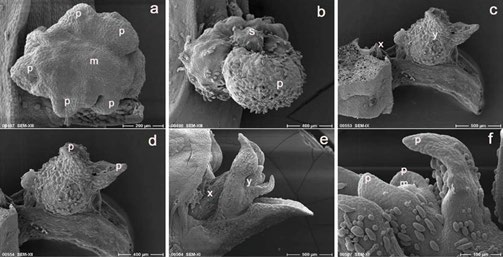
Figure 6: Explants development 16 days after re-culture (a-f). Advanced lateral swelling of leaf primordia surrounding the apical dome (a) and development of a smooth green structure (shoot) following the advanced growth of the leaf primordium (b) mark the initial stages of shoot formation and eventually plantlet development from the previously developed green structures of cryopreserved explants. Further development of the PVS2 treated explants indicating initial shoot growth at the axil (c) and bud with two leaf pri- mordia at the near apex (terminal) of the original leaf (c and d). An axillary bud with two leaf primordia (e) and a terminal bud with four leaf primordia (e and f) are shown by the in vitro cultured explants. p, leaf primordium, m, apical dome, s, shoot, x, axillary bud, y, terminal bud. Bars indicate 100µm (f), 200µm (a), 400 µm (b, d), 500 µm.
Development at 16 Days after Re-culture
Various stages of growth and development were exhibited by the explants of the three different treatments. The latest development of cryopreserved explants showed differentiation into the formation of new shoot at the previously developed green structure, initiated with the advanced lateral swelling on the sides of the structure indicated the development of leaf primordia surrounding the meristematic dome (Figure 6a). A more advanced development was indicated by the formation of small, fresh and smooth green structure on the previously developed tissue structure (leaf primordium–Figure 6b). These types of structures later developed into shoots and eventually into plantlets. The most advanced development of PVS2 treated explants was shown by the presence of new shoot with two leaves developed at the terminal leaf (near apex of the original leaf – Figure 6c,d). New shoot was also observed at the axil of the original leaf but with lesser advanced in development. The difference in development between axillary and terminal (developed from the original leaf) buds was also seen on the in vitro culture explants (Figure 6e,f). The terminal bud at 16 days after culture produced shoot with four leaves whilst axillary bud was on the two leaves stage. The terminal bud of the in vitro culture explant marked the most advanced developmental stage of all explants at 16 days after treatment.
Discussion
The meristematic tissues on the explants (apical leaf buds) used for cryopreservation are present at both axillar and terminal bud primordia (near the apex of the leaf), which enable the possible resumption of growth at the two sites. Rapid development was shown by the terminal buds as indicated by the in vitro culture (control) and most of the PVS2 treated explants (>90%). This rapid development leads to suppression of the growth of the axillary bud primordium due to the apical dominance. For the cryopreserved explants, however, the resumption of growth appeared mainly from surviving cells at the meristematic tissues present at the axils or near apex of the leaf. The original leaf usually dies as also observed by other authors [5, 11].
The fact that survival through callus formation was frequently observed at the terminal buds in comparison to that at the axillary buds might be attributable to the unprotected (exposure) position of the former to the cryopreservation treatment, which leads to more cellular injuries. Development of either shoots or calluses after cryopreservation is determined by the extent of damage taken placed during the application of cryopreservation protocol. Insufficient amount of undam- aged cells present after rewarming could hindered the reorganization of cells into a functional shoot [4]. This could also partly explain the reduction in the percentage of surviving explants developed into shoot.
The surviving and dead explants of cryopreserved potato shoot tips exhibited similar morphological features during the period of one day after rewarming as observed by [4]. Fukai and Oe [5], however, observed many small wrinkles on the dead shoot tips of Chrisanthenum immediately after rewarming. The slightly rough tissue’s surface and wrinkled trichoms observed on the original leaf bud of cryopreserved and PVS2 treated explants after 3 days of re-culture could indicate the shrinkage of cells as an effect of dehydration through PVS2 application. Considerable wrinkled tissue observed on the new axillary developed structure of cryopreserved explants in this study was similar to that shown by slow and rapid drying somatic embryos of Daucus carota [6]. Further observation by Tetteroo FAA, et al. [6] revealed that the extent of shrinkage was associated with the initial dry matter content of the cells and that cell shrinkage per se, does not induce lethal injuries during dehydration, but upon re- hydration, the non-viable somatic embryo had lost turgor and intracellular organization, whilst viable embryo had turgescent cells with intact cell structures.
Observation on Pea meristem [11] indicated that at 7, 9 and 11 days after re-culture, the old dead tissues on the original meristem of the survival cryopreserved explants have been pushed aside and compressed by masses of growing tissue and no meristematic dome was observed at this stage. Similar phenomenon was seen in this study, in which growth resumption continued one week after re-culture with additional growth of callus, replacement of rough and wrinkled tissue surface with smooth and turgescent tissue surface, elongation and enlargement of the developed green structure or the appearance of new axillary shoot.
Although shoot development can be observed at one week after re-culture (Figure 3c), development of organized growth (shoot primordium) on cryopreserved ex- plants of D. polystachya after green structures or after callus, seems to take place about two weeks after re-culture (Figure 4a,b). For Potato (solanum goniocalix), the expansion of leaf primordum from the surviving explants appeared within 7-10 days [4] whilst for Chrisantemum, initiation of shoots through small bump on the leaf primordium or green spot, which appeared on the rough surface callus were observed after 20 days of re-culture [5].
While cell differentiation into the formation of new axillary bud can be readily observed on the in vitro culture explants at one week after culture in addition to further growth of the original leaf bud, the PVS2 treated explants, suffering from the dehydration stress, showed a slower and abnormal development of the original leaf bud. The effect of cryoprotectant (PVS2) on redevelopment of the explant was also visible at 2 weeks after re-culture. Fukai and Oe [5] reported that al- though all explants developed into plantlets without callus formation, 46% ab- normal vitrified leaf for Chrisantemum shoot tips was observed following 2 days pre-culture in 5% DMSO continued with 1 h treatment with 10% DMSO solution at 0°C. PVS2 treated explants of Dioscorea polystachya for 20 min showed 93% survival and all explants developed into shoot and plantlets after 1 to 2 months re-culture [7]. In addition to 13.7% sucrose, 30% glycerol and 15% ethylene glycol, PVS2 contain 15% DMSO.
In view of the quantity and quality of recovery of D. polystachya in this and pre- vious studies, an improvement of the protocol is needed to reduce injuries and to enhance the amount of surviving cells, and therefore helping their reorganization into shoot formation. Since callus formation might lead to alteration in genetic constitution, further study aimed at verifying the genetic stability is necessary.
Conclusion
This study reveals that the recovery of cryopreserved explant of D. polystachya is visible less than 3 days after rewarming in the form of shoot, bump at the axil or swelling of the original leaf primordium, developed from a few surviving cells at the meristematic tissues. Even though, shoot and eventually plantlet can develop from both sites, axillary growth seems to provide more direct formation into shoot rather t h a n t h r o u g h callus i n comparison to swelling of the original leaf primordium. Cell differentiation on the surviving green structure or callus into shoot formation is initiated at about two weeks after re-culture. In vitro culture and most of the PVS2 treated explants resume growth from the original bud and develop into plantlets. However, PVS2 induces osmotic stress to the explants which resulted in slower and some abnormal growth. Due to the possibility of recovery through callus formation, further experiment to improve the protocol and to verify the genetic stability of cryopreserved explants is suggested.
Acknowledgements
The authors thank Mrs. W. Panitz and Mrs. M. Wiessner for the technical assistance in explants preparation for TEM, and Mrs. D. Büchner for in vitro propagation of the experimental materials. The first author thanks DAAD for the funding provided.
References
-
Reed BM (2002) Implementing cryopreservation for long-term germplasm preservation in vegetatively propagated species. In: Towill LE, Bajaj YPS (EDs.), Biotechnology in agriculture and forestry vol. 50. Cryopreservation of plant germplasm II. Springer-Verlag Berlin, pp: 22-33.
-
Benson EE (1990) Free radical damage in stored plant germplasm. IBPGR, Rome, ISBN 10: 92-9043-196-2.
-
Vesk M, Dibbayawan TP, Vesk PA, Egan EA (2000) Field emission scan- ning electron microscopy of plant cells. Protoplasma 210: 138-155.
-
Grout BWW, Henshaw GG (1980) Structural observation on the growth of potato shoot-tip cultures after thawing from liquid nitrogen. Ann. Bot 46(2): 243-248.
-
Fukai S and Oe M (1990) Morphological observation of Chrysanthemum shoot tips cultured after cryopreservation. J Jap Soc Hort Sci 59(2): 383-387.
-
Tetteroo FAA, van Aelst AC, von Recklinghause IR, Golovina EA and Hoekstra FA (1998) Membrane permeability, morphology, and desiccation toler- ance of Daucus carota, somatic embryos as influenced by drying rate. Proto- plasma 202: 202-212.
-
Leunufna S, Keller ERJ (2003) Investigating a new cryopreservation protocol for yams (Dioscorea spp.). Plant Cell Rep 21(12): 1159-1166.
-
Leunufna S, Keller ERJ (2005) Cryopreservation of yams using vitrification modified by including droplet method: Effects of cold acclimation and su- crose. Cryo Letters 26(2): 93-102.
-
Murashige T and Skoog F (1962) A revised medium for rapid grow and bioassays with tobacco tissue culture. Physiol Plant 15: 473-497.
-
Troll (1954) Practical introduction to plant morphology. Fisher-Verl, Jena.
-
Haskins RH, Kartha KK (1980) Freeze preservation of pea meristem: cell survival. Canadian Journal of Botany 58(8): 833-840.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note