Morpho-Physiological and Biochemical Responses of Plectranthus amboinicus to Varying Watering Regimes
Water stress is a detrimental event that adversely impacts crop productivity on a global scale. A study was carried out to examine the impact of fluctuating water stress conditions on Jamaican thyme (Plectranthus amboinicus) plant morpho-physiological and biochemical indices. The treatments were regular watering (RW), drought (DR), flooding (FL), and resumption of regular watering after flooding (DHFL) or after drought (RHDR). The DR and FL treatments significantly (p<0.01) reduced plant fresh weight by 92% and 88%, respectively, compared to RW. The relative water content of DR plants showed a significant (p<0.01) reduction by 60% compared to the RW plants. Net photosynthetic rate, transpiration rate, and stomatal conductance were significantly (p<0.01) reduced in DR and FL plants; whereas sub-stomatal carbon dioxide was significantly (p<0.01) high in DR plants. Chlorophyll a, b, and carotenoids were significantly (p<0.05) reduced in the DR and FL plants compared to the RW plants. On the other hand, soluble sugar content in the DHFL plants was significantly (p<0.05) increased compared to that of the RW plants. FL plants showed a significant (p<0.05) increase in total 2,2-diphenyl-1-picrylhydrazyl scavenging effect by 156%, total flavonoids by 225%, total phenolics by 242%, and malondialdehyde by 94% compared to RW. In conclusion, this study highlighted the morpho-physiological and biochemical changes in Jamaican thyme plants under varied watering regimes. Jamaican thyme is an aroma medicinal plant and thus, further research should assess volatile organic compounds.
Introduction
The shift in climatic patterns presents a significant challenge to the survival of life on earth. This can be partly attributed to the increasing difficulty of meeting the escalating demand for food and establishing sustainable agricultural practices to support a growing population [1]. The current state of changing climatic conditions, characterized by unexpected occurrences such as droughts, severe floods, earthquakes, and fluctuations in temperature destabilizes the hydrologic cycle [2]. In the last decade, there has been a significant increase in the expanse of arid regions worldwide. This expansion has been accompanied by a noticeable rise in the prevalence, severity, and frequency of drought and flooding events. Consequently, the cumulative impact of these droughts has led to an estimated global economic loss of almost $30 billion in agricultural production [3, 4]. Flooding is the second most significant climate-related calamity following drought. The number, frequency, and severity of flood disasters have risen from the 1990s [5, 6]. Plants exhibit adaptive responses to water stress (flooding, drought) through alterations in various plant morphological structures and biochemical processes. Such adaptation mechanisms include modifications in leaf and root morphology, adjustments in photosynthetic activity, and changes in antioxidant enzyme systems, and fluctuations in hormone levels [7, 8]. Currently, it is well acknowledged that drought stress primarily impacts the assimilation and translocation of nutrients from the root system to the foliage [9, 10].
Plants generate excessive amounts of reactive oxygen species (ROS) such as singlet oxygen, superoxide, and hydrogen peroxide in response to water stress. ROS exhibit a high reactivity and possess the ability to swiftly inflict damage on living tissues and macromolecules such as deoxyribonucleic acids, lipids, proteins, and carbohydrates [11]. Consequently, this might trigger the initiation of programmed cell death [11]. Sugars such as glucose, fructose, sucrose and sugar alcohols like mannitol, and amino acids such as proline accumulate in many plant species when exposed to water stress. These compounds serve as osmolytes to aid in the regulation of osmotic balance and function as antioxidants. Their presence helps in the detoxification of ROS, protection of cellular membranes, and stabilization of enzymes and proteins. Ultimately, these compounds contribute to enhancing plant resilience against abiotic stress [12, 13, 14]. The influence of water stress on plants is contingent upon various factors including the extent and length of water supply, soil quality, soil water gradients, the plant species, and their developmental stages [15]. Plants are sessile and have developed numerous adaption mechanisms that may improve their ability to thrive under both short- and long-term water stress conditions [16, 17].
In this study, we used Jamaican thyme (Plectranthus amboinicus) belonging to the Lamiaceae family with a diverse array of ethnobotanical benefits [18]. Jamaican thyme is a succulent Crassulacean acid metabolism plant whose growth and key metabolic pathways in the central carbon metabolism can be severely affected by different watering regimes [19]. However, it is not clear how varying watering regimes affect plant growth, photosynthesis rate, and stress response metabolites of Jamaican thyme. This knowledge can be utilized to enhance stress tolerance and ultimately enhance crop productivity [20]. The present study aims to examine the growth, chlorophyll fluorescence indices, and biochemical composition of Jamaican thyme plants under different watering regimes.
Materials and Methods
Plant propagation and treatment application were modified from Abbey, et al. [19].
Location
The research was performed in the Plant Physiology Laboratory of the Department of Plant, Food, and Environmental Sciences, Faculty of Agriculture between September 2022 and June 2023.
Preparation and Rooting of Cutting
A healthy mother plant was chosen from the PFES greenhouse plant stock. The plant was well watered to relieve it from stress conditions. Stem cuttings were taken from branches from the youngest second and third nodes of soft tissues on the main stem. The stem cuttings were then pruned to 5-cm length leaving five pairs of corresponding leaves before placing in a moist perlite medium (Perlite Canada Inc., Montreal, QC, Canada) contained in a plastic flat tray (50 length x 28 width x 6.5 depth). The planted trays were covered with a dome-shaped transparent cover to maintain ≥95% relative humidity for rooting, 24-hour fluorescent lighting at a temperature of 22°C. The cuttings were sprayed with water twice a day until transplanting after three weeks.
Planting of Rooted Cutting and Growing Condition
The Jamaican thyme cuttings were transplanted into 15-cm diameter plastic pots filled with a mixture of 200 g of Promix-BX (Premier Tech Horticulture in Québec, Canada) and 150 g of vermicast. The cuttings were placed in a Biotranette Mark II Environmental growing chamber (Lab- Line Instruments Inc., ILL, USA) in a completely randomized design with four replications under 24°/20°C and a 12/12- hour day and night cycle. The plants were watered regularly for 3 months.
Water Stress Treatment and Experimental Design
The experimental treatments used were previously reported by Abbey, et al. [19]. In brief, it consisted of regular watering (RW), extended drought (DR), extended flooding (FL), rehydration following extended drought (RHDR), and dehydration following extended flooding (DHFL). For RW, the plants were watered regularly on alternate days to maintain the optimal moisture level of the growing medium. For DR, the plants were not watered after the 3 months of regular watering, and for FL, the potted plants were placed in a larger pot of 20 cm diameter and filled with excess water. The treatments for both the DR and FL plants were administered for 60 days. The stress reversals were the resumption of regular watering following a continuous flooding period of 30 days (DHFL) or a continuous drought period of 30 days (RHDR). The experiment was designed using a completely randomized design with four replications.
Plant Growth and Physiological Analysis
Total Fresh Weight: The total fresh weights of the plants were recorded using an Ohaus navigator portable balance (TTM Instruments Inc., Canada) at the end of the experiment i.e., 60 days after transplanting.
Chlorophyll Fluorescence and Photosynthetic Parameters: Water stress traits were assessed using a Chlorophyll fluorometer (Optical Science, Hudson, NH, USA) to obtain maximum quantum efficiency (Fv/Fm) and potential photosynthetic capacity (Fv/Fo). Other photosynthetic parameters i.e., sub-stomatal carbon dioxide concentration, net photosynthetic rate, and transpiration rate were determined from two pairs of healthy opposite leaves (n = 4) from each plant per treatment using LCI portable photosynthesis system (ADC Bio Scientific Ltd., Hoddesdon, UK).
Electrolyte Leakage and Leaf Relative Water Content: Electrolyte leakages from disrupted cells were estimated by the electric conductivity using an EC 500 Ex-Stik multipurpose pH meter (EXTECH Instrument, Nashua, NH, USA). 1-cm diameter leaf discs were submerged in 20-mL deionized water and the electric conductivity was measured every 2 h for 12 h under room temperature and light conditions. Leaf relative water content was estimated by measuring the fresh, dry, and turgid weight of 1-cm diameter leaf discs with MXX-412 Denver precision electronic balance (Denver Instrument Company, CO, USA) from four opposite leaves in triplicate (n = 12). The turgid weight was determined by submerging the fresh leaf discs in 100 ml of deionized water for 48 h and weighed. The leaf discs were then dried in a Cole-Parmer mechanical convection oven dryer (Cole-Parmer Instrumental Company, Vernon Hills, IL, UK) at 65°C to constant weight for 48 h and weighed. The relative water content was calculated using the formula:
$$\text{Relative Water Content} \left( \frac{\text{RWC}}{\text{RWC}} \right) = \frac{\text{Fresh weight} - \text{Dry Weight}}{\text{Turgid Weight} - \text{Dry Weight}} \times 100$$
Biochemical Analysis: Healthy leaf samples were flash-frozen in liquid nitrogen. The samples were ground into fine powder and stored in a -80°C freezer until analysis.
Chlorophylls a and b, and Carotenoid Contents: Chlorophylls (Chls) a and b, and carotenoids were measured as described by Lichtenthaler HK, et al. [21] with little modification. Ground leaf samples (0.2 g) were thoroughly mixed in 1 mL of 80% acetone followed by centrifugation at 15000 × g for 15 min. The absorbance of the supernatant was recorded at 646.8 nm, 663.2 nm, and 470 nm using a UV-Vis spectrophotometer (Jenway, Staffodshire, UK) against 80% acetone as a blank. The Chls and carotenoid contents were expressed as mg g$^{-1}$ fresh weight (FW) of the sample.
Total Soluble Sugar (TSS): Total soluble sugar was determined using the method of DuBois M, et al. [22]. Briefly, a 0.2 g of the ground leaf sample was homogenized in 1.5 mL of ice-cold freshly prepared 5% trichloroacetic acid (TCA). The mixture was vortexed for 2 min and centrifuged at 12000 × g for 3 min. An aliquot of 1 mL was transferred into a thick-walled glass test tube containing 1 mL of 5% phenol and mixed thoroughly. The reaction mixture was mixed with 5 mL of concentrated sulfuric acid, vortexed for 20 s, and incubated in the dark for 15 min. After the mixture had reached room temperature, the absorbance was measured at 490 nm against a blank. Total sugar was calculated using a glucose standard sugar curve and expressed as $\mu$g of glucose g$^{-1}$ FW of the sample.
Total Phenolics: Total phenolic content (TPC) was determined using the Folin-Ciocalteau assay as described by Ainsworth EA, et al. [23] with little modification. A 0.2 g of the ground leaf sample was homogenized in 2 mL of ice-cold 95% methanol and incubated in the dark at room temperature for 48 h. The mixture was centrifuged at 13000 × g for 5 min before adding 100 mL of the supernatant to 200 μL of the 10% (v/v) Folin-Ciocalteau reagent. After vertexing for 5 min, the mixture was combined with 800 μL of 700 mM sodium carbonate (Na$_2$CO$_3$) and incubated in the dark at 25°C for 2 h. The absorbance of the supernatant was measured at 765 nm against a blank. TPC was calculated using a gallic acid standard curve and expressed as mg of gallic acid equivalents g-1 FW of the sample.
Total Flavonoid: Total flavonoid was measured following the colorimetric method described by Chang CC, et al. [24]. A 0.2 g ground leaf sample was homogenized in 2.5 mL of 95% methanol followed by centrifugation at 15000 × g for 10 min. A reaction mixture was obtained by adding 1.5 mL of 95% methanol, 0.1 mL of 10% aluminum chloride (AlCl$_3$), 0.1 mL of 1 M potassium acetate, and 2.8 mL of distilled water to 500 μL of the supernatant. The reaction mixture was incubated at room temperature for 30 min and the absorbance was measured at 415 nm against a blank lacking AlCl$_3$. Total flavonoid content was calculated using the quercetin equivalents and expressed as $\mu$g of quercetin g$^{-1}$ FW using the formula below:
$$\text{Total flavonoid} = \frac{\left( \text{Flavonoids} \right) \left( \frac{\mu \text{g}}{\text{mL}} \right) \times \text{total volume of methanolic extract}(\text{mL})}{\text{mass of extract}(\text{g})}$$
Lipid Peroxidation Concentration: The concentration of malondialdehyde (MDA) was used to determine the extent of lipid peroxidation following the method described by Hodges DM, et al. [25]. A 0.2 g of the ground leaf sample was homogenized at 1 mL of 0.1% (w/v) trichloroacetic acid (TCA) followed by centrifugation at 17000 × g and 4°C for 10 min. 500 mL of the supernatant was mixed with equal volume of 0.5% thiobarbituric acid (TBA) in 20% TCA. The mixture was vortexed for 30 s, incubated at 95°C for 30 min and centrifuged at 10,000 × g for 5 min. The absorbance of the reaction mixture was measured at 400 nm, 532 nm, and 600 nm against the blank of TBA solution. The MDA equivalent was expressed as nmol g-1 FW using the formula:
$$ M D A c o n c e n t r a t i o n = \left[ \frac {A b s 5 3 5 - A b s 6 0 0}{1 5 5 0 0 0} \right] \times 1 0 ^ {6} $$ DPPH Free Radical Scavenging Capacity: DPPH radical scavenging capacity was measured using the method described by Fan D, et al. [26]. A 1.5 mL pure methanol was added to 0.2 g of the ground leaf sample and mixed thoroughly. The mixture was centrifuged at 12000 × g for 10 min and 100 µL of the supernatant was added to 2.9 mL of 60 µM fresh DPPH methanolic solution. The mixture was incubated in the dark at 22°C for 30 min and the absorbance was measured at 515 nm against a methanol blank. The following formula was used to calculate the scavenging capacity:
$$ Inhibition \% = \left[ \frac {A b - A s}{A b} \right] \times 100 \% $$
As the absorption of treatments and Ab being the absorption of control samples.
Hydrogen Peroxide Accumulation: Hydrogen peroxide (H2O2) was determined according to the method described by Patterson BD, et al. [27]. A 0.2 g of the ground leaf sample was homogenized in 2 mL of cold acetone and the mixture was centrifuged at 10,000 × g for 10 min. A 0.4 mL Titanium (II) chloride and 0.2 mL of 17 M ammonia solution were added to 1 mL of the supernatant. The precipitate was washed five times with acetone by resuspension and dissolved in 2 mL of 1 M Sulfuric acid. The absorbance of the resultant mixture was measured at 410 nm against a blank. The H2O2 concentration was calculated according to a standard curve and expressed as mM g-1 FW.
Statistical Analysis
Statistical analysis was performed using Minitab version 21. The values were expressed as a means of four replicates with standard deviations (SD). All the data were subjected to one-way analysis of variance (ANOVA). Tukey’s honestly significant difference post-test was used to separate the means at 5% significance level.
Results and Discussion
Morpho-Physiological Parameters
The results indicated that the different levels of water stress conditions significantly (p<0.01) affected plant growth (Figure 1). The plants subjected to severe drought (DR) and flooding (FL) conditions exhibited a substantial decrease of 92% and 86% in total plant fresh weight, respectively, compared to the control (RW) plants (Figure 1). Similarly, a previous study revealed that severe water stress showed a significant (p<0.01) reduction in the total fresh and dry mass accumulation in Satureja hortensis L (Cimbru Savory Summer) Baher ZF, et al. [28]; DHFL and RHDR exhibited a significant (p<0.01) reduction in fresh weight by 43% and 33%, respectively, compared to that of the RW plants.
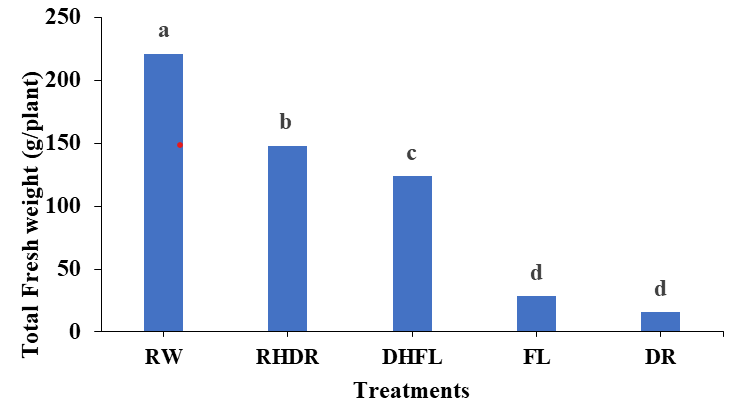
Figure 1: Total plant fresh weight as affected by varying watering regimes; Regular watering (RW), drought (DR), resumption of regular watering after drought (RHDR), flooding (FL), and resumption of regular watering after flooding (DHFL). Different alphabetical letters on the bars denote significant differences according to Tukey’s honestly significant difference post-test analyses at p ≤ 0.05.
Reactive oxygen species are elevated under situations of water stress, which leads to the disruption of the electron transport system in plants [29]. This disruption results in oxidative activity occurring in both the chloroplasts and mitochondria, ultimately leading to a decrease in plant fresh weight [29]. On the other hand, the lack of sufficient soil water leads to decreased nutrient uptake and distribution. As a result, the roots cannot absorb nutrients effectively and transport them to the shoot [30].
Leaf relative water content (RWC) represents the equilibrium between the amount of water supplied to the leaf tissue and the rate at which transpiration loses water [31]. In this study, the RWCs of the DR and FL plants dropped by 6% and 9%, respectively, compared to that of the RW plant (Figure 2).
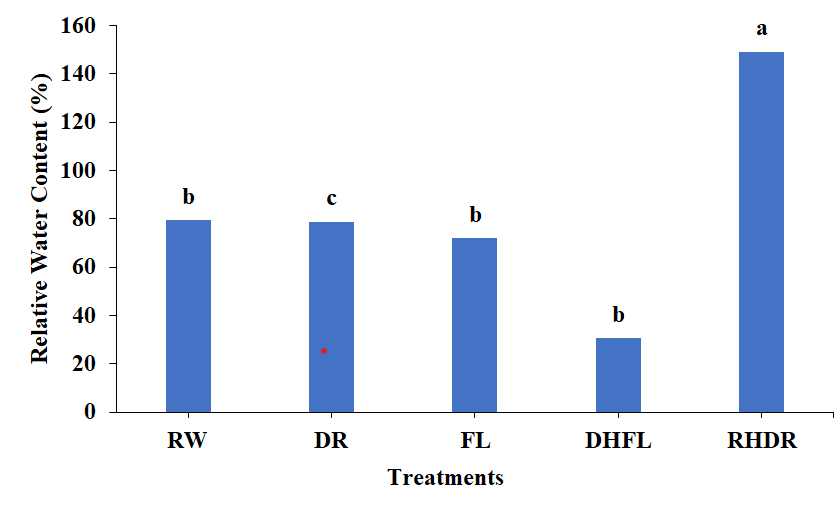
Figure 2: Relative Water Content (RWC) of the samples under various water stress conditions. The treatments include regular watering (RW), drought (DR), and resumption of regular watering after drought (RHDR), flooding (FL), and resumption of regular watering after flooding (DHFL). Different alphabetical letters on the bars denote significant differences according to Tukey’s honestly significant difference post-test analyses at a significant level of p < 0.05.
Moreover, the DHFL plants exhibited a high reduction of 61% in RWC compared to the RW plants while the RHDR plants exhibited a substantial increase of 88% compared to the RW plants (Figure 2).
The application of water stress has a considerable impact on the physiology of plants and their water interactions [32]. The results of this experiment demonstrated that plants exhibited varying levels of leaf-relative water content (LRWC) in response to varied levels of water stress, indicating the impact of water stress on LRWC. The outcome of this study validated the conclusions of Chen S, et al. [33], El Jaafari, et al. [34] and Boutraa T, et al. [35]. Who claimed that the leaf- relative water content declined as a result of increased water stress levels and enhanced resistance to water flow in stems and leaves [19]. A proficient xylem transport system that boosts plant capacity to absorb water could assist the plant in maintaining open stomata and high relative water content when soil water is reduced [36].
Electrolyte Leakage
Electrolyte leakage is a characteristic feature of stress response in intact plant cells [37]. This phenomenon is commonly employed to assess the damage caused by stress on plant tissues and as an indicator of a plant’s ability to tolerate stress [38]. In both the DR and the FL plants, there were substantial reductions in cell electrolyte leakage compared to the RW plants (Figure 3). However, the stress reversal treatments, DHFL and RHDR, had significant (p<0.05) increases in electrolyte cell leakage compared to the RW plants (Figure 3). In contrast, a study conducted by Babaei K, et al. [39] revealed that drought stress significantly (p<0.05) increases electrolyte leakage in plants.
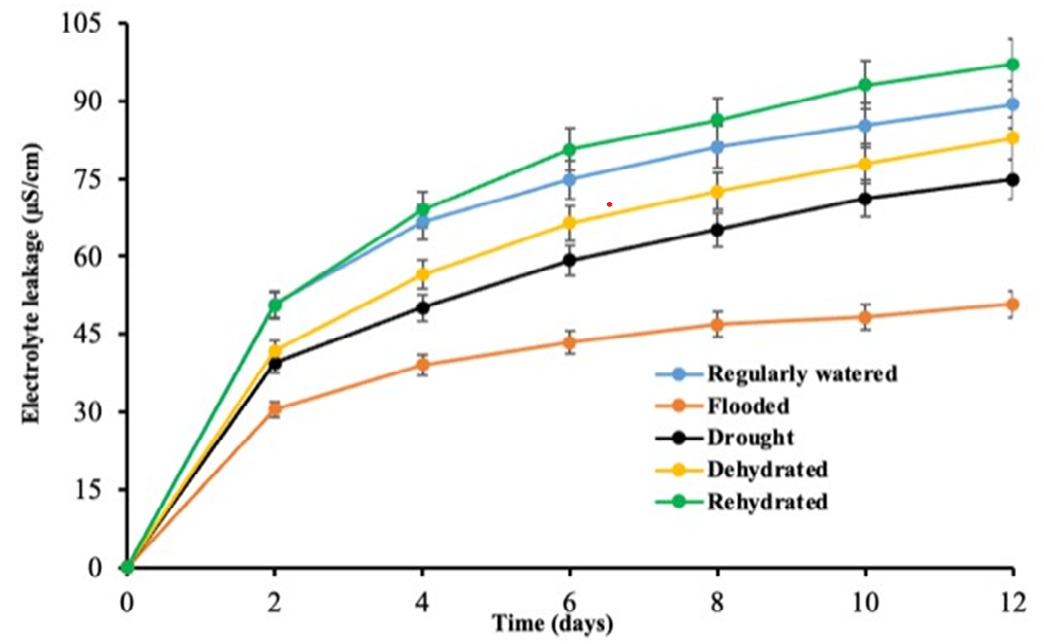
Generally, under extreme water stress conditions, plants typically experience cellular damage resulting in a high level of electrolyte leakage Demidchik V, et al. [37] which can explain the results of the present study. Nevertheless, the decrease in electrolyte leakage during severe water stress conditions as observed in FL followed by DR may be attributed to the capacity of the plants to withstand stressful conditions. This finding is consistent with research conducted by Leopold AC, et al. [40] contrastingly, severe water stress led to a substantial increase in electrolyte leakage (EL) by 127% and 183% in Camarosa and Gaviota, respectively, compared to normal growth conditions [41]. Furthermore, during periods of intense flooding, plant cells might have a dilution effect due to excessive water intake, which may lead to a decrease in the concentration of electrolytes. Furthermore, under drought conditions, plants primarily respond by closing their stomata, which might potentially restrict the exchange of gases and the absorption of ions Pirasteh-Anosheh H, et al. [42] leading to less concentration of electrolytes. This may be a contributing factor to the decline in electrolyte concentration in the FL and DR plants compared to their counterparts in the other treatments.
Chlorophyll Fluorescence Indices and Photosynthesis
Chlorophyll fluorescence indices are valuable parameters for investigating the impact of various environmental conditions on photosynthesis [43, 44]. Leaf Fv/Fm is essential for assessing the condition and functionality of the internal chlorophyll system during periods of water-induced stress [45, 46]. Our findings indicate that Fv/Fm exhibited a modest decline of 7% and 10% under DR and FL conditions, respectively, as compared to RW plants (Table 1).
| Treatment | Fv/Fm | Fv/Fo | A (μmolm-2 s-1) | Ci (μmol mol -1) | E (mol m-2 s-1) | gs (μmol mol -1) | L* | a* | b* | c* | h* |
|---|---|---|---|---|---|---|---|---|---|---|---|
| RW | 0.81ab | 4.48a | 0.48a | 341.80b | 0.22a | 0.01ab | 43.79ab | 10.89ab | 22.28a | 24.81a | 116.08a |
| FL | 0.73c | 2.68c | 0.30ab | 443.00b | 0.04c | 0.00b | 42.32b | 12.19a | 22.80a | 25.82a | 117.34a |
| DHFL | 0.82a | 4.76a | 0.46a | 357.81b | 0.28a | 0.01a | 42.51b | 12.45a | 23.93a | 26.99a | 117.48a |
| DR | 0.75bc | 3.10bc | 0.16b | 1075.00a | 0.06b | 0.00b | 50.84a | 9.82b | 27.80a | 29.57a | 109.93b |
| RHDR | 0.79ab | 4.12ab | 0.40ab | 380.40b | 0.23a | 0.01ab | 42.32b | 10.76ab | 24.67a | 27.11a | 114.57ab |
| p-value | 0 | 0 | 0.03 | 0 | 0.12 | 0.01 | 0.01 | 0.01 | 0.06 | 0.11 | 0.01 |
Table 1: Chlorophyll fluorescence and photosynthesis response of Jamaican thyme under varying water stress conditions.
Fv/Fm, maximal photochemical efficiency; Fv/Fo, potential photosynthetic capacity; A (net photosynthetic rate), Ci (sub-stomatal CO2 concentration), E (transpiration rate), gs (stomatal conductance), L* (lightness), a* and b* (chromaticity coordinates: -a*: green, +b*: yellow), c* (chroma), h* (hue angle) of Jamaican thyme affected by flooding (FL), drought (DR), dehydration after flooding (DHFL) and rehydration after drought (RHDR). Different alphabetical letters showed a significant difference. Different alphabetical letters denote significant differences according to Tukey’s honestly significant difference post-test analyses at a significant level of p ≤ 0.05.
This suggests that drought stress may cause damage to the photosystem II (PSII) and limit the primary reaction of photosynthesis [47]. Concurrently, the decrease in Fm under stress conditions may be influenced by a decline in the functioning of the water-splitting enzyme complex that may be accompanied by a simultaneous cyclic electron transfer inside or surrounding PSII [48]. Consequently, the Fv/Fm ratio will diminish in response to drought-induced stress. The results of the present study were in line with a prior investigation, which showed that drought stress impeded the photochemical activity of PSII and reduced leaf Fv/Fm [49]. A decrease in Fv/Fm signifies the inhibition of photosynthesis in response to water stress [47, 50]. The photosynthetic electron transport chain is primarily influenced by the Fv/Fo component, which is the most sensitive part of the process [51, 52]. The present study found that Fv/Fo was considerably decreased in DR, FL, and RHDR plants compared to RW and DHFL plants (Table 2). However, the DHFL plants showed a non-significant (p>0.05) rise of 6% in Fv/Fo compared to the RW plants (Table 2).
Photosynthesis is the mechanism that provides the foundation for plant growth Osakabe Y, et al. [53] while stomatal conductance (Gs) represents the extent to which stomata are open. Stomata serve as openings in leaves that regulate the exchange of gases, hence controlling the processes of photosynthesis and transpiration in vegetation. Gs was reduced significantly (p<0.05) with DR and FL by 74% and 100% respectively, compared to the RW plants. Stress reversal treatments, DHFL and RHDR, also showed a slightly significant (p<0.05) reduction in stomatal conductance compared to the RW plants by the transpiration rate reduced significantly (p<0.05) in the DR, FL, and DHFL plants whereas RHDR plants showed a moderate increase in the transpiration rate compared to the RW plants (Table 2). Interestingly, the intercellular carbon dioxide concentration was significantly (p<0.05) increased by all the treatments compared to RW (Table 2). It is well established that water stress in plants is characterized by stomatal closure and restricted gas exchange [54]. Thus, plants often constrict their stomata in response to water stress to minimize water loss via transpiration. This might have restricted the intake of atmospheric CO2, possibly causing a buildup of sub-stomatal CO2 concentration (Ci).
Water stress can impede gas exchange and hence, impact the photosynthetic ability of plants [55]. The present study demonstrated that water stress substantially decreased net photosynthesis rate (A), transpiration €, and stomatal conductance (gs), which agree with the findings of Ma SC, et al. [56] and Liu EK, et al. [57]. Other studies demonstrated that water stress induces either stomatal restriction, non- stomatal restriction, or a combination of both during the process of photosynthesis [58, 59]. The primary limiting factor for photosynthesis in this study was stomatal conductance, as it dropped significantly (p<0.05) under conditions of water stress (Table 2). Stomatal closure has been regarded as an initial plant response to water stress, aimed at regulating water loss [60, 61]. Therefore, plants usually exhibit stomatal closure and reduced photosynthesis as reactions to water stress [62].
Biochemical Parameters Under Various Stress Conditions
Chlorophyll a and b, and carotenoids Jamaican thyme plants exposed to prolonged periods of drought showed a non-significant (p<0.05) decrease in leaf chlorophyll content by 5%, compared to the RW plants (Table 2). At the same time, both prolonged periods of FL and DR showed a significant (p<0.05) reduction in chlorophyll b content by 12% and 25% respectively, compared to the RW plants (Table 2). In a similar study by Enneb H, et al. [63], they reported a notable decrease in chlorophylls a and b contents in three assertions of Vicia faba L. under water stress conditions. These previous study reveals that drought stress leads to a reduction in total chlorophyll, which can lead to a diminished ability to capture light [64].
| Chla (μg g−1 FW) | Chlb (μg g−1 FW) | Car (μg g−1 FW) | Sugar (μg /g FW) | ROS (%) | MDA (nmol g−1 FW) | DPPH (%) | Total Flavonoid (%) | Total Phenolics (mg GAE/g FW) | |
|---|---|---|---|---|---|---|---|---|---|
| RW | 128.91d | 85.52c | 28.94d | 19.59ab | 157.30ab | 1.04c | 6.80b | 245.50c | 212.00b |
| FL | 159.04c | 75.24cd | 52.49b | 25.39ab | 216.05a | 2.03a | 17.47a | 800.20a | 725.38a |
| DHF | 249.53a | 147.87a | 111.45a | 42.92a | 144.50ab | 1.53b | 4.49c | 265.80c | 185.10c |
| DR | 121.26d | 63.61d | 40.32c | 31.32ab | 232.20a | 0.99c | 8.36b | 386.00b | 319.30b |
| RHD | 190.39b | 125.82b | 112.26a | 11.52b | 110.50b | 0.69d | 7.56b | 198.50c | 304.09b |
| p-value | 0 | 0 | 0 | 0.02 | 0.01 | 0 | 0 | 0 | 0 |
Table 2: Biochemical responses of Jamaican thyme under varying water stress conditions.
Chla (chlorophyll a), Chlb (chlorophyll b), Car (carotenoid), Sugar (soluble sugar determination), Total flavonoid, Total phenolics, DPPH (ROS Scavenging effect), MDA (malondialdehyde concentration), ROS (reactive oxygen species determination) of Jamaican thyme affected by flooding (FL), drought (DR), dehydration after flooding (DHFL) and rehydration after drought (RHDR). Different alphabetical letters denote significant differences according to Tukey’s honestly significant difference post-test analyses at a significant level of p < 0.05.
Additionally, water stress slows down the rate of carbon fixation by reducing the amount of CO2 influx into leaves, which directly affects overall plant metabolism [65, 66]. Thus, a reduction in Ribulose bisphosphate carboxylase oxygenase (Rubisco) activity, photosynthetic pigments, and photosynthetic electron transport components may cause a drop in molecular oxygen that may lead to the accumulation of reactive oxygen species (ROS) that can cause oxidative stress and harm the photosynthetic apparatus [66, 67, 68, 69]. RHDR and DHFL showed significant (p<0.05) increases in chlorophylls a and b compared to RW plants (Table 2).
Both DR and FL plants showed a significant (p<0.05) increase in carotenoid by 39% and 81% respectively, compared to RW plants (Table 2). The above findings were also in agreement with a study conducted by Mohammadkhani N, et al. [69] where carotenoid contents were increased in drought-stressed Origanum vulgare L. In RHDR plants, the carotenoid content significantly (p<0.05) decreased by 57% compared to that of the RW plants. Similarly, a study conducted by Deng X, et al. [70] revealed a notable increment in the carotenoid content in Boea hygrometrica following a sudden drop in the initial hours of desiccation and subsequently, returned to its original level upon rehydration. On the contrary, the pigment-protein complexes in the leaves of Chirita heterotrichia were destroyed after being dehydrated for 8 days and did not recover after being rehydrate [70]. The results of the present study indicated that DHFL plants showed a massive increase of 285% in carotenoid content compared to that of the RW plants (Table 2). Carotenoids are believed to have a protective function against photoinhibition.
Xanthophyll is vital components of carotenoids and plays a significant role in safeguarding photosynthesis. Therefore, carotenoids are essential for enhancing the ability of plants to tolerate water stress [70]. It is reasonable to postulate that Jamaican thyme may have acquired an adaptive strategy to preserve membrane integrity and antioxidant defense by elevating the carotenoid content during periods of water stress.
Phenols and Flavonoids
Total phenolics was significantly (p<0.05) affected by differences in watering regime. There was a tremendous increase in the FL plants phenolics compared to that of the RW plants. The DR plants also showed a significant (p<0.05) increase in total phenolics by 43% compared to the RW plants (Table 2). The RHDR plants showed a non-significant (p<0.05) increase in the total phenolic concentrations compared to the RW plants (Table 2). Interestingly, DHFL plants showed a significant (p<0.05) decrease in phenolics compared to that of the RW plants.
Flavonoids, which possess a significant antioxidant capacity, have gained interest due to their favourable impact on human health and their ability to safeguard plants against oxidative damage caused by water stress [71]. In this study, the production of total flavonoids was increased significantly (p<0.05) by 225% in FL plants compared to the RW plants. Also, RHDR plants showed an increase in total flavonoids by 57% compared to that of RW plants. Interestingly the stress reversals i.e., DHFL and RHDR showed a non-significant (p<0.05) alteration in the total flavonoid content (Table 2). The concentration of total flavonoids in DHFL plants exhibited a marginal increase of 8% when compared to RW plants. However, in contrast, plants treated with RHDR had a reduced flavonoid concentration by 19% when compared to the plants treated with RW.
Water stress poses a significant challenge for plants due to the physiological response triggered by limited water availability. Specifically, a chemical signal is conveyed from the roots to the leaves through the xylem, causing partial closure of the stomata. This leads to a substantial reduction in intracellular CO2 levels. Consequently, there is a decrease in the availability of NADPH+ and H+ for the Calvin cycle, leading to a reduction in the concentration of NADP+ and the electron receptor potential for the electron transport chain. This results in the elevation of free radicals and reactive oxygen species (ROS). Hence, phenolic compounds such as tannins and flavonoids are produced as a defence strategy against oxidative stress to protect the photosynthetic machinery, prevent damage to cell membranes, limit protein denaturation, and avoid reduction of plant growth. The differences in the levels of secondary metabolites are influenced by each species in response to water stress. Similarly, conducted a study that demonstrated that the concentration of flavonoids was enhanced during drought circumstances. A study conducted by de Lima VT, et al. [72] in Croton floribundus and Croton urucurana revealed that the antioxidant activity was higher in reaerated plants after continuous flooding. It is crucial to understand that the enhancement of secondary metabolite production is not consistent across all plant tissues and organs. However, it may alter among various parts or stages of development within the same plant species. For instance, in a study conducted by Pirbalouti AG, et al. [73] the amount of phenolic compounds was increased in the shoots of two types of basil (Ocimum basilicum L.) plants when they received less water (30% FC) compared to when they were irrigated normally. In contrast, an investigation conducted by Caser M, et al. [74] demonstrated that leaves of Salvia dolomitica Codd. When exposed to intense water scarcity, showed a notable reduction in the overall concentrations of phenols and flavonoids.
Reactive Oxygen Species
Water stress triggers the buildup of reactive oxygen species (ROS) [75, 76]. High amounts of reactive oxygen species (ROS) are extremely reactive and toxic, causing oxidative stress and molecular and cellular damage [77]. Additionally, they have the potential to induce cellular apoptosis [78]. In the present study, there was a significant increase of 155% in ROS concentration in FL plants compared to RW plants. In DR plants, there was a significant (p<0.05) accumulation of ROS by 22% compared to RW plants. RHDR showed a slightest increase of 11% in ROS compared to RW plants (Table 2). In contrast, the DHFL plants showed a significant (p<0.05) decrease in ROS compared to the RW plants. Chloroplast is the primary producer of ROS in green plant parts. This occurs when the photosynthetic electron transport system becomes too active, resulting in an overflow of reduced power. This excess reducing power is responsible for the conversion of oxygen into various ROS [79, 80]. While ROS is formed under normal plant growth conditions, it is significantly amplified under stress conditions Dat J, et al. [81] as observed in the present study. Additionally, mitochondrial electron transport can generate ROS, such as superoxide and H2O2, especially during periods of water stress [82]. The increase in ROS concentration under different water stress conditions was also consistent with various studies conducted. For instance, a study conducted by Radwan A, et al. [83] revealed that when Thymus vulgaris L. plants when exposed to 30-40% of water holding capacity, there was a significant (p<0.05) increase in the concentration of ROS compared to those plants that received 70-80% of water.
Malondialdehyde (MDA) is a well-known biochemical marker that indicates an increase in the activity of ROS and oxidative stress in plant tissues during unfavourable conditions [84]. MDA is regarded as the ultimate outcome of lipid peroxidation and a key signal of oxidative damage that may occur in cellular membranes under water stress conditions [84]. In the present study, MDA concentration was significantly (p<0.05) high i.e., 94% more in FL plants and 47% in DHFL compared to RW plants. In contrast, RHDR plants had significantly (p<0.05) reduced concentrations of MDA compared to those of RW plants. In a similar, Khaleghi A, et al. [7] reported comparable findings of reduced levels of MDA with rehydration of drought-stressed Osage orange (Maclura pomifera) plants. Unexpectedly, the DR plants showed a significantly (p<0.05) the least concentrations of MDA of5% compared to the RW plants although several studies under similar conditions have reported otherwise [7, 84, 85]. These findings might demonstrate that Jamaican thyme has a distinctive adaptive strategy, as well as a strong antioxidant defense system and effective cellular repair.
Total Soluble Sugar
The cellular buildup of osmolytes such as sugars in response to water stress is frequently linked to mechanisms for tolerating water stress [86]. Total soluble sugar (TSS) contents in DR and FL were non-significantly (p<0.05) increased by 60% and 30%, respectively, compared to the RW plants (Table 2), which is similar to previous observations that reported a substantial rise in TSS concentrations in plants under severe water stress [84]. The DHFL plants exhibited a significant (p<0.05) increase of about 119% in TSS concentration compared to the RW plants (Table 2). The regulation of TSS in plants experiencing water stress is influenced by various mechanisms that impact the production and transport of soluble sugars inside the leaves [87]. Furthermore, soluble sugars have a crucial function in regulating osmotic adjustment in plants [88]. For instance, the RHDR plants exhibited a notable reduction in TSS of about 41% compared to the RW plants. In contrast, a study conducted on orchids (Dendrobium moniliforme) revealed that TSS increased in rehydrated plants [89]. In general, it is widely acknowledged that plants respond to water stress by accumulating organic molecules such as sugars, which are known as osmolytes or compatible solutes Oraki H, et al. [90] and Ibrahim M, et al. [91] which are crucial for plants adaptation to water stress [84]. In addition, it can safeguard crucial enzymes and various components of plant cells from harmful oxidation by scavenging reactive oxygen species (ROS) [92].
Conclusion
The occurrence of global climate change and extreme weather patterns has resulted in water stress emerging as a significant constraint to the production of agriculture. The current study provides evidence that water stress has a detrimental effect on plant growth, physiological processes, and chemical composition. The timing, duration, and amount of growing medium water level determine the severity of water stress that influences a plant’s growth and development performance. The study also suggests that water stress severely affects the photosynthetic machinery of Jamaican thyme which may lead to reduced plant growth and development. The results demonstrated a swift recovery process upon rehydration of drought-stricken plants or dehydration of waterlogged plants, returning them to their typical growth circumstances. To deepen our understanding of these plant adaptations and clarify the precise metabolic responses that enable water stress tolerance, we suggest conducting additional experiments that encompass a broader spectrum of water stress levels. This will facilitate a more extensive investigation of plant response and provide more profound insights into the underlying plant physiologic and metabolic alterations associated with water stress tolerance.
References
-
Amna, Ali B, Azeem MA, Qayyum A, Mustafa G, et al. (2021) Bio-Fabricated Silver Nanoparticles: A Sustainable Approach for Augmentation of Plant Growth and Pathogen Control. In: Faizan M, Hayat S, et al. (Eds.), Sustainable Agriculture Reviews 53 Nanoparticles: A New Tool to Enhance Stress Tolerance 53: 345-371.
-
Zainab N, Amna, Khan AA, Azeem MA, Ali B, et al. (2021) PGPR-Mediated Plant Growth Attributes and Metal Extraction Ability of Sesbania sesban L. in Industrially Contaminated Soils. Agronomy 11(9): 1-19.
-
Lesk C, Rowhani P, Ramankutty N (2016) Influence of extreme weather disasters on global crop production. Nature 529(7584): 84-87.
-
Gupta A, Medina RA, Delgado CAI (2020) The physiology of plant responses to drought. Science 368(6488): 266- 269.
-
Du W, FitzGerald GJ, Clark M, Hou XY (2010) Health impacts of floods. Prehospital and Disaster Medicine 25(3): 265-272.
-
Shi W, Wang M, Liu Y (2021) Crop yield and production responses to climate disasters in China. Science of The Total Environment 750: 141147.
-
Khaleghi A, Naderi R, Brunetti C, Maserti BE, Salami SA, et al. (2019) Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Scientific Reports 9(1): 19250.
-
Ren B, Zhang J, Dong S, Liu P, Zhao B (2018) Responses of carbon metabolism and antioxidant system of summer maize to waterlogging at different stages. Journal of Agronomy and Crop Science 204(5): 505-514.
-
Bista DR, Heckathorn SA, Jayawardena DM, Boldt JK (2020) Effect of drought and carbon dioxide on nutrient uptake and levels of nutrient-uptake proteins in roots of barley. American Journal of Botany 107(10): 1401-1409.
-
Jiao P, Wu Z, Wang X, Jiang Z, Wang Y, et al. (2021) Short- term transcriptomic responses of Populus euphratica roots and leaves to drought stress. Journal of Forestry Research 32: 841-853.
-
Waszczak C, Carmody M, Kangasjärvi J (2018) Reactive Oxygen Species in Plant Signaling. Annual Review of Plant Biology 69: 209-236.
-
Bartels D, Sunkar R (2005) Drought and salt tolerance in plants. Critical Reviews in Plant Sciences 24(1): 23-58.
-
Ashraf M, Foolad MR (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and Experimental Botany 59(2): 206- 216.
-
Seki M, Umezawa T, Urano K, Shinozaki K (2007) Regulatory metabolic networks in drought stress responses. Current Opinion in Plant Biology 10(3): 296- 302.
-
Tardieu F, Simonneau T, Muller B (2018) The Physiological Basis of Drought Tolerance in Crop Plants: A Scenario-Dependent Probabilistic Approach. Annual Review of Plant Biology 69(1): 733-759.
-
Fu J, Huang B (2001) Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environmental and Experimental Botany 45(2): 105-114.
-
Yin C, Peng Y, Zang R, Zhu Y, Li C (2005) Adaptive responses of Populus kangdingensis to drought stress. Physiologia Plantarum 123(4): 445-451.
-
Lukhoba CW, Simmonds MSJ, Paton AJ (2006) Plectranthus: A review of ethnobotanical uses. Journal of Ethnopharmacology 103(1): 1-24.
-
Abbey Lord, Ofoe R, Wang Z, Chada S (2023) How Central Carbon Metabolites of Mexican Mint (Plectranthus amboinicus) Plants Are Impacted under Different Watering Regimes. Metabolites 13(4): 539.
-
Reddy AR, Chaitanya KV, Vivekanandan M (2004a) Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. Journal of Plant Physiology 161(11): 1189-1202.
-
Lichtenthaler HK (1987) [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology 148: 350-382.
-
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric Method for Determination of Sugars and Related Substances. Analytical Chemistry 28(3): 350-356.
-
Ainsworth EA, Gillespie KM (2007) Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nature Protocols 2(4): 875-877.
-
Chang CC, Yang MH, Wen HM, Chern JC (2020) Estimation of total flavonoid content in propolis by two complementary colometric methods. Journal of Food and Drug Analysis 10(3).
-
Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207(4): 604-611.
-
Fan D, Subramanian S, Smith DL (2020) Plant endophytes promote growth and alleviate salt stress in Arabidopsis thaliana. Scientific Reports 10(1): 12740.
-
Patterson BD, MacRae EA, Ferguson IB (1984) Estimation of hydrogen peroxide in plant extracts using titanium (IV). Analytical Biochemistry 139(2): 487-492.
-
Baher ZF, Mirza M, Ghorbanli M, Bagher Rezaii M (2002) The influence of water stress on plant height, herbal and essential oil yield and composition in Satureja hortensis L. Flavour and Fragrance Journal 17(4): 275-277.
-
Khorasani H, Rajabzadeh F, Mozafari H, Pirbalouti AG (2023) Water deficit stress impairment of morphophysiological and phytochemical traits of Stevia (Stevia rebaudiana Bertoni) buffered by humic acid application. South African Journal of Botany 154: 365- 371.
-
Zarei F, Hezarjaribi A, Khorasaninejad S, Zakerinia M (2021) The effect of foliar application of humic acid on increasing Stevia rebaudiana tolerance under different irrigation regimes. Iranian Journal of Rangelands and Forests Plant Breeding and Genetic Research 28(2): 281- 297.
-
Lugojan C, Ciulca S (2011) Evaluation of relative water content in winter wheat. Journal of Horticulture, Forestry and Biotechnology 15(2): 173-177.
-
Alordzinu KE, Jiuhao L, Appiah SA, Aasmi AL, Blege PK, et al. (2021) Water stress affects the physio-morphological development of tomato growth. African Journal of Agricultural Research 17(5): 733-742.
-
Chen S, Zhou Z, Andersen MN, Hu T (2015) Tomato yield and water use efficiency–coupling effects between growth stage specific soil water deficits. Acta Agriculturae Scandinavica Section B-Soil & Plant Science 65(5): 460-469.
-
El Jaafari S (2000) Durum wheat breeding for abiotic stresses resistance: Defining physiological traits and criteria. Options Mediterranéennes 40(31): 251-256.
-
Boutraa T, Akhkha A, Al-Shoaibi AA, Alhejeli AM (2010) Effect of water stress on growth and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in Saudi Arabia. Journal of Taibah University for Science 3: 39-48.
-
Ezzine H, Metougui ML, Boukcim H, Abbas Y (2023) Physiological responses of three field-grown species (Ceratonia siliqua, Eucalyptus camaldulensis, and Moringa oleifera) to water deficits in a Mediterranean semi-arid climate. Scientific Reports 13(1): 4536.
-
Demidchik V, Straltsova D, Medvedev SS, Pozhvanov GA, Sokolik A, et al. (2014) Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. Journal of Experimental Botany 65(5): 1259-1270.
-
Lee B, Zhu JK (2010) Phenotypic analysis of Arabidopsis mutants: Electrolyte leakage after freezing stress. Cold Spring Harbor Protocols 2010(1): 4970.
-
Babaei K, Moghaddam M, Farhadi N, Ghasemi Pirbalouti A (2021) Morphological, physiological and phytochemical responses of Mexican marigold (Tagetes minuta L.) to drought stress. Scientia Horticulturae 284: 110116.
-
Leopold AC, Musgrave ME, Williams KM (1981) Solute leakage resulting from leaf desiccation. Plant Physiology 68(6): 1222-1225.
-
Zahedi SM, Hosseini MS, Fahadi Hoveizeh N, Kadkhodaei S, Vaculík M (2023) Physiological and Biochemical Responses of Commercial Strawberry Cultivars under Optimal and Drought Stress Conditions. Plants 12(3): 496.
-
Pirasteh-Anosheh H, Saed-Moucheshi A, Pakniyat H, Pessarakli M (2016) Stomatal responses to drought stress. In: Ahmad P (Ed.), Water Stress and Crop Plants, John Wiley & Sons pp: 24-40.
-
Clijsters H, Van Assche F (1985) Inhibition of photosynthesis by heavy metals. Photosynthesis Research 7(1): 31-40.
-
Allakhverdiev SI, Murata N (2004) Environmental stress inhibits the synthesis de novo of proteins involved in the photodamage–repair cycle of photosystem II in Synechocystis sp. PCC 6803. Biochim Biophys Acta 1657(1): 23-32.
-
Krause G, Weis E (1991) Chlorophyll fluorescence and photosynthesis: The basics. Annual Review of Plant Biology 42(1): 313-349.
-
Urban L, Aarrouf J, Bidel LP (2017) Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Frontiers in Plant Science 8: 2068.
-
Lichtenthaler HK, Rinderle U (1988) The role of chlorophyll fluorescence in the detection of stress conditions in plants. CRC Critical Reviews in Analytical Chemistry 19(1): S29-S85.
-
Porcar-Castell A, Tyystjärvi E, Atherton J, Van der Tol C, Flexas J, et al. (2014) Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. Journal of Experimental Botany 65(15): 4065-4095.
-
Meng LL, Song JF, Wen J, Zhang J, Wei JH (2016) Effects of drought stress on fluorescence characteristics of photosystem II in leaves of Plectranthus scutellarioides. Photosynthetica 54(3): 414-421.
-
Zhuang J, Wang Y, Chi Y, Zhou L, Chen J, et al. (2020) Drought stress strengthens the link between chlorophyll fluorescence parameters and photosynthetic traits. Peer J 8: e10046.
-
Mohammed GH, Zarco-Tejada P, Miller JR (2003) Applications of chlorophyll fluorescence in forestry and ecophysiology. In: Practical applications of chlorophyll fluorescence in plant biology, Springer, pp: 79-124.
-
Kumar D, Singh H, Raj S, Soni V (2020) Chlorophyll a fluorescence kinetics of mung bean (Vigna radiata L.) grown under artificial continuous light. Biochemistry and Biophysics Reports 24: 100813.
-
Osakabe Y, Osakabe K, Shinozaki K, Tran LS (2014) Response of plants to water stress. Frontiers in Plant Science 5.
-
Moradi P, Ford-Lloyd B, Pritchard J (2014) Plant-water responses of different medicinal plant thyme (Thymus spp.) species to drought stress condition. Australian Journal of Crop Science 8(5): 666-673.
-
Subrahmanyam D, Subash N, Haris A, Sikka AK (2006) Influence of water stress on leaf photosynthetic characteristics in wheat cultivars differing in their susceptibility to drought. Photosynthetica 44(1): 125- 129.
-
Ma SC, Duan AW, Wang R, Guan ZM, Yang SJ, et al. (2015) Root-sourced signal and photosynthetic traits, dry matter accumulation and remobilization, and yield stability in winter wheat as affected by regulated deficit irrigation. Agricultural Water Management 148: 123- 129.
-
Liu EK, Mei XR, Yan CR, Gong DZ, Zhang YQ (2016) Effects of water stress on photosynthetic characteristics, dry matter translocation and WUE in two winter wheat genotypes. Agricultural Water Management 167: 75-85.
-
Izanloo A, Condon AG, Langridge P, Tester M, Schnurbusch T (2008) Different mechanisms of adaptation to cyclic water stress in two South Australian bread wheat cultivars. Journal of Experimental Botany 59(12): 3327- 3346.
-
Carmo-Silva AE, Keys AJ, Andralojc PJ, Powers SJ, Arrabaca MC, et al. (2010) Rubisco activities, properties, and regulation in three different C4 grasses under drought. Journal of Experimental Botany 61(9): 2355- 2366.
-
Harb A, Krishnan A, Ambavaram MM, Pereira A (2010) Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiology 154(3): 1254-1271.
-
Yan W, Zhong Y, Shangguan Z (2016) A meta-analysis of leaf gas exchange and water status responses to drought. Scientific Reports 6(1): 20917.
-
Bota J, Medrano H, Flexas J (2004) Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress?. New Phytologist 162(3): 671-681.
-
Enneb H, Yahya LB, Ilyas M, Dhale DA, Bagues M, et al. (2020) Influence of Water Stress on Growth, Chlorophyll Contents and Solute Accumulation in Three Accessions of Vicia faba L. from Tunisian Arid Region. In Abiotic Stress in Plants.
-
Mafakheri A, Siosemardeh AF, Bahramnejad B, Struik PC, Sohrabi Y (2010) Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Australian Journal of Crop Science 4(8): 580- 585.
-
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009) Plant Drought Stress: Effects, Mechanisms and Management. In: Lichtfouse E, Navarrete M, et al. (Eds.), Sustainable Agriculture, Springer, Netherlands, pp: 153- 188.
-
Yusuf N, Hamed NFI (2021) Effects of Water Deficit on the Growth and Chlorophyll Content of Capsicum Frutescens. Journal of Sustainability Science and Management 16(6): 148-158.
-
Reddy AR, Chaitanya KV, Vivekanandan M (2004) Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. Journal of Plant Physiology 161(11): 1189-1202.
-
Basu S, Ramegowda V, Kumar A, Pereira A (2016) Plant adaptation to drought stress. F1000Research.
-
Mohammadkhani N, Heidari R (2007) Effects of water stress on respiration, photosynthetic pigments and water content in two maize cultivars. Pakistan Journal of Biological Sciences 10: 4022-4028.
-
Deng X, Hu ZA, Wang HX, Wen XG, Kuang TY (2003) A comparison of photosynthetic apparatus of the detached leaves of the resurrection plant Boea hygrometrica with its non-tolerant relative Chirita heterotrichia in response to dehydration and rehydration. Plant Science 165(4): 851-861.
-
Qiu J, Gao F, Shen G, Li C, Han X, et al. (2013) Metabolic engineering of the phenylpropanoid pathway enhances the antioxidant capacity of Saussurea involucrata. PLoS One 8(8): e70665.
-
De Lima VT, De Paula Quintão Scalon S, Cardoso CAL, Reis LC, Kolb RM (2023) Does flooding affect the survival and primary and secondary metabolism of two species of Croton?. Trees 37(4): 1081-1095.
-
Pirbalouti AG, Malekpoor F, Salimi A, Golparvar A, Hamedi B (2017) Effects of foliar of the application chitosan and reduced irrigation on essential oil yield, total phenol content and antioxidant activity of extracts from green and purple basil. Acta Scientiarum Polonorum Hortorum Cultus 16(6).
-
Caser M, Chitarra W, D’Angiolillo F, Perrone I, Demasi S, et al. (2019) Drought stress adaptation modulates plant secondary metabolite production in Salvia dolomitica Codd. Industrial Crops and Products 129: 85-96.
-
Per TS, Khan NA, Reddy PS, Masood A, Hasanuzzaman M, et al. (2017) Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiology and Biochemistry 115: 126-140.
-
Tang Y, Sun X, Wen T, Liu M, Yang M, et al. (2017) Implications of terminal oxidase function in regulation of salicylic acid on soybean seedling photosynthetic performance under water stress. Plant Physiology and Biochemistry 112: 19-28.
-
Atkinson NJ, Urwin PE (2012) The interaction of plant biotic and abiotic stresses: From genes to the field. Journal of Experimental Botany 63(10): 3523-3543.
-
Lehmann M, Schwarzländer M, Obata T, Sirikantaramas S, Burow M, et al. (2009) The Metabolic Response of Arabidopsis Roots to Oxidative Stress is Distinct from that of Heterotrophic Cells in Culture and Highlights a Complex Relationship between the Levels of Transcripts, Metabolites, and Flux. Molecular Plant 2(3): 390-406.
-
Foyer CH, Descourvières P, Kunert KJ (1994) Protection against oxygen radicals: An important defence mechanism studied in transgenic plants. Plant Cell & Environment 17(5): 507-523.
-
Foyer CH, Noctor G (2003) Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiologia Plantarum 119(3): 355-364.
-
Dat J, Vandenabeele S, Vranová E, Van Montagu M, Inzé D, et al. (2000) Dual action of the active oxygen species during plant stress responses. Cellular and Molecular Life Sciences 57(5): 779-795.
-
Elstner EF, Osswald W (1994) Mechanisms of oxygen activation during plant stress. Proceedings of the Royal Society of Edinburgh, Section B: Biological Sciences 102: 131-154.
-
Radwan A, Kleinwächter M, Selmar D (2017) Impact of drought stress on specialised metabolism: Biosynthesis and the expression of monoterpene synthases in sage (Salvia officinalis). Phytochemistry 141: 20-26.
-
Farag R, Abd Elbar O, Shehata S (2019) Impact of Drought Stress on Some Growth, Biochemical and Anatomical Parameters of Thymus Vulgaris L. Arab Universities. Journal of Agricultural Sciences 27: 37-50.
-
Sarker U, Oba S (2018) Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Scientific Reports 8(1): 16496.
-
Pirzad A, Shakiba MR, Zehtab-Salmasi S, Mohammadi SA, Darvishzadeh R, et al. (2011) Effect of water stress on leaf relative water content, chlorophyll, proline and soluble carbohydrates in Matricaria chamomilla L. Journal of Medicinal Plants Research 5(12): 2483-2488.
-
Arabzadeh N (2012) The effect of drought stress on soluble carbohydrates (sugars) in two species of Haloxylon persicum and Haloxylon aphyllum. Asian Journal of Plant Sciences 11(1): 44-51.
-
Zhou Q, Yu BJ (2009) Accumulation of inorganic and organic osmolytes and their role in osmotic adjustment in NaCl-stressed vetiver grass seedlings. Russian Journal of Plant Physiology 56: 678-685.
-
Wu X, Yuan J, Luo A, Chen Y, Fan Y (2016) Drought stress and re-watering increase secondary metabolites and enzyme activity in dendrobium moniliforme. Industrial Crops and Products 94: 385-393.
-
Oraki H, Aghaalikhana M (2012) Effect of water deficit stress on proline contents, soluble sugars, chlorophyll and grain yield of sunflower (Helianthus annuus L.) hybrids. African Journal of Biotechnology 11(1): 164- 168.
-
Ibrahim M (2014) Induced drought resistance in common bean (Phaseolus vulgaris L.) by exogenous application with active yeast suspension. Middle East J Appl Sci 4(4): 806-815.
-
Farooq M, Hussain M, Wahid A, Siddique KHM (2012) Drought stress in plants: An overview. Plant Responses to Drought Stress pp: 1-33.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note