Mechanism of Antibacterial Activities of Cu (â…¡) Ions against Staphylococcus aureus and Escherichia coli on the Ground of Results Obtained from Dilution Medium Method
Antibacterial activities by copper(â…¡) ion solution based antibacterial results of broth dilution medium method were investigated. From dilution medium method, Minimum Inhibitory Concentration (MIC) = 625 mg/L as bacteriostatic action, Minimum Bactericide Concentration (MBC) = 1250 mg/L as bactericide action by Cu2+ ion solution were obtained against Staphylococcus aureus. Bacteriolysis of Staphylococcus aureus peptidoglycan (PGN) cell wall by Cu2+ ions is ascribed to the inhibition of PGN elongation due to the damages of PGN biosynthesis of transglycosylase (TG) and transpeptidase (TP), and the activations of PGN forth autolysins. The other, bacteriolysis of Escherichia coli outer membrane cell wall by Cu2+ ions is attributed to the destruction of outer membrane structure and to the inhibition of PGN elongation due to the damage of PGN biosynthesis TP and the activations of PGN autolysins. Cu2+ ions induced ROS such as O2ï¼, H2O2,ï½¥OH, OHï¼ producing in bacterial cell wall occur oxidative stress.
Introduction
Silver, copper, and zinc of transition metals have highly antibacterial activities and are utilized as chemotherapy agents. Recently, antibacterial activities of copper, zinc and these complexes call attention to potential treatments such as control of infectious diseases [1, 2], exploitation during bacterial pathogenesis [3, 4, 5], adaptive immune response to virus [6] and binds with viral proteins [7], Mechanism of Antibacterial Activities of Cu (Ⅱ) Ions against Staphylococcus aureus and Escherichia coli on the Ground of Results Obtained from Dilution Medium Method novel copper-based formulations on such as E.coli O157, Salmonella, Helicobacter pylori [8, 9], and regulation of cancer and tumor cells [10, 11]. Copper is required for the function of several important enzymes including the cytochrome oxidase, copper-zinc superoxidase. Copper is viral inhibitors with the commonest metal ion that binds to viral proteins. However, copper can also be extremely Virol Immunol J
toxic, requiring homeostatic mechanism. Research of Cu ions killing mechanism has been carried out that Cu ions induced lead to the instability of membrane fatty acid integrity and homeostasis, increased levels of lipid peroxidation against both Gram-negative and Gram- positive bacteria [12]. In this way, the detailed mechanisms by which copper ions enter the bacterial cells are scarcely known. In this study, the broth dilution medium method test against S.aureus and E.coli, were carried out, where in it was turned out that antibacterial effects of Cu2+ ion solution were examined. On the basis of the high antibacterial activities for these Cu2+ ions, the processes of bacteriolysis and destructions of bacterial cell walls by copper (Ⅱ) ions had been considered against S. aureus peptidoglycan (PGN) and E.coli outer membrane cell walls. Furthermore, the bacteriolytic mechanisms by copper (Ⅱ) ion have been also revealed against both S.aureus and E.coli bacteria.
Method
Two-fold Broth Dilution Medium Method Tests for Cu2+ Ion Solutions
This method is quantitatively obtained for the antibacterial activity on the bactericidal assay. Bacteria intended for two-fold broth dilution medium method were treated as Staphylococcus aureus (NBRC12732) and Escherichia coli (ATCC25922). The other, the antibacterial copper ion of commercial copper (Ⅱ) ion agent (Japan ion production Ltd., original Cu2+ ion solution; 500 mg/L) are used as bacteriostasis, and copper nitrate (Cu(NO3)23H2O,Wako Pure Reagents) of special class reagent was used as bactericide action. Firstly, the sample test tube of Cu2+ ion concentration of 10,000 mg/L have been prepared in heart infusion agar medium(Nissui). Next, the diluted solutions of 10-stagesby two-fold dilution solution method was adjusted in tenth sample tubes for Cu2+ ion solution concentration of 9.8~5,000 mg/L. Afterwards, the adjustment solution within final solution of 5×105cfu/mL was prepared, and then with a sterile micropipette, fungous liquid 1 mL of bacterial suspension was respectively transferred from tube No 1 to other tubes that were inoculated into the respective tubes. Finally, the tubes were incubated at 35℃ for 24 hours, in which the incubated solutions were afforded to minimum inhibitory concentration (MIC), minimum bactericide concentration (MBC), colony forming unit (CFU) measurements.
Search and Analysis for Bacterial Cell Molecular Structure, and PGN Biosyntheses and Autolysins
The surface envelop cell structures of S.aureus as representative of Gram-positive bacterium and E.coli as representative of Gram-negative bacterium, molecular structures of these cell walls, molecular structure of peptidoglycan (PGN), and PGN biosyntheses and autolysins were searched in detail. Further, the reaction and the behavior of metallic ions and bacterial cell, molecular bonding manner, and copper ion characteristics were also searched.
Results
Bacteriostatic and Bactericide Actions for Cu2+ Ionsolution by the Broth Dilution Medium Method
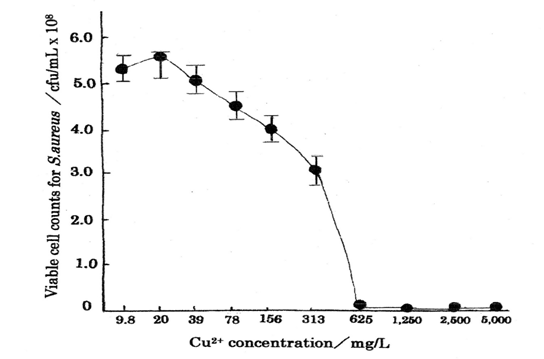
Table 1 shows the bacteriostasis as disinfection agent inhibiting the bacteria growth and multiplying organism of Cu2+ ion, in which minimum inhibitory concentration, MIC=50 mg/L above was obtained for Cu2+ ion concentration range of 0.10~50 mg/L [13]. The other, Table 2 indicates the results as bactericide action, in which MIC=625 mg/L and minimum bactericide concentration, MBC=1250mg/L were obtained for Cu2+ ion concentration range of 9.8~5000 mg/L [14]. The killing curve of Cu2+ ions is shown in Fig.1 (measurement’s error=±6%), in which killing effects for the copper (Ⅱ) ions appear sufficiently.
| Cu2+ solution concentration(mg/L) | MIC 50 mg/L above | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cu2+ solution agent·original conc 500 mg/L | MIC 50 mg/L above | 50 | 25 | 12.5 | 6.25 | 3.13 | 1.56 | 0.78 | 0.39 | 0.2 | 0.1 |
| Cu2+ solution agent·original conc 500 mg/L | MIC 50 mg/L above | + | + | + | + | + | + | + | + | + | + |
Table 1: MIC measurements of commercial Cu2+ solution agents as a bacteriostatic action against E.coli by broth dilution (+): Vis
| Antibacterial agent Cu(NO3)23H2O solution | Cu2+ concentration(mg/L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Antibacterial agent Cu(NO3)23H2O solution | 5000 | 2500 | 1250 | 625 | 313 | 156 | 78 | 39 | 20 | 9.8 |
| MIC | - | - | - | - | + | + | + | + | + | + |
| MBC | - | - | - | + | + | + | + | + | + | + |
| CFU(cfu/mL) | <10 | <10 | <10 | 1.1×10^{2}$ | $3.1×10^{8}$ | $4.0×10^{8}$ | $4.5×10^{8}$ | $5.1×10^{8}$ | $5.5×10^{8}$ | $5.3×10^{8}$ |
Table 2: MIC, MBC, and CFU of Cu2+ in Cu (NO3)23H2O solution as a bactericidal action against S.aureus by 10-fold diluted (+): Ba
Table 2: MIC, MBC, and CFU of Cu2+ in Cu (NO3)23H2O solution as a bactericidal action against S.aureus by 10-fold diluted (+): Bacterial growth (visible turbidity), (-): No visible bacterial growth Table 2: MIC, MBC, and CFU of Cu2+ in Cu (NO3)23H2O solution as a bactericidal action against S.aureus by 10-fold diluted solution medium method.
Results of Search and Analysis
S.aureus and E.coli Cell walls, Action Sites of PGN biosyntheses of transglycosylase TG and transpeptidase TP and PGN autolysins: S.aureus surface cell envelop consists of teichoic acids, lipoteichoic acids, and thick peptidoglycan (below PGN) cell wall [13], whereas E.coli cell wall comprised of lipid A, lipopoly- saccharide, porin proteins, outer membrane of lipoprotein, and thinner 2-7 nm PGN layer in 30-70 nm periplasmic space [15]. Figure 2 shows the molecular structure of S.aureus PGN cell wall, including the action sites of PGN biosynthesis enzymes of TG/TP, and PGN forth autolysins and Lysostaphin enzyme. Furthermore, Figure 3 represents the molecular structure of E.coli cell wall and periplasmic peptidoglycan, containing the action sites of the hydrolases of lipoproteins, the peptidoglycan biosynthetic enzymes TG/TP, and the autolysins. Further, interactions of PGN molecular structure, PGN syntheses andautolysins influence essentially in any event the bacteriolysis of bacterial cell walls.
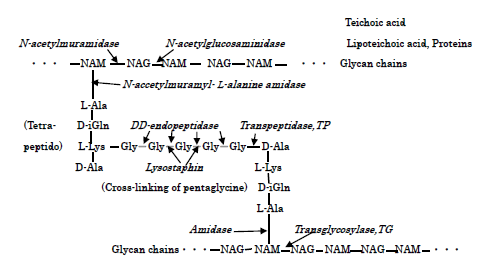
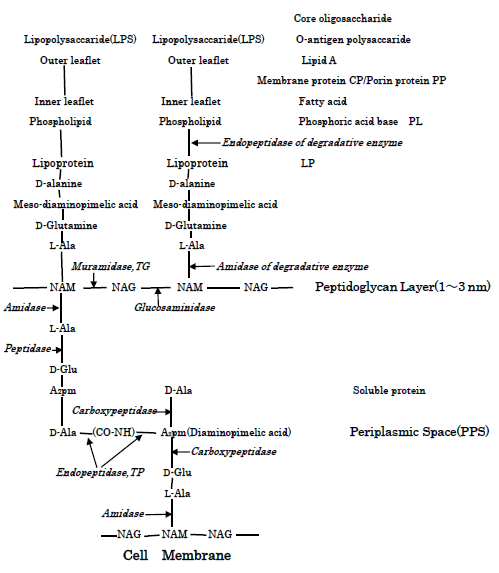
Characteristics of Copper (Ⅱ) Ion Solution: In this experiment, Cu (Ⅱ) ion solution was used as antibacterial agent, in which the rapid killing of various bacteria in contact with metallic copper (Ⅱ) ion is thought to be influenced by the Cu (Ⅱ)/Cu (Ⅰ) redox reaction strength within the bacterial cell. Then, for example, by the reaction of Cu2+ ions with S.aureus surface, Cu2+-proteins may be formed, on the ground that is due to formation of S-atom containing Cu-cysteine complex in bacteria [16].
Discussions
Bacteriolysis of S.aureus PGN Cell Wall by Cu2+ Ions
Bacteriolysis by balance deletion between biosynthesis enzyme and decomposition enzyme (autolysin) in PGN cell wall: For the sake of growth of S.aureus PGN cell wall, there is necessarily required for the adequate balance between PGN biosynthesis and PGN autolysin. When the balance is broken by Cu2+ penetration, Cu2+ ions are self-catalytically treated as coenzyme, that this is indicated that activation of autolysin is preceded, in which bacteriolysis and killing may result. Hence, bacteriolysis of S.aureus PGN cell wall by Cu2+ions is thought to be due to inhibition of PGN elongation owing to the damages of PGN synthetic TG/TP [17] and the activations of PGN autolysin, AmiA [18, 19]. Inhibition of Polymerization of Glycan Chains Bonding and Cross-Linking of Side Peptide: Cu2+ ions inhibit polymerization of glycan chains, forming copper complex in which is partial action sites of glycan saccharide chains. L is coordinated molecular.
Cu2+ + LH → CuL+ + H+
CuL+ + LH → CuL2 + H+
Copper-complexes on saccharide chains may be ・―NAG-(NAM-Cu-2O-2N-NAG)-NAM―・ [20]. The other, Cu2+ ions inhibit cross-linked reaction by peptide copper complex formation bonding to side- peptide chains [21].
Cu2+ + 2LH → CuL2 + H+
Peptide copper complex may be 3N-Cu-O, Cu (Gly-L-Ala) H2O [20]. Specially, Cu2+ ions react with cross-molecular penta glycine (Gly)5, copper-glycine complex may be formed [22]. Amino acid:Cu2+ + Gly-→ Cu (Gly)+, Cu (Gly)+ + Gly― → Cu(Gly)2, Peptido: Cu2+ + GlyGly→ Cu (GlyGly), Cu (GlyGly) + Gly―→Cu(GlyGlyGly)―.
Bacteriolysis and Destruction of E.coli Outer Membrane Cell Wall by Cu2+ Ions
Inhibition of outer membrane cell wall: Cu2+ ions inactivate catalyst enzyme with forming Cu+ ions.
Cu2+ + -SH → -SCu (Ⅰ) + H+ By the penetration of Cu2+ ions, as shown in Fig.3, the activations of amidase enzyme of N-terminal and endopeptidase enzyme of C-terminal are enhanced [23, 24]. Accordingly, the activations of decomposition at N-, C-terminals of lipoproteins may occur with the destruction of outer membrane structure.
Inhibition of Biosynthesis and Activation of Autolysin, or Regulation and Deletion of Autolysin: Inhibition of E.coli PGN by Cu2+ ions is reported [25], however, the site of concrete action is not described. In E.coli, it is unlikely thought that Cu2+ ions inhibit both TG and TP [26]. The other, it is unclear that Cu2+ inhibits the polymerization of NAM and NAG chains. It is perhaps simpler to think that TP enzyme of cross-linked reaction is inhibited by Cu2+ ions and the activation of PGN autolysin occurs. By the accumulation of Cu2+ ions in periplasmic space, it might be possible that bacteriolysis of cell wall occur by the activation of PGN autolysin within periplasmic space. Many autolysins of E.coli are regulated by metals ion such as Hg2+, Cu2+ [27]. This regulation or deletion of decomposition enzyme inhibits PGN elongation, in which the bacteriolysis of the cell wall is induced. These facts are consistent with that the destruction by bacteriolysis of cell wall had been observed against E.coli. Hence, bacteriolysis of E.coli cell wall by Cu2+ ions occurs by destruction of outer membrane structure due to degradation of lipoprotein at N-, C-terminals, damage of TP enzyme and activations of PGN autolysins [19]. Furthermore, deletion of PGN autolysin also becomes bacteriolystic factor. Antibacterial activities of cell membrane and cytoplasm: Copper in solution is present as cupric ions, Cu2+. The Cu2+ in bacteria is reduced to Cu1+ and that the cuprous ions are considerably more toxic and very unstable. In E.coli cell wall, reactive oxygen species (ROS) production occur by Cu2+ ion penetration.
-・ Cu2+ + O2
O2 + e → O2
-→ O2 + Cu+ O2
- + e + 2H+ → H2O2 Cu2+ + H2O2 → Cu+ + H+ + HO2・ ROS O2
-and H2O2 generated in the cell wall, permeate into cell membrane and cytoplasm, in which high reactive・ OH and OH- in cell membrane are formed by Haber-Weiss and Fenton reactions. Haber-Weiss reaction [28];
-→ ・OH + OH- + O2 Fenton reaction [29];
H2O2 + O2
Cu+ + H2O2 → ・OH + OH-+ Cu2+
Furthermore, new ROS productions occur by Fenton-like type. L=Ligand
LCu (Ⅱ) + H2O2 → LCu (Ⅰ) + ・OOH + H+
LCu (Ⅰ)+H2O2 → LCu (Ⅱ)+・OH + OH― Lipid peroxidation and the radical are accumulated, in which cell membrane oxidation and the decreasing function induce. Superoxide anion radical O2―thathydroxyl radical・OH induce ROS production, the - induce SOD production by hydrogen peroxide H2O2. Especially, and high toxic hydroxyl radical・OH can damage a range of cellular macromolecules, including causing mutations in DNA [30]. As above-mentioned, the bactericidal processes of bacteriolysis of the S.aureus and E.coli cell walls by Cu2+ ions, and also the antibacterial activities of cell membrane and cytoplasm are shown in Table 3.
other O2
| Cell wall | Cell membrane | Cytoplasm | |||
|---|---|---|---|---|---|
| Cu2+ | S.aureus cell wall | • Cu-ammine complex and Cu-protein formationsCu+, Cu2+O2- H2O2• OH OH- (O2-) H2O2 | • Cu-complex and Cu-3N-O complex formations Cu2+• OH OH- (O2-) H2O2 | ||
| Cu2+ | PGN layer cell wall | • Cu-ammine complex and Cu-protein formationsCu+, Cu2+O2- H2O2• OH OH- (O2-) H2O2 | • Cu-complex and Cu-3N-O complex formations Cu2+• OH OH- (O2-) H2O2 | ||
| Cu2+ | Cu2+, Cu+• Bacteriolysis of PGN cell wall by TG,TP synthesis inhibitions and activations of S. Aureus PGN autolysins• Cu complex of glycan saccharide chain and Cu peptide complex formations• Reactive oxygen species O2-,H2O2 | • Cu-ammine complex and Cu-protein formationsCu+, Cu2+O2- H2O2• OH OH- (O2-) H2O2 | • Cu-complex and Cu-3N-O complex formations Cu2+• OH OH- (O2-) H2O2 | ||
| Cu2+ | Cu2+ | • Cu-ammine complex and Cu-protein formationsCu+, Cu2+O2- H2O2• OH OH- (O2-) H2O2 | • Cu-complex and Cu-3N-O complex formations Cu2+• OH OH- (O2-) H2O2 | E.coli cell wall | |
| Lipopolysaccharide (LPS) | Cu2+ | • Cu-ammine complex and Cu-protein formationsCu+, Cu2+O2- H2O2• OH OH- (O2-) H2O2 | • Cu-complex and Cu-3N-O complex formations Cu2+• OH OH- (O2-) H2O2 | Outer membrane(OM) Lipoprotein(LPT) | Periplasmic space(PS) |
| Core polysaccharide Phosphorus lipid, | Cu2+ | • Cu-ammine complex and Cu-protein formationsCu+, Cu2+O2- H2O2• OH OH- (O2-) H2O2 | • Cu-complex and Cu-3N-O complex formations Cu2+• OH OH- (O2-) H2O2 | OmpF, A,C, Porin, Protein | PGN layer Miscible protein |
| Cu2+,Cu+• Variation of Charge properties• Cu ion induced increase in permeability• O2-, Cu+ | Cu2+ | • Cu-ammine complex and Cu-protein formationsCu+, Cu2+O2- H2O2• OH OH- (O2-) H2O2 | • Cu-complex and Cu-3N-O complex formations Cu2+• OH OH- (O2-) H2O2 | Cu2+, Cu+• Cu ion binding proteins• Damages of outer membrane structure due to the degradable enzymes of lipoprotein at N-, C-terminals and the activation of PGN hydrolases• Cu+, H2O2 | Cu2+, Cu+• Cu2+ ions accumulation• Inhibitions of PGN elongation by the deletions of PGN-TP Enzyme and PGN autolysins• Cu+, HO•, H2O2 |
Table 3: Bactericidal action processes of Cu2+ solutions within the cell wall /the cellmembrane /the cytoplasm against Table 3: B
Conclusions
1. From the result of antibacterial activities of Cu2+ ion solution by the two-fold broth dilution medium method, for bacteriostasis MIC=50mg/L above was obtained in Cu2+ concentration range of 0.10~50 mg/L against E.coli. The other, for bactericide action MIC=625 mg/L and MBC=1250 mg/L were obtained in Cu2+ concentration range of 9.8~5,000 mg/L against S.aureus.
2. Bacteriolysis of S.aureus PGN cell wall by Cu2+ ions is caused for the inhibition of PGN elongation due to damages of PGN synthetic TG/TP and activation of PGN autolysins. The other, bacteriolysis of E.coli outer membrane cell wall by Cu2+ ions is attributed to the destruction of outer membrane structure and to the inhibition of PGN elongation due to the damage of PGN biosynthesis TP and the activation of PGN autolysins. 3. By the penetration of copper ions into bacterial cell -, H+, H2O2, ONOO- occurs. The other, in E.coli cell wall, the productions of O2 wall, productions of O2 -, H+ in outer membrane, and H2O2, OH-, ・OH in periplasmic space occur. These ROS and H2O2 damage the cell membrane and the DNA molecules by oxidase stress.
References
-
Michels HT, Wilks SA, Noyce JO, Keevil CW (2005) Copper Alloys for Human Infectious Disease Control.
-
Huang HI, Shih HY, Lee CM, Yang TC, Lay JJ, et al. (2008) In vitro efficacy of copper and silver ions in eradicating Pseudomonas aeruginosa, Stenotrophomonas matophilia and Acinetobacter baumannii: Implications for on-site disinfection for hospital infection control. Water Research 42(1-2): 73-80.
-
Ma L, Terwilliger A, Maresso AW (2015) Iron and Zinc Exploitation during Bacterial Pathogenesis, Metallomics 7(12): 1541-1554.
-
Sarin S, Khamsri B, Sarin C (2011) Isolation of Biosurfactant-Producing Bacteria with Antimicrobial Activity against Bacterial Pathogens, Environment Asia 4(1): 1-5.
-
Ibrahim M, Wang F, Lou MM, Xie GL, Li B et al. (2011) Copper as an antibacterial agent for human pathogenic multidrug resistant Burkholderia cepacia complex bacteria. J Bioscience and bioengineering 112(6): 570-576.
-
Braciale TJ, Hahn YS (2013) Immunity to viruses. Immunol Rev 255(1): p1-11.
-
Chaturvedi UC, Shrivastava R (2005) Interaction of vital proteins with metal ions; role in maintaining the structure and functions of viruses. FEMS Immunology and Medical Microbiology 43(2): 105-114.
-
Warnes SL, Caves V, Keevil CW (2012) Mechanism of copper surface toxicity in _E.coli_ O157:H7 and Salmonella involves immediate membrane depolarization followed by slower rate of DNA destruction which differs from that observed for Gram-positive bacteria. Environmental Microbiology 14(7): 1730-1743.
-
Saracino HM, Zaccaro C, Re GL, Vaira D, Holton J (2013) The effects of two novel copper-based formations on Hericobacter pylori. Antibiotics 2: 265- 273.
-
Skrajnowska D, Bobrowska B, Tokarz A, Kuras M, Rybicki P, et al. (2011) The Effect of Zinc- and Copper Sulphate Supplementation on Tumor and Hair Concentration of Trace Elements (Zn, Cu, Fe, Ca, Mg, P) In Rats with DMBA-Induced Breast Cancer. Pol J Environ Stud 20(6): 1585-1592.
-
Elsen JMH, Kuntz DA, Rose D (2001) Structure of Golgi α-mannosidase Ⅱ; a target for inhibition of growth and metastasis of cancer cells. The EMBO Journal 20(12): 3008-3017.
-
Meyer TJ, Ramlall J, Thu P, Gadura N (2015) Antimicrobial Properties of Copper in Gram-Negative and Gram-Positive Bacteria, International Scholarly and Scientific Research & Innovation 9(3): 274-278.
-
Tsuneo Ishida (2014) Antibacterial activities of Cu2+ion solution against Gram-negative bacteria. J copper and Copper Alloy 53: 272-278.
-
Tsuneo Ishida (2015) Antibacterial susceptibility tests of Cu2+solution and bacteriolysis and killing action of peptidoglycan cell wall. Chemistry and Industry 66: 611-617.
-
Silhavy TJ, Kahne D, Walker S (2010) The Bacterial Cell Envelope. Cold Spring Harbor Perspectives in Biology 2: a000414.
-
Crichton RT, Shioya M (2016) Japanese Translation, Biological Inorganic Chemistry. Tokyo Kagaku-Dojin Limited 175-188.
-
Egan AJF, Biboy J, Veer IV, Breukink E, Vollmer W (2015) Activities and regulation of peptidoglycan synthases. Philosophical Transactions B 370(1679).
-
Zoll S, Patzold B, Schlag M, Gotz F, Kalbacher H, et al. (2010) Structural Basis of Cell Wall Cleavage by a Staphylococcal Autolysin. PloS Pathogens 6(3): 1-13.
-
Humann J, Lenz LL (2009) Bacterial peptidoglycan degrading enzymes and their impact on host muropeptido detection. J innate Immun 1: 88-97.
-
Tsuneo Ishida (2015) Antibacterial activity action of copper ion and copper complex against bacteria. J Materials Life Society 27(1): 23-32.
-
Atilano ML, Pereira PM, Yates J, Reed P, Veiga H, et al. (2010) Teichoic acid are temporal and spatial regulators of peptidoglycan cross-liking _S.aureus._ PNAS 107(44): 18991-18996.
-
Ramadurai L, Lockwood KJ, Nadakavukarenet MJ, Jayaswal RK (1999) Characterization of a chromosomally encoded glycyglycine endopeptidase of _S.aureus_. Microbiology 145: 801-808.
-
Heidrich C, Ursinus A, Berger J, Schwarz H, Holtje JV (2002) Effects of multiple deletions of murein hydrolases on viability, septum cleavage, and sensitivity to large toxic molecules in _E.coli_. J Bacteriology 184(22): 6093-6099.
-
Heijenoot JV (2011) Peptidoglycan Hydrolases of _E.coli_. Microbiology and Molecular Biology Reviews 75(4): 636-663.
-
Bai W, Zhao K, Asami K (2007) Effects of copper on dielectronic properties of _E.coli_ cells. Colloids and Surfaces B: Biointerfaces 58: 105-115.
-
Ramachandran V, Chandrakala B, Kumar VP, Usha V, Solapure SM, et al. (2006) Screen for inhibitors of the coupled transglycosylase-transpeptidase of peptidoglycan biosynthesis in _E.coli_. Antimicrob Agents Chemother 50(4): 1425-1432.
-
Bernadsky G, Beveridge TJ, Clarke AJ (1994) Analysis of the Sodium Dodecyl Sulfate-Stable Peptidoglycan Autolysins of Select Gram-Negative Pathogens by Using Renaturing Polyacrylamide Gel Electrophoresis. J Bacteriology 176(17): 5225-5232.
-
Kehrer JP (2000) The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 149: 43-50.
-
Krzysztof Barbusiński (2009) Fenton reaction- controversy concerning the chemistry. Ecological chemistry and engineerings 16(3): 347-358.
-
Tkeshelashvili LK, McBride T, Spence K, Loeb LA (1992) Mutation spectrum of copper-induced DNA damage. Journal of Biological Chemistry 267(19): 13778.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions