In Vitro Multiplication Rate and Pot Yield of Cryopreservation Derived Buds of Yams (Dioscorea Spp)
One of the principal objectives of the germplasm preservation is to maintain their unique genes combinations. A study was conducted in the laboratory and glass house to evaluate the multiplication rate of in vitro cultured plantlets, several morphological and yield characters of glass house pots grown plants derived from cryopreservation using modified droplet method on yams (Dioscorea spp.). In vitro MR was similar among in vitro culture, PVS2 treatment and cryopreservation derived plantlets for D. polystachya and D. cayenensis. In vitro only derived plantlets exhibited significantly lower MR in comparison to the two other treatment variants for D. bulbifera. This was, however, could be attributed to physiological effects which will be recovered in further sub-culture cycles. The morphological characters, number of tuber per plant and tuber' harvest weight per plant were constant for plant derived after cryopreservation and in vitro culture (control) for D. cayenensis. Modified droplet method is, therefore, reliable for the cryopreservation of yams germplasm and maybe other crops.
Introduction
In vitro maintenance has been one of the methods implemented in the conservation of yams [1, 2, 3, 4, 5]. This method reduces the risk of germplasm losses due to operational error, pest and diseases, climatic changes, as well as the demand for ground area and labour intensive, beard by the field maintenance which is traditionally implemented in the conservation of seedless, recalcitrant and semi-recalcitrant seeds plants [6, 7, 8, 9, 10].
There is, however, some discussion, that alteration in the genetic constitution of the conserved materials might occur through the in vitro maintenance. Genetic variability generated during tissue culture (somaclonal variation-[11]) is well documented [12]). Imposition of environmental stress or minimal growth during the in vitro maintenance as well as type of plant materials used and regeneration modes such as callus and protoplast are some of the frequent factors which could lead to the genetic changes [12, 13]). Harding [14, 15] found that potato shoot tips grown in mannitol- supplemented medium, indicated a DNA hypermethylation, which could be attributable to an adaptive response to condition of high osmotic stress. Since the methylation status may be inherited and alter the phenotypes of the subsequent progenies, this finding has a significant implication to the use of tissue culture techniques in the genetic conservation of potato germplasm [15].
Assuming that all physiological processes are in a ‘stand still’ stage at cryogenic temperature (-196 for LN -[16]), the materials cryopreserved should retain their genetic constitution upon recovery. This assumption, however, has to be empirically verified since cryopreservation involves many steps including tissue culture as a transit phase, preconditioning, cryoprotection, cooling and rewarming, recovery and plant regeneration [14] each of which has its own effect on the genetic stability even with the use of explants reportedly stable such as meristem shoot tips [12]. For instance, dimethylsulfoxide (DMSO) in 2 to 10% solution was predicted to be involved in generating a variety of genetic and/or epigenetic changes [17].
Assessment of genetic stability has been performed at the genetic level covering chromosomes and restriction enzymes generated DNA fragment analysis as well as at the phenotypic level including protein electrophoresis, secondary products and morphological (quantitative and qualitative) characters [18, 19]). Studies regarding genetic stability of cryopreseration derived materials have been implemented on a number of plant species including some medicinal plants [20], strawberry and raspberry [21, 22]), citrus [23]), sugarcane [24], kiwi and grape [25], banana [26]) and yams species of D. deltoidea [27], D. bulbifera [28]) and D. floribunda [29]. The most intensive study, however, has mainly involved potato (Solanum tuberosum L.) germplasm where various techniques have been implemented [14, 15, 19, 30, 31, 32, 33, 34]. A wide and in-depth discussion regarding genetic stability after cryopreservation is provided by Harding [35].
Three of four species of Dioscorea spp. used in our previous study in refining the protocols for yams species using modified-droplet method were successfully recovered into plantlets and transplanted to the glass house. The present report reveals the results of the experiments comparing the in vitro multiplication rate (MR) of cryopreserved (+LN), cryoprotectant treated (-LN) and plantlets derived from in vitro culture only as well as several phenotypic characters including morphological and yield characters, observed on the glasshouse-grown plants derived from in vitro culture and cryopreserved apical buds.
Materials and Methods
Plant Materials
Study on the in vitro MR involved three genotypes of yams representing three species, namely Yam 16 (D. bulbifera L.), Yam 21 (D. polystachya Turcz.) and Yam 58 (D. cayenensis Lam.) whilst the study on morphological and yield characters was done only on Yam 58 (D. c_a_yenensis). The history, the in vitro establishment and maintenance of the materials were described in detail in the previous reports [36, 37, 38].
Cryopreservation Procedure
Six weeks old in vitro plantlets cultivated in MS [39]) medium supplemented with 2.0 mg/l 6-benzylaminopurine (BAP), 0.1 mg/l α-naphtaleneacetic acid (NAA) designated as M2 medium plus 3% (w/v) sucrose, 0.2 mg/l activated charcoal (AC) and 1% (w/v) agar were further cold- acclimated for 3 weeks in an alternating thermo-photo- period of 28/5 °C, 12 h. Previously, the materials have been cultivated in the maintenance room at constant temperature of 25-27 °C, 16h/8h light/dark periods and light intensity of 60-80 µmol.m-2.s-1 provided by fluorescent lamps. Following cold acclimation, the apical buds (plus first axillary nodes for D. bulbifera) were dissected under the light microscope to 2-4 mm in length and used as explants. Subsequent preculture in 15% sucrose was applied for the explants of D. bulbifera and D. polystachya, and 10% (w/v) for D. cayenensis, all in M2 medium with 0.7% (w/v) agar.
After a three-days preculture period, the explants were loaded in solution A (loading solution) for 20 min, followed by in PVS2 for 20 min [40]. The explants were than rapidly cooled in liquid nitrogen after placement in droplets of PVS2 on aluminium foil. Rewarming was done by quickly emerging the explants into MS solution with 3% (w/v) sucrose. The explants were than cultivated in semi-solid medium M2 for one week in dark. Further cultivation was done in semi-solid medium MS without hormones plus 3% sucrose. The same medium was regularly renewed in two-weeks intervals, and the shoot recovery from survived explants was observed up to 4 months. The explants (cooled and control) showing shoot development were transferred into medium M2 + 3% sucrose with 0.2% AC and 1% agar in culture tubes for further multiplication. The detailed protocol is described [38].
In experiments of ten cooled plantlets each, 70% survival followed by 30-50% shoot recovery and subsequent plantlet development were obtained using this protocol for three genotypes [38].
In Vitro Multiplication Rate
Plantlets developed after cryopreservation and cryoprotectant treatment (control) were multiplied to a maximum of 18 plantlets per treatment and genotype, the same number as that of in vitro maintained plantlets, for the measurement of in vitro MR. The plantlets from these treatment variants (cryopreserved, cryoprotectant treated and the in vitro derived plantlets) were maintained in the same maintenance medium and culture condition. MR, defined as number of explants obtained after two months culture divided by number of survived explants, was measured for each of the 18 plantlets. An explant was defined as a single node with one leaf attached obtained from any part of the plantlet excluding the original node [36]. Three consecutive measurements were made for each treatment variant. At the first and the second subcultures, a number of plantlets (5-8) were randomly selected and multiplied to maintain 18 plantlets for the next sub-culture.
Glass House Transplantation and Data Measurement
Cryopreservation and in vitro derived plantlets (nine plantlets each) were randomly selected from eight-weeks old micropropagated plantlets and transplanted to the glass house. Upon transferred, the plantlets were treated (by dipping into the solution) with a fungicide mixture of 0.2% v/v Polyram Combi (Active ingredient Metiram; BASF AG, Ludwigshafen) and 0.2% v/v Ridomil (Syngenta Agro GmbH, Maintal) in the ratio of 1:1. In the second week of April, transplanting was performed into small pots (diameter 9 cm) filled with 1:1:1 ratio of compost, peat substrate and sand, respectively. After two months cultivation in the small pots, the plants were transferred to bigger pots (diameter 22 cm) containing the same components as that in the small pots in the ratio of 3:1:1. Watering of the plant was conducted once a day and continued up to the middle of October, when the leaves turned yellow and dry. No staking and/or additional fertilization were given at the first cycle of growth (using in vitro plantlets) at the second cycle of growth (using underground tubers of the first harvest). However, a single staking of rope was used for the twinning of the stem. During the spring and summer (the second week of March to the second week of October), the lowest temperature of the glass house was 20-22°C, and during the fall and winter (October to March), it was maintained at 8-10°C. Harvest was conducted in the middle of March, and the tubers harvested from each pot, were replanted together after data measurements.
Observation on morphological traits was conducted during the growing periods of the plants, and at harvest time for the first and second cycles of growth. Some important traits of stem, leaf and tuber were determined based on the descriptor list for Dioscorea IPGRI [41] and compared to the description given by Martin FW [42] and Purseglove JW [43]. Colour description (including colour numbers) of some of the traits was based on the Royal Horticultural Society (RHS) Colour Chart. The number of plantlets, survived upon transplanting, was also indicated as percentage of surviving plantlets. Yield parameters measured were number of tubers and tuber harvest weight per plant. Measurement was taken from all surviving plants.
Statistical analysis
Data of the experiments on in vitro MR and yield of glasshouse-grown plants were subjected to ANOVA since they met the assumption of normal distribution and equal variance. One-Way ANOVA was applied to analyse the in vitro MR in each genotype and yield traits followed by the Tukey’s test to determine the differences among treatment variants, when the ANOVA indicated significant difference. Statistical analysis was accomplished with the help of the software SigmaStat Version 2.0, SPSS Inc., Chicago.
Results
In Vitro Multiplication Rate
In vitro MR of the cryopreserved, cryoprotectant treated and in vitro derived plantlets exhibited significant difference in D. bulbifera (p = 0.002) but not in D. polystachya (p = 0.546) and D. cayenensis (p = 0.36) based on the ANOVA. Tukey’s test indicated that MR of cryopreservation and cryoprotectant treatment derived plantlets were significantly higher than that of in vitro derived plantlets of D. bulbifera. No significant difference of MR was observed between cryopreservation and cryoprotectant treatment derived plantlets in this genotype (Figure 1).
Morphological and Yield Characters
One hundred percent (100%) of the cryopreservation- derived plantlets survived the transplantation as opposed to 67% of the in vitro node culture derived plantlets. Plants derived from the two treatment variations, however, further exhibited normal and similar growth and developments.
Observation on the qualitative traits during the first (from the in vitro plantlets) and the second growing cycles (from tubers) indicated no variation between the two treatment variants. The important traits observed are described as follows (some of the traits can be observed in Fig. 2 and 3): Stem is round in shape, without hairs (glabrous) and twinning anti-clockwise (typical for members of section Enantiophyllum). It is dark green in colour (Green group No.137A and B) and branched at the bottom. Spines are present along the stem but mainly concentrated at its lower part and at the axils of plants developed from tuber. Spines productions, however, were less on the plants developed from the in vitro transplanted plantlets. Leaf is single, broadly ovate in shape with acute apex. Old leaves are dark green in colour (No. 137 A) and the younger ones are yellow green (No. 146 A and B), 7 veins appeared on the lamina. Leaf position is alternate and opposite, and the petiole has wide wings at the bottom. Tuber is cylindrical in shape, has a smooth surface with brown or yellowish skin colour. Tuber sprouting started at harvest time and 78% of cryopreservation derived and 83 % of the in vitro derived plants sprouted after 7 weeks (short dormancy period).
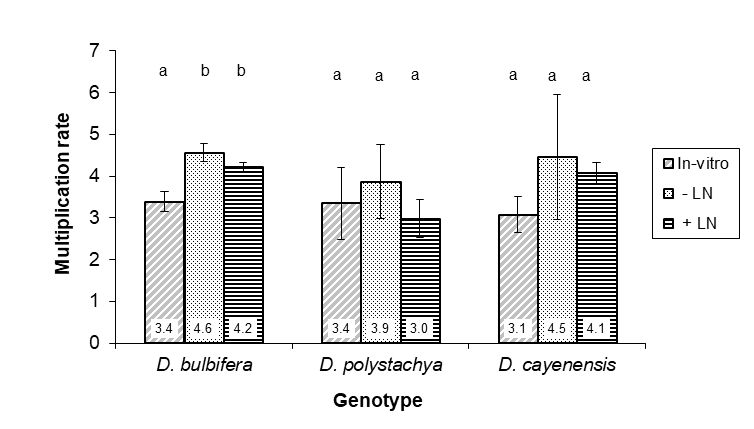
Figure 1: In vitro multiplication rate of cryopreservation, cryoprotectant (PVS2) treatment and in vitro culture derived yams (Dioscorea spp.) of 3 genotypes (species) after measurement of 3 replications with 18 plantlets each. Column on each genotype with the similar letter indicate no significant difference at α = 0.05 based on Tukeys' test. Bars indicate standard error.

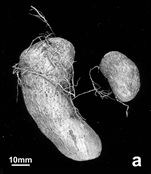
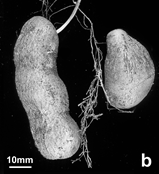
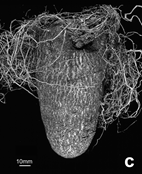
Figure 2: Vegetative development of in vitro and cryopreservation derived plants of Dioscorea cayenensis, 5-6 months after planting. General condition of plants derived from in vitro culture only (left row) and from cryopreserved apical buds at the first cycle (from in vitro plantlets) of growth (a), and spikes formation on the lower part of the stem at the second cycle (from the harvested tubers) of in vitro only (b) and cryopreservation (c) derived plants.


A similar number of tubers (average of two tubers per plant) was produced after the two treatment variants. As well, tuber harvest weight exhibited no obvious difference (Table 1) both at the first (46 vs. 44 g per plant) and the second growth cycles (400 vs. 345 g per plant).
| Source of plants | N | Number of tubers | Ination of weight (g) | |||
|---|---|---|---|---|---|---|
| C1 | C2 | C1 | C2 | C1 | C2 | |
| In vitro | 6 | 5 | 1.8±1.2 | 2.2±1.0 | 45.3±20.8 | 400.4±141.5 |
| Cryopreservation | 9 | 9 | 2.2±1.0 | 2.0±0.9 | 44.9±23.1 | 345.1±164.0 |
Table 1: Number of tubers and tuber harvest weight per plant of pot-grown D. cayenensis derived from in vitro node cultures and c
N = number of replications. C1 first cycle, C2 second cycle The values are followed by the respective standard deviations. Table 1: Number of tubers and tuber harvest weight per plant of pot-grown D. cayenensis derived from in vitro node cultures and cryopreserved apical buds at first and second growth cycles.
Discussion
The effect of cryopreservation using modified droplet (combining droplet and vitrification – 38) method on the physiological and phenotypic characters of yams (Dioscorea spp) at the tissue culture and glass house culture stages was studied.
Cryopreservation might have two consequences on the genetic stability of the germplasm; selection effects especially in heterogeneous populations [16] and additional genetic variability due to various steps implemented in the cryopreservation protocol [14, 22]. Physiological effects causing phenotypic plasticity may also be the basis of variation in the experimental findings. Physiological effects, however, will decrease in further progeny cycles [19]). Results of this study indicated that none of the mechanisms mentioned above has significantly taken place for cryopreserved materials of two genotypes (D. polystachya and D. cayenensis) in terms of in vitro MR as well as all investigated phenotypic qualitative and yield characters for D. cayenensis. Evaluation in further generations might determine whether the alteration in the MR in this study (cases of cryopreservation and PVS2 treatment derived plantlets in D. bulbifera) was due to genetic or physiological causes. However, based on our two years study covering ten consecutive subculture frequencies of MR [44], where the fluctuation exists in every cycle of subculture, it may be assumed that the difference in the MR on D. bulbifera genetic back ground in this study was of physiological cause and will be recovered in further cycles of subculture.
Investigation on the stability of MR of in vitro shoot or node cultures after cryopreservation has not been published for yams. Studies on Panax ginseng, however, indicated that roots regenerated from cryopreserved hairy roots, proliferated fewer new lateral roots compared to the control but exhibited no differences in further development [20]. The suppression of proliferation rate was attributed to direct effect arising from cryogenic procedure [20]. Morphological observation of the plantlets regenerated from somatic embryo and further microtuber production on Dioscorea bulbifera reported a similarity between cryopreserved and control plantlets [28].
Retainment of plant morphological characters after cryopreservation on the glass house grown plants has been reported for potato [31, 32]. Field studies Ahuja S [29] observing several phenotypic characters on D. floribunda, reported similar qualitative characters and no significant differences on the quantitative characters between cryopreserved and in vitro culture generated plants. Plants regenerated from cryopreserved suspension cells of banana Côte Fx [26] and from cryopreserved calluses of sugarcane Martínez-Montero ME [24]) generally indicated stability in several agronomic and morphological traits. Although there were variations between cryopreserved and control materials on two traits at the first cycle of growth of banana plants, these differences disappeared at the second growth cycle [26]. In vitro derived sugarcane plants regardless of their cryopreservation status produced significantly smaller diameter and shorter plant height in comparison to the macro- propagated plants during the first six months of cultivation. These differences disappeared, however, after one year of field growth [24]. In contrast to those studies, a number of quantitative characters including tuber harvest weight, plant height at the flowering stage, length of petiole and breath of terminal leaflet were changed after cryopreservation compared to the tissue culture control plants of potato cv. ’Golden Wonder’[19]. Biometric indicators were also found to be changed after cryopreservation of strawberry, based on the field trials [21].
Except for variation in methylation status (epigenetic change) of ribosomal RNA and nuclear DNA [22] [45], studies on various species confirmed the stability of DNA after cryopreservation. Those studies covered Solanum tuberosum shoot-tips cryopreserved using encapsulation- dehydration and droplet methods [32, 33, 34], Atropa belladona hairy roots and Papaver somniferum calli cryopreserved using vitrification method [20], Citrus spp vitrified calli [23], shoot tips of Dioscorea floribunda [29] and embryogenic tissues of D. bulbifera implementing encapsulation- dehydration method [28]. Further observations have also indicated stability in both chromosome number and ploidy level for potato [30, 31], Dioscorea deltoidea [46] and Citrus spp. Hao YJ [23]. At the biochemical level, consistency on the secondary metabolites productions after cryopreservation has been confirmed for several medicinal species such as P_anax ginseng_, Papaver somniferum Yoshimatsu K [20] and Dioscorea deltoidae [46]. For the species Atropa belladona, the secondary metabolite production was fluctuated at the initial culture after cryopreservation but recovered to the same level of untreated roots after further subculture Yoshimatsu K [20].
The practical impact of stability in the MR is that cryopreserved yam germplasm, upon retrieval from cryopreservation, can be directly propagated for the purpose of rapid plantlet multiplication or for germplam exchange with the same number of plantlets as that normally obtained with the in-vitro multiplication. Furthermore, in the development of cultural practices of yams’ breeding and field production, in vitro plantlets of two or more months old, will be transferred to the field or nursery beds in the first year and the tubers produced are selected based on their characteristics and used for the normal production in the next year [47, 48]. The similarity in tuber production of plants derived from cryopreserved apical buds and in vitro plantlets of D. cayenensis in this study indicates that the similar cultural and selection practices for in vitro plantlets can be applied directly to plantlets recovered from cryopreservation. The number of tubers per plant produced for D. cayenensis in the first cycle of growth (45 g), despite the application of only simple cultural practices (without staking and additional fertilizers), was in the range of that produced in nursery beds (20-250 g) reported for D. rotundata, D. dumetorum and D. bulbifera [48].
Conclusions
Cryopreservation is regarded to be the most suitable method for conservation of seedless, semi-recalcitrant and recalcitrant seeds plants. This method, however, involves different steps which could contribute to the alteration of genetic constitution of preserved germplasm especially when the protocol used is not well developed. The present study indicated no alteration in the i_n vitro_ MR for two of three genotypes tested, and constant morphological characters, number of tubers per plant and tuber harvest weight per plant of glasshouse pots-grown plants of D. cayenensis after cryopreservation. These findings support the concept that cryopreservation, specifically the modified droplet protocol developed for yams, is reliable for the preservation of the genetic constitution of stored yams’ germplasm.
Acknowledgements: The authors thank Mr. J. Marlow and the crew for the technical assistances in handling the plants in the glass house. Mrs. Doris Büchner provided technical assistances in the tissue culture works. German Academic Exchange Service (DAAD) has supported the research work of the first author.
References
-
Malaurie M, Pungu O, Dumont R and Trouslot M-F (1993) The creation of an _in vitro_ germplasm collection of yam (_Dioscorea_ spp.) for genetic resources preservation. Euphytica 65, 113-122.
-
Nair NG and Chandrababu S (1994) A slow growth medium for _in vitro_ conservation of edible yams. J Root Crops 20, 68-69.
-
Ng SYC and Ng NQ (1996) Yam _in vitro_ genebank management at IITA. _In_: Management of field and _in vitro_ germplasm collections. Ed.: Engelman F. IPGRI-FAO, Cali, Columbia. pp. 124-126.
-
Ng SYC and NQ Ng (1997) Germplasm conservation in food yams (_Dioscorea_ spp.): constrain, application and future prospects. _In_: Conservation of plant genetic resources _in vitro_. Eds.: Razdan MK and EC Cocking. Vol. 1. Science Publishers, Inc. USA. pp. 257-286.
-
Taylor M (1996) _In vitro_ Conservation of Root and Tuber Crop in the South Pacific. _In_: Management of field and _in_ _vitro_ germplasm collections. Ed.: Engelman F, IPGRI-FAO, Cali, Columbia. pp. 93-95.
-
Acheampong E (1996) _In vitro_ genebank management of clonally propagated crops under minimal conditions. _In_: Management of field and _in vitro_ germplasm collections. Ed.: Engelman F, IPGRI-FAO, Cali, Columbia, P. 75-75.
-
Ng NQ and SYC Ng (1996) Yam field genebank management at IITA. _In_: Management of field and _in vitro_ germplasm collections. Ed.: Engelman F, IPGRI-FAO, Cali, Columbia. pp.16-18.
-
Taylor M (1996) _In vitro_ Conservation of Root and Tuber Crop in the South Pacific. _In_: Management of field and _in_ _vitro_ germplasm collections. Ed.: Engelman F, IPGRI-FAO, Cali, Columbia. pp. 93-95.
-
Engelmann F (2000) Importance of cryopreservation for the conservation of plant genetic resources. _In_: Cryopreservation of tropical plant germplasm, current research progress and application. Eds.: Engelman F. and Takagi H.. JIRCAS International Agriculture Series. No.8. IPGRI, Rome. ISBN 92-9043-428-7. P. 8-20.
-
Taylor M and T Murikami (2000) Current status and cryopreservation research and future perspectives of its application in the South Pacific. _In_: Cryopreservation of tropical plant germplasm, current research progress and application. Eds.: Engelman F and Takagi H, JIRCAS International Agriculture Series. No.8. IPGRI, Rome. ISBN 92-9043-428-7. p. 330-133.
-
Larkin PJ and Scowcroft WR (1981) Somaclonal variation - a novel source of variability from cell culture for plant improvement. Theor Appl Genet 60: 197-214.
-
Scowcroft WR (1984) Genetic variability in tissue culture: Inpact on germplasm conservation and utilization. IBPGR, Rome, P. 41.
-
Dodds JH, Huaman Z and Lizarraga R (1991) Potato germplasm concervation. _In_: _In vitro_ methods for conservation of plant genetic resources. Ed.: Dodds JH Chapman and Hall. University press, Cambridge, P. 93- 109.
-
Harding K (1991) Molecular stability of the ribosomal RNA genes in _Solanum tuberosum_ plants recovered from slow growth and cryopreservation. Euphytica 55: 141- 146.
-
Harding K (1994) The methylation status of DNA derived from potato plants recovered from slow growth. Plant Cell, Tissue and Organ Culture 37: 31-38.
-
Kartha KK, Engelmann F (1994) Cryopreservation and germplasm storage. _In_: _In vitro_ methods for conservation of plant genetic resources. Ed.: Dodds JH, chapman and Hall, Cambridge, P. 195-230.
-
Finkle BJ, Zavala ME and Ulrich JM (1985) Cryoprotective compounds in the viable freezing of plant tissues. _In_: Cryopreservation of plant cells and organs. Ed.: Kartha KK, CRC press inc, Boca Raton, Florida. P. 75-111
-
Potter RH, Jones MGK (1991) Molecular analysis of genetic stability. _In_: _In vitro_ methods for cryopreservation of plant genetic resources. Eds. Dodds JH, Chapman Hall, London, ISBN 0 412 33870 X, p. 71-9.
-
Harding K and Staines H (2001) Biometric analysis of phenotypic characters of potato shoot-tips recovered from tissue culture, dimethyl sulphoxide treatment and cryopreservation. CryoLetters 22: 255-262.
-
Yoshimatsu K, Touno K and Shimomura K (2000) Cryopreservation of medicinal plant resources: retention of biosynthtetic capabilities in transformed cultures. In: Engelmann F, Takagi H (Eds) Cryopresevation of tropical plant germplasm, current progress and application, IPGRI, Rome, pp 77-88.
-
Vysotskii VA, Karpova OV, Vysotkaya ON (2002) Stability of main properties of berry fruit plant regeneratns after cryopreservation. Sadovodstvo i Vinogradarstvo 3: 9-12.
-
Hao YJ, You CX and Deng XX (2002b) Analysis of ploidy and patterns of amplified fragment length plolymorphsm and methylation sensitive amplified polymorphism in strawberry plants recovered from cryopreservation Cryo Letters 23(1): 37-46.
-
Hao YJ, You CX, Deng XX (2002) Effects of cryopreservation on developmental competency, cytological and molecular stability of citrus callus. Cryo Letters 23(1): 27-35.
-
Martínez-Montero ME, Ojeda E, Espinosa A, Sanches M, Cástillo R, et al. (2002) Field performance of sugarcane (Sacharum sp.) plants derived from cryopreserved caluses. Cryo Letters 23(1): 21-26.
-
Zhai Z, Wu Y, Engelmann F, Chen R, Zhao YH (2003) Genetic stability assessments of plantlets regenerated from cryopreserved in vitro cultured grape and kiwi shoot-tips using RAPD _Cryo Letters_ 24(5): 315-322.
-
Côte Fx, Goue O, Domergue R, Panis B and Jenny C (2000) In-field behaviour of banana plants (_Musa_ aa sp.) obtained after regeneration of cryopreserved embrogenic cell suspensions. Cryoletters **21**(1): 19-24.
-
Butenko RG, Popov AS, Volkova LA, Chernyak ND and Nosov AM (1984) Recovery of cell cultures and their biosythetic capacity after storage of _Dioscorea deltoidae_ and _Panax ginseng_ cells in liquid nitrogen. Plant Sci. Lett 33: 285-292.
-
Dixit S, Mandal BB, Ahuja S and Srivastava PS (2003) Genetic stability assessment of plants regenerated from cryopreserved embryogenic tissues of _Dioscorea_ _bulbifera_ L. using RAPD, biochemical and morphological analysis. CryoLetters 24(2): 77-84.
-
Ahuja S, Mandal BB, Dixit S and Srivastava PS (2002) Molecular, phenotypic and biosythetic stability in _Dioscorea floribunda_ plants derived from cryopreserved shoot tips. Plant Science 163(5): 971-977.
-
Ward ACW, Benson EE, Blackhall NW, Cooperbland S, Powell W, et al. (1993) Flow-cytometric assessments of ploidy satbility in cryopreserved dihaploid solanum- tuberosum and wild solanum species. CryoLetter 14(3): 145-152.
-
Benson EE, Wilkinson M, Todd A, Ekuere U and Lyon J (1996) Developmental competence and ploidy stability in plants regenerated from cryopreserved potato shoot- tips. CryoLetters 17(2): 119-128.
-
Schäfer-Menuhr A, Schumacher H-M and Mix-Wagner G (1997) Long-term storage of old potato varieties by cryopreservation of shoot-tips in liquid nitrogen. Plant Genetic Resources Newsl 11: 19-24.
-
Harding K and Benson EE (2000) Analysis of nuclear and chloroplast DNA in plants regenerated from cryopreserved shoot-tips of potato. CryoLetters 21: 279- 288.
-
Harding K and Benson EE (2001) The use of microsatellite analysis in _Solanum tuberosum_ L. _in vitro_ plantlets derived from cryopreserved germplasm. Cryo Letters 22(3): 199-208.
-
Harding K (2004) Genetic integrity of cryopreserved plant cells: A review. Cryo Letters 25(1): 3-22.
-
Leunufna S and Keller ERJ (2000) Establishement of yams (_Dioscorea_ spp.) in vitro cultures; an initial step of preserving their genetic diversity. Proceeding (CD Room) of “Deutscher Tropentag”, International Conference on Agriculture Research and Development. Bonn, 8p.
-
Leunufna S and Keller ERJ (2003) Investigating a new cryopreservation protocol for yams (_Dioscorea_ spp.). Plant Cell Rep 21(12): 1159-1166.
-
Leunufna S and Keller ERJ (2005) Cryopreservation of yams using vitrifica- tion modified by including droplet method: Effects of cold acclimation and su- crose. CryoLetters 26(2): 93-102.
-
Murashige T and Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15(3): 473-497.
-
Sakai A, Kobayashi S and Oiyama I (1990) Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasilienses Tanaka) by vitrification. Plant cell report 9(1): 30-33.
-
IPGRI (1997) Descriptors for Yam (_Dioscorea_ spp.) IPGRI, Rome, ISBN 92-9043-353-1, pp: 61.
-
Martin FW, Sadik S (1977) Tropical yams and their potential, Part 4. Dioscorea rotundata and Dioscorea cayenensis. USDA, Washington DC, pp: 40.
-
Purseglove JW (1972) Tropical monocotiledone, Dioscoreaceae. Longman Goup Ltd., London, ISBN 0582 466482, pp. 97-117.
-
Leunufna S (2004) Improvement of the in-vitro maintenance and cryopreservation of yams (Dioscorea sp.). Dissertation, Verlag Dr. Köster, Berlin, pp: 130.
-
Harding K (1997) Stability of the ribosomal RNA genes in _Solanum tuberosum_ L plants recovered from cryopreservation. Cryo Letters 18(4): 217-230.
-
Popov AS, Volkova LA, Butenko RG (1995) Cryopreservation of germplasm of _Dioscorea deltoidae_ (medicinal yam). _In_: Biotechnology in agriculture and forest, cryopreservation of plant germplasm 1 (32). Ed.: BAJAJ YPS. Sringer-Verlag, Heidelberg, Berlin, ISBN 3-540-57451-4, pp: 488-499.
-
Wilson JE (1982) Progress in the breeding of yams, Dioscorea spp. _In_: Yams, Ignames (Eds.), MIEGE J, LYONGA SN. Oxford University Press, New York, ISBN 0-19-854557-6, pp.17-22.
-
Coursey DG and Degras L (1986) Yams (_Dioscorea_ spp.) In: Leon J, Withers LA (Edr.), Guidelines for seed exchange and plant introduction in tropical crops. FAO, Rome, 59-64.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note