A Holistic Review on Chloroquine/Hydroxychloroquine the Anti- Parasitic Therapeutics
Hydroxychloroquine and chloroquine have driven attention as potential anti-parasitic therapeutics with extended influences against other various medical conditions. Recently, these drugs were presented against the pandemic of COVID-19. In addition, there is a growing data concerning the resistance to the traditional anti-parasitic regimens. Here, we introduce a general descriptive review to demonstrate (1) a rapid historical view for these drugs, (2) their pharmacokinetics, (3) mode of action and how this affected malaria and whether it can applied in other parasitic diseases or not, and (4) their medical manipulations in other medical conditions involving COVID-19 in the top of the list. The review also tried to introduce their limitations of usage in accordance to previous in vitro and clinical settings. We call for further development in their formula for less toxic and more effective harmaceutical versions.
Introduction
Hydroxychloroquine (HCQ) (brand name Plaquenil) and chloroquine (CQ) are 4-aminoquinolines antimalarial agents, which has recently been regarded as an advantageous therapy in a wide scale of diseases other than malaria. Interestingly, quinine was attributed conventionally to the descendants of the Incan people in Peru dating back to 1630. Also, it worth to mention that they had been used as effective therapeutics since 1894 for cutaneous lupus. During the second world war, chloroquine was tested in the USA as a large series of alkaloids constituted of 4-aminoquinolines with prophylactic effects against malaria and collateral improvement of skin rashes and inflammatory arthritis. However, Germans rejected its usage owing to its toxicity in the avian experimental models [1, 2].
Parasitic diseases cause continual morbidity
predominantly in tropical areas. Available therapeutic anti- parasitic drugs are far from being optimal in spite of being introduced many past decades. However, the financial forces of the market are insufficient to support the development of new drugs. By the new millennium, the global investment presented only 0.1% of the total drug research for tropical diseases that subsidize about 5% of the world-wide disease burden.
Many parasites tempt to show drug resistance against their common anti-parasitic compounds. At the same time, HCQ/CQ are of well-known dynamics and are cost- effective [3, 4, 5]. So that in this review we tempt to display the pharmacokinetics and the pharmaceutical actions of HCQ and CQ interchangeable; and how can it be beneficial to different protozoal infections? Also, we extended in this review to show whether HCQ/CQ can be adventitious in other medical conditions or not? And to which extend is their safety?
Pharmacokinetics
HCQ is water soluble with well achieved absorption at pH (7.0-8.0) when being administrated orally. Severe malnutrition for instance kwashiorkor affects absorption while increased intestinal motion does not. The peak plasma level occurs in a time interval of 4–12 h after a single dose. A stable plasma level occurs after 4–6 weeks of steady daily dose, yet the interindividual variations is significant. Its bioavailability is 75% and its half-life is prolonged up to 40 - 50 days, with low blood clearance (96 ml/min) [6].
In regards to its distributive pattern, HCQ and its metabolites is characterized by its affinity to the pigmented tissues, the mononuclear cells besides other tissues including myocytes. The plasma concentrations of HCQ is influenced by its robust affinity for a lot of blood constituents, involving granulocytes, thrombocytes in addition to the plasma proteins; α-acid glycoprotein and albumin. In rheumatoid arthritis (RA) patients, blood concentrations vary widely up to an eleven-fold in similar doses as part of the drug is held on the platelets which increases dramatically in this inflammatory disease. Also, in the chronic use, metabolites of HCQ involving des-ethyl-hydroxychloroquine and bi-des- ethyl-hydroxychloroquine are noted to accumulate, and to bind to several body tissues, this subsequently affects the total plasma level of the drug and its excretion [7, 8, 9, 10].
High concentrations increase liability to drug toxicity of HCQ that can be manifested in the form of EKG abnormalities. Also, it induces profound hypoglycaemia, hence it is mandatory to monitor blood glucose levels in diabetic patients. HCQ may also increase the serum insulin level in prediabetics who are at high risk of developing diabetes mellitus. Studies concerning drug-drug interactions in spite of being limited their potential kinetic interactions for cimetidine and d-penicillamine were documented while being speculated safe for ranitidine, aspirin, and imipramine. CQ may reduce the hepatotoxicity of methotrexate as it contributes to the reduction of its bioavailability. HCQ inhibits biotransformation of metoprolol (β-blocker) by CYP2D6 [11, 12, 13, 14, 15].
Excretion of this drug is mainly renal where 40-50% is stereo-selective while 20 to 47% pass in urine unchanged. Since HCQ is a weak base, urine acidification enhances its excretion. Minor excretion occurs only through the secretory pathway in the sweat and saliva or bile. Concerning drug efficacy, it worth to mention that the two pharmaceutical forms, chloroquine (CQ) and hydroxychloroquine (HCQ) are of equal efficacy against malignant malaria; nevertheless, the hydroxyl group in HCQ declines its toxicity [16, 17, 18]. Pharmacological Effects of HCQ Involve Immune Modulatory Effect Innate Immunity The First Activation Signal of Innate Immunity Activation (Figure: 1): HCQ inhibits the signalling of the toll-like receptors (TLR) by blocking TLR-7 and TLR-9 in the dendritic cells. In addition, it interferes with the lysosomal acidification and thus inhibit proteolysis besides chemotaxis, endosomal phagocytosis, antigen processing, assembly of MHC α and β chains, and MHC recycling such that only MHC complexes with high affinity are accessible to the cell surface. Moreover, HCQ hinders TLR9-mediated B cell functions [19, 20, 21, 22, 23].
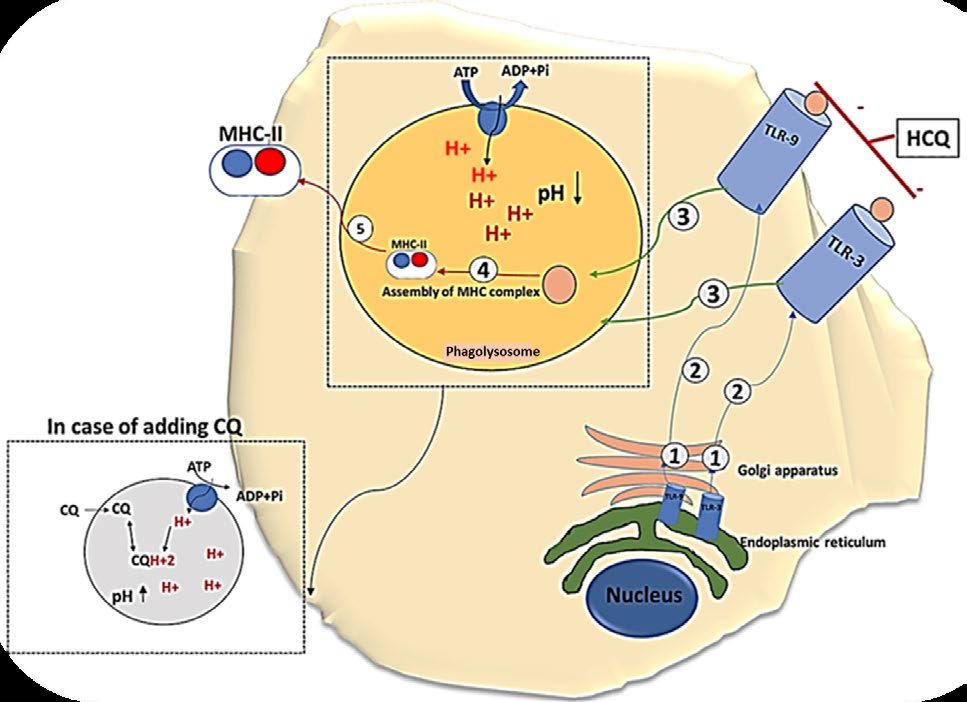
Figure 1: The first signal of dendritic cell activation. The signal starts by the synthesis of TLR in the endoplasmic reticulum to be modified and encased in Golgi apparatus (1) and then it is transported to the cell membrane as a spanning protein (2) to be activated by the recognition of the unique molecular configuration of the foreign antigens. Subsequently, the foreign antigens will be processed in the phagolysosome (3) and assembled with the MHC-II (4) to be presentable to the second line of immune defence (5). The diagram illustrated and clarified the text. Drawn by dr. Enas A. Elsaftawy.
The Second Signal of Innate Immunity Activation (Figure 2): HCQ has been regarded as an ion channel inhibitory agent. Electrophysiological studies speculated that HCQ blocks Ca++-activated K+ conductance in macrophages which is ATP dependent. Subsequently, K+ efflux, inflammasome, IL-1β and IL-18 secretion, and NO production are all inhibited. In this context, HCQ was found to inhibit interferon-α, a crucial cytokine in SLE pathogenesis, IL-1 and tumour necrosis factor. HCQ also antagonize the effect of the prostaglandins by inhibiting phospholipase A2 and the matrix metalloproteinase enzymes [24, 25, 26, 27, 28].
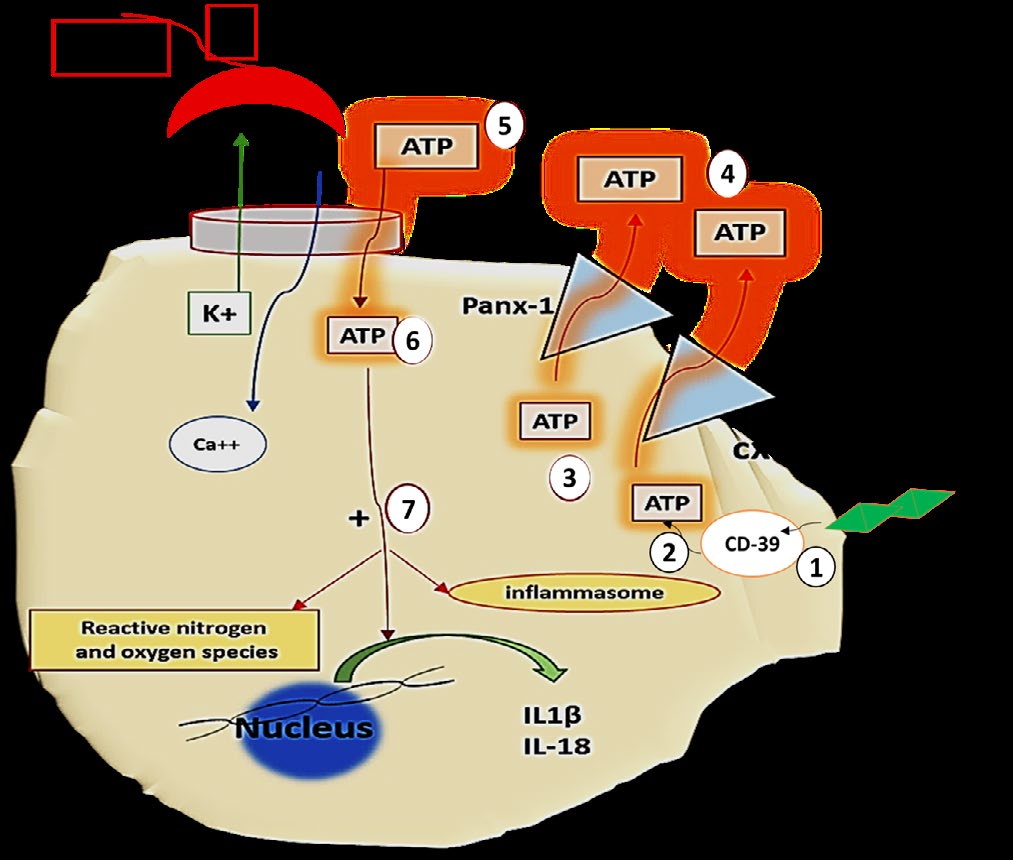
Figure 2: The second signal of dendritic cells activation. TLR induce internaliztion of CD39 (1) that consequently leads to increased intracellular ATP accumulation (2) to be subsequently released by Panx-1 and cx43 channels (3). Bioavailability of extracellular ATP (4) is mandatory for the activation of the P2x7 channels (proinflammatory Ca++-dependent K+ channels) (5) with subsequent pro-inflammatory sequels. HCQ is proficient in blocking P2x7 channels and thus suppress inflammation. Note the diagram illustrates the above text. Drawn by dr. Enas A. Elsaftawy.
Adaptive Immune Response
- Inhibition of T and B-cell receptors calcium signalling.
- Inhibition of antigen-antibody reaction. T cells Resting lymphocytes maintain a low concentration of Ca2+. However, engagement of antigen receptors induces calcium influx from the extracellular space by several routes as illustrated in figure (3), where calcium acts as a second messenger fundamental for activation, differentiation of the naive- T-cells and effector functions of mature T cells. Strikingly, HCQ exerts anti-TCR-crosslinking that hinders calcium mobilization, in a concentration dependent manner. Pharmaceutical actions involve primary T-cells and mature T-cell lines.
• In the proximal events: TCR-mediated intracellular calcium mobilization was inhibited by HCQ in a dose- dependent fashion. Meanwhile, effect of HCQ (1) don’t cover the phosphorylation of the inductive protein tyrosine or tyrosine phospholipase C-Ɣ1 enzyme, or (2) the total production inositol phosphate [21, 25, 29, 30, 31, 32, 33].
• In the distal events: HCQ suppress anti-TCR-induced up-regulation of CD69 expression as induction of CD69 is mandatory for T regulatory cell activation, which is beneficial in treating patients with auto-immune disease [21, 34, 35].
B cells
HCQ suppresses deeply the TLR(9)-mediated B-cell functions during the inflammatory processes: (1) similar to T-cells, HCQ hinders B-cell receptor induced calcium signal, (2) inhibits the differentiation of (CD19+IgD-CD27+ memory B-cells) in to plasma-blasts and thus (3) inhibit their IgG production[36].
![Figure 3: Characterization of Ca2+ channels during TCR signalling. (1,2) Antigen presenting cells activate TCR, (3) hydrolysis of PIP2 into DAG and IP3, (4) IP3 activates IP3 sensitive Ca channel (endoplasmic reticulum), (5) Ca2+ activates protein kinase C, which is also activated by DAG (6). Activated protein kinase C undergoes phosphorylation cascade of the cellular substrates. (7) Ca2+ induce activation of cytokine transcription factors. The diagram illustrated the inhibitory effect of HCQ on T cell activation. Drawn by dr. Enas A. Elsaftawy. Antigen Antibody Complexes: CQ was found to dissociate antigen antibody immune complexes and thus it can reverse the antibodydependent cellmediated cytotoxicity (ADCC) and complementdependent cytotoxicity (CDC) in a unique reaction that was not seen by other immune modulate agents for instance methotrexate [37,38].](/fulltextimages/5830/fig_3.jpeg)
Figure 3: Characterization of Ca2+ channels during TCR signalling. (1,2) Antigen presenting cells activate TCR, (3) hydrolysis of PIP2 into DAG and IP3, (4) IP3 activates IP3 sensitive Ca channel (endoplasmic reticulum), (5) Ca2+ activates protein kinase C, which is also activated by DAG (6). Activated protein kinase C undergoes phosphorylation cascade of the cellular substrates. (7) Ca2+ induce activation of cytokine transcription factors. The diagram illustrated the inhibitory effect of HCQ on T cell activation. Drawn by dr. Enas A. Elsaftawy. Antigen Antibody Complexes: CQ was found to dissociate antigen antibody immune complexes and thus it can reverse the antibodydependent cellmediated cytotoxicity (ADCC) and complementdependent cytotoxicity (CDC) in a unique reaction that was not seen by other immune modulate agents for instance methotrexate [37, 38].
Anti-Coagulant Effect of Hydroxychloroquine was Demonstrated in Figure (4)
![Figure 4: Impact of HCQ on coagulation system reduces thrombotic evidence [39,40]. Drawn by dr. Enas A. Elsaftawy.](/fulltextimages/5830/fig_4.png)
Hydroxychloroquine in Parasitic Diseases
Sporozoan
Blood Sporozoan: Plasmodium Spp: The antimalarial actions of CQ and its analogues is attributed to (1) its lysosome- tropic properties as it is a weak base so it accumulates (nearly
1000-fold) in the acidic media of the parasitic lysosomes [41], and (2) the potent immunomodulatory actions of these compounds [42]. The process involves the immune recognition and the downstream cascade of inflammation [43, 44].
Administration: By dosing: (1) In CQ: immediate oral administration of 1000 mg is followed by oral 500 mg at hours 6, 24, and 48 hours. (2) While in Hydroxychloroquine: 800 mg immediate oral intake is followed by 400 mg oral doses at hours 6, 24, 48 hours [45, 46]. Action of CQ can be potentiated by amlodipine (calcium channel blocker), as its accumulation increases inside the erythrocytes of the murine models infected with P. falciparum. Also, this combination may be beneficial in the treatment protocol of resistant strains of P. falciparum (in vitro studies) [47]. In resistant strains of P.falciparum HCQ was found to be lesser than CQ in efficacy [48].
Limitations of Use in HCQ/CQ in Treatment of Malaria: Not recommended for treatment of (1) complicated malaria, (2) chloroquine or hydroxychloroquine-resistant strains of Plasmodium species, (3) in geographic areas where chloroquine resistance occurs, and (4) relapses of P. vivax or P. ovale because of being not active against the hypnozoite forms of these parasites [49].
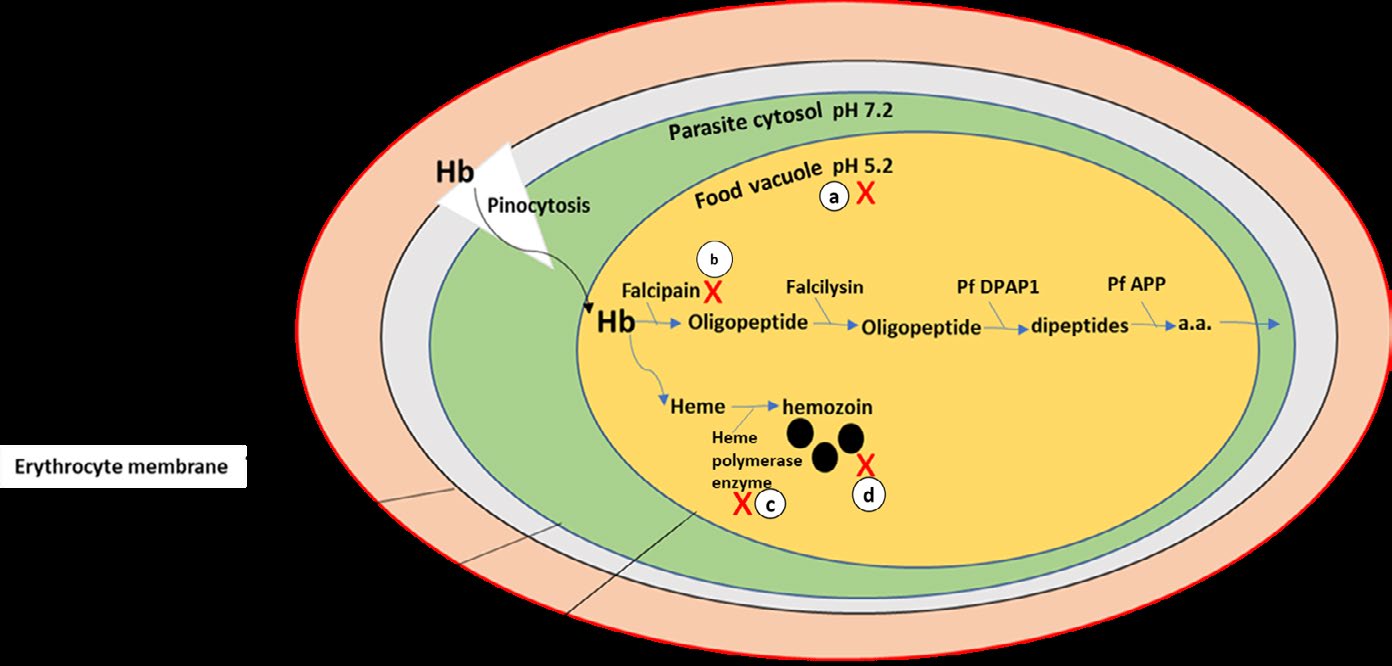
Figure 5: In regards to prior studies, HCQ hinders growth and replication of the merozoite as it (1) accumulates in the food vacuole (lysosome) of the malaria parasite to raise its pH, (2) interferes with the ability of the parasite to proteolyse hemoglobin (Hb) by acting on the protease enzyme called falcipain, (3) interfere with the action of (the parasitic) heme-polymerase enzyme that leads to the accumulation of the toxic by-product (β-hematin). The diagram clarified anti-parasitic actions of HCQ in malaria. Drawn by dr. Enas A. Elsaftawy.
Tissue Sporozoan: Toxoplasma Gondii
Therapeutic studies on this parasite started as early as 1950s on murine models. In 2011, a case report for SLE showed that the immune modulatory effect of HQ activates latent toxoplasmosis. In 2019, chloroquine resistant transporter in bradyzoites was found to aid in the maintenance of the parasite’s viability and its digestive vacuole [50, 51, 52].
Intestinal Sporozoan: Cryptosporidium Parvum
Causes self-limiting diarrhoea in immunocompetent hosts. Several case reports conveyed activation of cryptosporidiosis secondary to the immunosuppressive status caused by HCQ [53].
Flagellate
Intestinal Flagellates: Giardia Lamblia: Despite of being cosmopolitan and nearly 5,000 people are hospitalized due to giardiasis in USA, few therapeutic protocols have been issued. a 5-Nitroimidazole compound is implicated in the treatment of most case; however, emergence of treatment failure, drug resistance in addition, to the drug side effects was introduced [54]. Implication of CQ in giardiasis infection dates back to 1961with encouraging results [55, 56, 57, 58]. However, hydroxy chloroquine despite being less toxic no single report can be found in treatment of giardiasis.
Tissue and Blood Flagellates
Visceral Leishmania (Leishmania Donovani): In one case report published in 2008, a patient suffered of visceral leishmaniasis and SLE was treated with hydroxychloroquine combined with lysine acetylsalicylate, steroids, enalapril and experienced complete remission [59]. Cutaneous Leishmaniasis: Few case reports were published for successful treatment of cutaneous leishmaniasis (spp. L.mexicana) in soldiers deployed in Iraq. In addition, oral chloroquine in murine models infected with (spp. L. amazonensis) showed reduction in the lesion size and low parasite number [60, 61, 62]. Trypanosoma Brucei Brucei And T. Brucei Gambiense: The causative agents of the sleeping sickness. In vivo studies CQ had a sub-curative effect and prolonged the survival period is prolonged. However, HCQ is still naive model for further research [63, 64]. Trypanosoma Cruzi: The causative agent of Chagas disease (CD). An amusing case report reported that Chagas disease was reactivated during treatment with hydroxychloroquine; a finding attributable to the opportunistic nature of T. cruzi and the immunomodulatory effect of the hydroxychloroquine [65, 66]. Amebiasis: Entamoeba Histolytica: In 1975, a comparative therapeutic study documented the priority of metronidazole over hydroxychloroquine in hepatic amebiasis; where the study reported the accelerated rate of healing. Similar result in intestinal amoebiasis was documented [67].
However, Cq/Hcq was Found to be Recommended in the Treatment of Several Infectious and Medical Conditions other than Parasitic Disease
Viral Infections
Pandemic of Coronavirus Disease 2019 (COVID-19): It seems to show good prognosis. At first COVID-19 was described as severe pandemic acute respiratory syndrome, however, shortly after, COVID -19 was clarified not to be any form of pneumonia or ARDs. A predictable pathway for the initial events following the viral invasion to the type-II pneumocytes is illustrated in Figure (6). SARS2 Corona Virus starves body of oxygen by binding to heme group (porphyry) in the hemoglobin in a certain way that releases the iron ion into the circulation. Mode of action of CQ in COVID-19 was guided by that played in malaria infection, where it protects against Hb invasion [68]. Cytokine Storm: in the form of IL1, IL6, and TNF-α, besides the inflammatory biomarkers (ESR and C-reactive protein) elevates dramatically. In this regard CQ seems to be beneficial being anti-inflammatory [69, 70]. Ferritin Levels: Increase dramatically due to increased iron release in the blood. However, cellular mobilization of iron from transferrin and ferritin depends on the acidic media and proteolytic degradation in the lysosomal like compartment. CQ acts by raising the pH therefore it (1) depresses ferritin and iron uptake by hepatocytes, (2) interferes with intracellular free iron availability and iron remains bound to transferrin, and (3) hinders the recycling of the ferritin receptors [71, 72, 73, 74, 75]. Lymphopenia: COVID-19 was found to predilect CBC profile to lymphopenia not lymphocytosis and instead WBCs differentiation is favoured towards monocytes to engulf the excess iron load [75]. However, these drugs are supposed to interfere with T-cell activation for instance the major histocompatibility complex (MHC) class-II-antigen presentation, besides the intracellular calcium signalling [21]. D-Dimer: Due to activated coagulation, excessive consumption of coagulation factors, and induced fibrinolytic pathways. Also, macrophage activation like syndrome seemed to induce severe intra-pulmonary vascular inflammation that involve micro-thrombosis and haemorrhages. This results in pulmonary intravascular coagulopathy (PIC) rather than the DIC. Anticoagulants are generally advised in DIC; nevertheless, there is still a debate regarding the benefit of CQ in spite of having anti-coagulant properties [77, 78, 79, 80, 81].
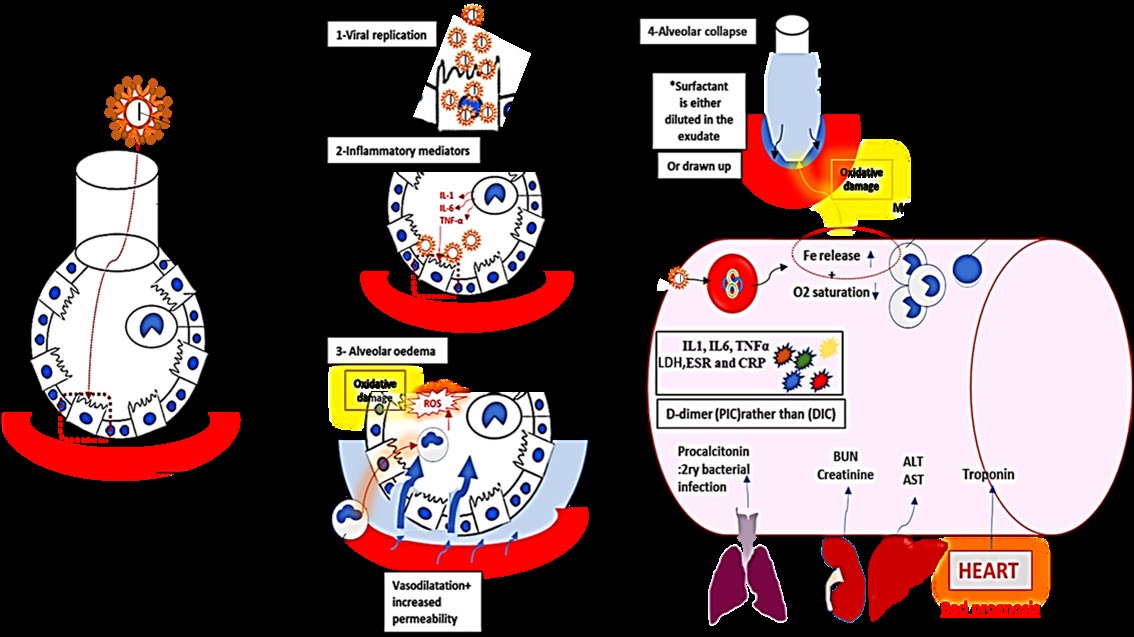
Figure 6: Pulmonary haemophagocytosis has been reported in COVID-19 with severe extensive alveolar and interstitial inflammation in the juxtaposed pulmonary vasculature. The diagram shows also the eventually O2 desaturation and release in the blood in addition to the powerful oxidative damage occurs in the lungs by the released iron. The associated cytokine storm, D-dimer, elevated liver and kidney functions were all illustrated in the figure.
HIV: HCQ treatment was correlated with (1) optimal suppression of HIV replication, (2) decrease in plasma lipopolysaccharide level; (3) decline in count of TLR4- expressing CD14+ cells, (4) reduction in IFNα and IL6 (5) increase in naive Tregs and TLR4-expressing Tregs, and (6) increase in CD4+ T cells count [82]. Bacterial Infection: the combination of CQ and doxycycline in chronic Q-fever endocarditis caused by Coxiella burnetii is recommended for18–36 months [83]. Autoimmune Diseases: Hydroxychloroquine is the keystone of the management of SLE (1) as it possesses antithrombotic properties, figure (4), (2) modulates blood sugars, (3) improves lipids, and (4) the overall autoimmune disease activity [84, 85, 86]. Cardiovascular Diseases: HCQ is bradycardic agents suggested in the treatment of ischemic heart disease and heart failure to minimize myocardial oxygen consumption. HCQ exert direct inhibitory effect on SA node through multichannel inhibition besides its anti-thrombotic actions [87]. Renal Diseases: After adjusting for possible confounding factors the protective effect of hydroxychloroquine in retarding renal damage and CKD occurrence in SLE and rheumatoid diseases is still evident. However, clinical trials speculated that sudden withdrawal of HCQ in SLE can lead to flare up of the disease including the life-threatening lupus nephritis [88, 89]. Organ Transplantation: suggested to treat the graft-versus- host disease being evaluated in the bone marrow transplant [90, 91]. Oncology: over the past 2–3 decades being of prominent effects in promoting autophagy and apoptosis processes in malignant cells [92]. Metabolic Diseases: HCQ appears to protect against the occurrence of diabetes, and dyslipidemia [93, 94].
Safety of Hydroxychloroquine
CHQ and HCQ compounds have revealed excellent safety profile with a virtuous long-term tolerance. 4-aminoquinolines are recorded on World Health Organization’s List of Essential Medicines as the safest and most potent medicines essential in a health system. In 1955 medical use of Hydroxychloroquine was approved in the USA; and in 2017, it was recorded the 128th most frequently prescribed medical treatment for more than five million prescriptions. Nevertheless, HCQ is better than CQ in regards to the clinical efficacy and the safety profile not only in the general population but also among special cases including pregnancy and those suffer from renal failure. In this accordance, it remains one of the available therapeutic regimens in pregnancy. On the other hand, it noteworthy to mention the debate about these anti- malarial drugs in pregnancy being accused for the possible link with renal agenesis and spina bifida in some cases. CHQ and HCQ have never been proved to be carcinogenic, in spite of binding to DNA [95]. Common Side Effects: vomiting is the most common side effect besides other manifestation in the form of headache, blurring vision, and myopathy, allergic reactions. Ocular Side Effects: in the form of (1) corneal deposits, (2) opacity of the posterior subcapsular lens, (3) dysfunction of the ciliary body, (4) and most important, irregularity in the macular pigmentation in the early phase, a ring of macular pigment dropout in the advanced stage, and (5) peripheral bone spicule formation, (6) vascular attenuation, and (7) optic disc pallor (end-stage) [96, 97, 98]. Accordingly, the guidelines recommend safe dosing as the risk of retinopathy is less than 2% at a dose 4–5 mg/kg/day [100]. Diagnosis of Visual Field Defects: The visual computerized campimetry (visual field test): paracentral scotomas are very common in retinal toxicity due to CQ and HCQ. However, the correlation between the findings of the visual field test and the ocular lesions in the fundus are not usually equal [101, 102].
The optical coherence tomography (OCT): which is an image modality that allows the early detection of subtle changes signing for toxic retinopathy using high-resolution and cross-sectional tomographic. Image findings were found to be in the form of (1) direct binding to the melanin in the retinal pigmented epithelium (RPE)thickening of the retinal pigment epithelium, (2) thinning of the outer segments of the foveal photoreceptor and blunted foveal reflex (3) direct toxicity of the macular ganglion cell–inner plexiform layer, (4) reduction in the central foveal thickness (deteriorating prognosis), and (5) bull’s-eye maculopathy which is a ring- shaped pattern of damage around the macula [103, 104, 105]. G6pd Glucose-6-Phosphate Dehydrogenase (G6PDH)- Deficiency: G6PDH is pivotal to reduce oxidative stress, where its deficiency leads to excessive accumulation of free radicals and the eventual haemolysis in RBCs. Severity of RBCs haemolysis relays chiefly on the genotype of G6PD, which is different in various geographic areas where the Mediterranean variant is the most severe. Although the American College of Rheumatology recommend against the routine G6PD testing for prior to initiation HCQ [106, 107]. Skin: hyperpigmentation related to HCQ, appears to be due to local bruising following deposition of iron in the soft tissue; however, the exact reason still unclear [108, 109]. Musculoskeletal: HCQ-related skeletal myopathy is an uncommon adverse effect. The condition improves with discontinuation of the treatment in few weeks due to prolonged elimination half-life [110]. Psychological Changes: There is minimal evidence for HCQ- induced psychosis and the condition requires the presence of several predisposing risk factors e.g. familial history of psychiatric diseases, drug interactions or low doses of glucocorticoid, alcohol intake, and in female gender [111].
Conclusion
Hydroxychloroquine and chloroquine are considered as cheap and well-studied medical formulas that need more concern and development to achieve more benefits with less toxic results. These drugs seem to affect pathogens with certain invasive phenomena to the red blood corpuscles in specific e.g. malaria and COVID-19. Also, owing to their extended mode of action they appeared to be beneficial in other therapeutic protocols concerning diseases of specific immune properties.
References
-
Woodward R, Doering W (1944) The total synthesis of quinine. J Am Chem Soc 66: 849.
-
Iredale J, Fieger H, Wainer IW (1993) Determination of the stereoisomers of hydroxychloroquine and its major metabolites in plasma and urine following a single oral administration of racemic hydroxychloroquine. Semin Arthritis Rheum 23(2 Suppl 1): 74-81.
-
Furst DE (1996) Pharmacokinetics of hydroxychloroquine and chloroquine during treatment of rheumatic diseases. Lupus 1: 11-15.
-
Hastings IM (2001) Modelling parasite drug resistance: lessons for management and control strategies. Tropical Medicine & International Health 6(11): 883-890.
-
Pink R, Hudson A, Mouries MA, Bendig M (2005) Opportunities and challenges in antiparasitic drug discovery. Nature reviews Drug discovery 4(9): 727-740.
-
Costedoat Chalumeau N, Dunogué B, Morel N, Le Guern V, Guettrot Imbert G (2014) Hydroxychloroquine: a multifaceted treatment in lupus. La Presse Médicale 43(6): e167-180.
-
Tett SE, Cutler DJ, Day RO, Brown KF (1989) Bioavailability of hydroxychloroquine tablets in healthy volunteers. British journal of clinical pharmacology 27(6): 771-779.
-
McLachlan AJ, Tett SE, Cutler DJ, R O Day (1994) Bioavailability of hydroxychloroquine tablets in patients with rheumatoid arthritis. Br J Rheumatol 33(3): 235- 239.
-
Ono C, Yamada M, Tanaka M (2003) Absorption, distribution and excretion of 14C-chloroquine after single oral administration in albino and pigmented rats: binding characteristics of chloroquine-related radioactivity to melanin in vivo. J Pharm Pharmacol 55(12): 1647-1654.
-
Tanaka E, Taniguchi A, Urano W, Yamanaka H, Kamatani N (2004) Pharmacogenetics of disease-modifying anti- rheumatic drugs. Best Pract Res Clin Rheumatol 18(2): 233-247.
-
Carmichael SJ, Charles B, Tett SE (2003) Population pharmacokinetics of hydroxychloroquine in patients with rheumatoid arthritis. Therapeutic drug monitoring 25(6): 671-681.
-
Augustijns P, Verbeke N (1993) Stereoselective pharmacokinetic properties of chloroquine and de- ethyl-chloroquine in humans. Clinical pharmacokinetics 24(3): 259-269.
-
Sheikhbahaie F, Amini M, Gharipour M, Aminoroaya A, Taheri N (2016) The effect of hydroxychloroquine on glucose control and insulin resistance in the prediabetes condition. Advanced biomedical research 5: 145.
-
Tett SE, McLachlan AJ, Cutler DJ, Day RO (1994) Pharmacokinetics and pharmacodynamics of hydroxychloroquine enantiomers in patients with rheumatoid arthritis receiving multiple doses of racemate. Chirality 6(4): 355-359.
-
Somer M, Kallio J, Pesonen U, Pyykkö K, Huupponen R, et al. (2000) Influence of hydroxychloroquine on the bioavailability of oral metoprolol. British journal of clinical pharmacology 49(6): 549-554.
-
Emami J, Pasutto FM, Jamali F (1998) Effect of experimental diabetes mellitus and arthritis on the pharmacokinetics of hydroxychloroquine enantiomers in rats. Pharmaceutical research 15(6): 897-903.
-
Henriksen AM (2015) Quantitative salivary gland scintigraphy in the evaluation of patients with sicca symptoms (Master’s thesis, Universitetet i Tromsø).
-
Bothwell B, Furst DE (2005) Hydroxychloroquine. Antirheumatic Therapy Actions and Outcomes, pp: 81- 92.
-
Kužnik A, Benčina M, Svajger U, Jeras M, Rozman B, et al. (2011) Mechanism of endosomal TLR inhibition by antimalarial drugs and imidazoquinolines. The Journal of Immunology 186(8): 4794-804.
-
Miyake K, Shibata T, Ohto U, Shimizu T, Saitoh SI, et al. (2018) Mechanisms controlling nucleic acid-sensing Toll-like receptors. International immunology 30(2): 43- 51.
-
Goldman FD, Gilman AL, Hollenback C, Kato RM, Premack BA, et al. (2000) Hydroxychloroquine inhibits calcium signals in T cells: a new mechanism to explain its immunomodulatory properties. Blood, The Journal of the American Society of Hematology 95(11): 3460-3466.
-
van Loosdregt J, Spreafico R, Rossetti M, Prakken BJ, Lotz M, et al. (2013) Hydroxychloroquine preferentially induces apoptosis of CD45RO+ effector T cells by inhibiting autophagy: a possible mechanism for therapeutic modulation of T cells. Journal of Allergy and Clinical Immunology 131(5): 1443-1446.
-
Qiao L, Zhang J (2009) Inhibition of lysosomal functions reduces proteasomal activity. Neuroscience letters 456(1): 15-19.
-
Schroeder ME, Russo S, Costa C, Hori J, Tiscornia I, et al. (2017) Pro-inflammatory Ca++-activated K+ channels are inhibited by hydroxychloroquine. Scientific reports 7(1): 1-7.
-
Savignac M, Mellstrom B, Naranjo JR (2007) Calcium- dependent transcription of cytokine genes in T lymphocytes. Pflügers Archiv-European Journal of Physiology 454(4): 523-533.
-
Perecko T, Kassab RB, Vasicek O, Pekarova M, Jancinova V, et al. (2014) The effects of chloroquine and hydroxychloroquine on nitric oxide production in RAW 264.7 and bone marrow-derived macrophages. Folia biologica 60(S1): 39-44.
-
Cohen HB, Briggs KT, Marino JP, Ravid K, Robson SC, et al. (2013) TLR stimulation initiates a CD39-based autoregulatory mechanism that limits macrophage inflammatory responses. Blood, The Journal of the American Society of Hematology 122(11): 1935-45.
-
Lohman AW, Isakson BE (2014) Differentiating connexin hemichannels and pannexin channels in cellular ATP release. FEBS letters 588(8): 1379-1388.
-
Sperber K, Quraishi HU, Kalb TH, Panja AS, Stecher V, et al. (1993) Selective regulation of cytokine secretion by hydroxychloroquine: inhibition of interleukin 1 alpha (IL-1-alpha) and IL-6 in human monocytes and T cells. The Journal of Rheumatology 20(5): 803-808.
-
Bolanos‐Meade J, Zhou L, Hoke A, Corse A, Vogelsang G, et al. (2005) Hydroxychloroquine causes severe vacuolar myopathy in a patient with chronic graft‐versus‐host disease. American journal of hematology 78(4):306-9.
-
Fox RI (1993) Mechanism of action of hydroxychloroquine as an antirheumatic drug. InSeminars in arthritis and rheumatism 23(2): 82-91.
-
Xu JC, Peng YB, Wei MY, Wu YF, Guo D, et al. (2015) Chloroquine inhibits Ca2+ signaling in murine CD4+ thymocytes. Cellular Physiology and Biochemistry 36(1): 133-140.
-
Wallace DJ, Gudsoorkar VS, Weisman MH, Venuturupalli SR (2012) New insights into mechanisms of therapeutic effects of antimalarial agents in SLE. Nature Reviews Rheumatology 8(9): 522-533.
-
Schmidt RL, Jutz S, Goldhahn K, Witzeneder N, Gerner MC, et al. (2017) Chloroquine inhibits human CD4+ T-cell activation by AP-1 signaling modulation. Scientific Reports 7(1): 1-4.
-
Yu L, Yang F, Zhang F, Guo D, Li L, et al. (2018) CD69 enhances immunosuppressive function of regulatory T-cells and attenuates colitis by prompting IL-10 production. Cell death & disease 9(9): 1-4.
-
Torigoe M, Sakata K, Ishii A, Iwata S, Nakayamada S, et al. (2018) Hydroxychloroquine efficiently suppresses inflammatory responses of human class-switched memory B cells via Toll-like receptor 9 inhibition. Clinical Immunology 195: 1-7.
-
Li DG, Hu WZ, Ma HJ, Liu W, Yang QQ, et al. (2016) Hydroxychloroquine protects melanocytes from autoantibody-induced injury by reducing the binding of antigen-antibody complexes. Molecular medicine reports 14(2): 1275-1282.
-
Segal Eiras A, Segura GM, Babini JC, Arturi AS, Fraguela JM, et al. (1985) Effect of antimalarial treatment on circulating immune complexes in rheumatoid arthritis. The Journal of rheumatology. 12(1): 87-89.
-
Miranda S, Billoir P, Damian L, Thiebaut PA, Schapman D, et al. (2019) Hydroxychloroquine reverses the prothrombotic state in a mouse model of antiphospholipid syndrome: Role of reduced inflammation and endothelial dysfunction. PloS one 14(3): e0212614.
-
Sciascia S, Branch DW, Levy RA, Middeldorp S, Pavord S, et al. (2016) The efficacy of hydroxychloroquine in altering pregnancy outcome in women with antiphospholipid antibodies. Thrombosis and haemostasis 115(02): 285- 290.
-
Harhaji Trajkovic L, Arsikin K, Kravic Stevovic T, Petricevic S, Tovilovic G, et al. (2012) Chloroquine-mediated lysosomal dysfunction enhances the anticancer effect of nutrient deprivation. Pharmaceutical research 29(8): 2249-2263.
-
Zhu X, Pan Y, Li Y, Jiang Y, Shang H, et al. (2012) Targeting Toll-like receptors by chloroquine protects mice from experimental cerebral malaria. International immunopharmacology 13(4): 392-397.
-
Lim HS, Im JS, Cho JY, Bae KS, Klein TA, et al. (2009) Pharmacokinetics of hydroxychloroquine and its clinical implications in chemoprophylaxis against malaria caused by Plasmodium vivax. Antimicrobial agents and chemotherapy 53(4): 1468-1475.
-
Baradaran Eftekhari R, Maghsoudnia N, Dorkoosh FA (2020) Chloroquine: A brand-new scenario for an old drug. EODD 17(3): 1.
-
Pathak DP, Sharma V, Kumar S (2018) Designing novel inhibitors against falcipain-2 of Plasmodium falciparum. Bioorganic & medicinal chemistry letters 28(9): 1566- 1569.
-
Pinheiro LCS, Feitosa LM, Silveira FFD, Boechat N (2018) Current Antimalarial Therapies and Advances in the Development of Semi-Synthetic Artemisinin Derivatives. An Acad Bras Cienc 90(1 Suppl 2): 1251-1271.
-
Deloron P, Basco LK, Dubois B, Gaudin C, Clavier F, et al. (1991) In vitro and in vivo potentiation of chloroquine against malaria parasites by an enantiomer of amlodipine. Antimicrobial agents and chemotherapy 35(7): 1338-1342.
-
Warhurst DC, Steele JC, Adagu IS, Craig JC, Cullander C (2003) Hydroxychloroquine is much less active than chloroquine against chloroquine-resistant Plasmodium falciparum, in agreement with its physicochemical properties. Journal of Antimicrobial Chemotherapy 52(2): 188-193.
-
White NJ, Krishna S (1989) Treatment of malaria: some considerations and limitations of the current methods of assessment. Transactions of the Royal Society of tropical medicine and hygiene 83(6): 767-777.
-
Kannan G, Di Cristina M, Schultz AJ, Huynh MH, Wang F, et al. (2019) Role of Toxoplasma gondii Chloroquine Resistance Transporter in Bradyzoite Viability and Digestive Vacuole Maintenance. mBio 10(4): e01324-19.
-
Pagalavan L, Kan FK (2011) Cerebral toxoplasmosis in systemic lupus erythematosus following intravenous methylprednisolone. Med J Malaysia 66(1): 68-70.
-
McCann (2019) Cryptosporidium Enteritis in an Immunosuppressed Child: Applying the Adult HIV Treatment Regimen. American Academy of Pediatrics 144(2): 592.
-
Lalle M, Hanevik K (2018) Treatment-refractory giardiasis: challenges and solutions. Infection and drug resistance 11: 1921-1933.
-
Kurban S, Rozenbojm J, Rozenbojm P (1961) Treatment of giardiasis with hydroxychloroquine (Win. 1.258-2). Revista do Hospital das Clinicas 16: 233-240.
-
Escobedo AA, Nunez FA, Moreira I, Vega E, Pareja A, et al. (2003) Chloroquine and albendazole in the treatment of paediatric patients with giardiasis. Ann Trop Med Parasitol 97(4): 367-371.
-
Cañete R, Ribas D, Escobedo AA, González ME, Almirall P, et al. (2010) A randomized, controlled, openlabel trial evaluating the efficacy and safety of chloroquine in the treatment of giardiasis in children. West Indian Med J 59(6): 607-611.
-
Sowunmi A, Walker O, Salako LA (1989) Pruritus and antimalarial drugs in Africans. Lancet 2(8656): 213.
-
Celant A, Chichino G, Dal Bello B, Massarotti M, Minoli L, et al. (2008) Visceral leishmaniasis, systemic lupus erythematosus and acute hepatitis. Rheumatology international 29(1): 111-112.
-
Pehoushek JF, Quinn DM, Crum WP (2004) Cutaneous leishmaniasis in soldiers returning from deployment to Iraq. Journal of the American Academy of Dermatology 51(5): S197-200.
-
Knobloch J, Demar M (1997) Accidental Leishmania mexicana infection in an immunosuppressed laboratory technician. Tropical Medicine & International Health 2(12): 1152-1155.
-
Rocha VP, Nonato FR, Guimaraes ET, Freitas LA, Soares MB (2013) Activity of antimalarial drugs in vitro and in a murine model of cutaneous leishmaniasis. Society for General Microbiology 62(7): 1001-1010.
-
Ogunbanwo JA, Agbonlahor DE, Adamu A, Dalyop P, Elesha SO, et al. (2001) Effects of anti-protozoal drugs and histopathological studies on trypanosome species. FEMS Immunology & Medical Microbiology 30(1): 73- 83.
-
Ringer A (1752) Hydrocortisone/hydroxychloroquine/ prednisone. Reactions 187(11): 2019.
-
Brener Z, Gazzinelli RT (1997) Immnunological Control of Trypanosoma cruzi infection and pathogenesis of Chagas disease. International archives of allergy and immunology 114(2): 103-110.
-
Pinazo MJ, Espinosa G, Gállego M, López-Chejade PL, Urbina JA, et al. (2010) Successful treatment with posaconazole of a patient with chronic Chagas disease and systemic lupus erythematosus. The American journal of tropical medicine and hygiene 82(4): 583-587.
-
Cohen HG, Reynolds TB (1975) Comparison of metronidazole and chloroquine for the treatment of amoebic liver abscess: A controlled trial. Gastroenterology 69(1): 35-41.
-
Gattinoni L, Coppola S, Cressoni M, Busana M, Rossi S, et al. (2020) Covid-19 does not lead to a “typical” acute respiratory distress syndrome. American journal of respiratory and critical care medicine 201(10): 1-2.
-
Ye Q, Wang B, Mao J (2020) Cytokine Storm in COVID-19 and Treatment. Journal of Infection.
-
Karres I, Kremer JP, Dietl I, Steckholzer U, Jochum M, et al. (1998) Chloroquine inhibits proinflammatory cytokine release into human whole blood. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 274(4): 1058-1064.
-
Sibille JC, Kondo H, Aisen P (1989) Uptake of ferritin and iron bound to ferritin by rat hepatocytes: modulation by apotransferrin, iron chelators and chloroquine. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1010(2): 204-209.
-
Ramm GA, Powell LW, Halliday JW (1994) Pathways of intracellular trafficking and release of ferritin by the liver in vivo: the effect of chloroquine and cytochalasin D. Hepatology 19(2): 504-513.
-
Starke PE, Gilbertson JD, Farber JL (1985) Lysosomal origin of the ferric iron required for cell killing by hydrogen peroxide. Biochem Biophys Res Commun 133(2): 371-379.
-
Newman SL, Gootee L, Brunner G, Deepe GS (1994) Chloroquine induces human macrophage killing of Histoplasma capsulatum by limiting the availability of intracellular iron and is therapeutic in a murine model of histoplasmosis. J Clin Invest 93(4): 1422-1429.
-
Lerkvaleekul B, Vilaiyuk S (2018) Macrophage activation syndrome: early diagnosis is key. Open access rheumatology: research and reviews 10: 117-128.
-
Tan L, Wang Q, Zhang D, Ding J, Huang Q, et al. (2020) Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study. Signal transduction and targeted therapy 5(1): 1-3.
-
Levi M, Tencate H (1999) Disseminated intravascular coagulation. N Engl J Med 341: 586-592.
-
Dolhnikoff M, Duarte‐Neto AN, de Almeida Monteiro RA, Ferraz da Silva LF, Pierre de Oliveira E, et al. (2020) Pathological evidence of pulmonary thrombotic phenomena in severe COVID‐19. Journal of Thrombosis and Haemostasis.
-
OBE BH, Retter A, McClintock C (2013) Practical guidance for the prevention of thrombosis and management of coagulopathy and disseminated intravascular coagulation of patients infected with COVID-19.
-
Lippi G, Plebani M (2020) Laboratory abnormalities in patients with COVID-2019 infection. Clinical Chemistry and Laboratory Medicine.
-
Alexander V, Zachariah U, Goel A, Kandasamy S, Chacko B, et al. (2020) Low-volume plasma exchange and low-dose steroid to treat secondary hemophagocytic lymphohistiocytosis: A potential treatment for severe COVID-19?. Current Medical Issues 18(2): 77-82.
-
Romanelli F, Smith KM, Hoven AD (2004) Chloroquine and hydroxychloroquine as inhibitors of human immunodeficiency virus (HIV-1) activity. Current pharmaceutical design 10(21): 2643-2648.
-
Raoult D, Houpikian P, Tissot Dupont H, Riss JM, Arditi Djiane J, et al. (1999) Treatment of Q Fever Endocarditis: Comparison of 2 Regimens Containing Doxycycline and Ofloxacin or Hydroxychloroquine. Arch Intern Med 159(2): 167-73.
-
Lee SJ, Silverman E, Bargman JM (2020) The role of antimalarial agents in the treatment of SLE and lupus nephritis. Nature Reviews Nephrology 7(12): 718-729.
-
Wozniacka A, Lesiak A, Narbutt J, McCauliffe DP, Sysa-Jedrzejowska A (2006) Chloroquine treatment influences proinflammatory cytokine levels in systemic lupus erythematosus patients. Lupus 15(5): 268-275.
-
Borba EF, Bonfá EL (2001) Longterm beneficial effect of chloroquine diphosphate on lipoprotein profile in lupus patients with and without steroid therapy. The Journal of rheumatology 28(4): 780-785.
-
Capel RA, Herring N, Kalla M, Yavari A, Mirams GR, et al. (2015) Hydroxychloroquine reduces heart rate by modulating the hyperpolarization-activated current If: Novel electrophysiological insights and therapeutic potential. Heart rhythm 12(10): 2186-2194.
-
Tsakonas E, Joseph L, Esdaile JM, Choquette D, Senécal JL, et al. (1998) A long-term study of hydroxychloroquine withdrawal on exacerbations in systemic lupus erythematosus. Lupus 7(2): 80-85.
-
Pons‐Estel GJ, Alarcón GS, McGwin Jr G, Danila MI, Zhang J, et al. (2009) Protective effect of hydroxychloroquine on renal damage in patients with lupus nephritis: LXV, data from a multiethnic US cohort. Arthritis Care & Research: Official Journal of the American College of Rheumatology 61(6): 830-839.
-
Khoury H, Trinkaus K, Zhang MJ, Adkins D, Brown R, et al. (2003) Hydroxychloroquine for the prevention of acute graft-versus-host disease after unrelated donor transplantation. Biology of Blood and Marrow Transplantation 9(11): 714-721.
-
Fong T, Trinkaus K, Adkins D, Vij R, Devine SM, et al. (2007) A randomized double-blind trial of hydroxychloroquine for the prevention of chronic graft- versus-host disease after allogeneic peripheral blood stem cell transplantation. Biology of Blood and Marrow Transplantation 13(10): 1201-1206.
-
Verbaanderd C, Maes H, Schaaf MB, Sukhatme VP, Pantziarka P, et al. (2017) Repurposing Drugs in Oncology (ReDO)-chloroquine and hydroxychloroquine as anti-cancer agents. Ecancermedical science 11.
-
Emami J, Gerstein HC, Pasutto FM, Jamali F (1999) Insulin-sparing effect of hydroxychloroquine in diabetic rats is concentration dependent. Canadian journal of physiology and pharmacology 77(2): 118-123.
-
Mercer E, Rekedal L, Garg R, Lu B, Massarotti EM, et al. (2012) Hydroxychloroquine improves insulin sensitivity in obese non-diabetic individuals. Arthritis research & therapy 14(3): R135.
-
Shippey EA, Wagler VD, Collamer AN (2018) Hydroxychloroquine: An old drug with new relevance. Cleveland Clinic journal of medicine 85(6): 459-467.
-
Yam JC, Kwok AK (2006) Ocular toxicity of hydroxychloroquine. Hong Kong Medical Journal 12(4): 294-304.
-
Tehrani R, Ostrowski RA, Hariman R, Jay WM (2008) Ocular toxicity of hydroxychloroquine. Seminars ophthalmology 23(3): 201-209.
-
Shearer RV, Dubois EL (1967) Ocular changes induced by long-term hydroxychloroquine (Plaquenil) therapy. American journal of ophthalmology 64(2): 245-252.
-
Elder M, Rahman AM, McLay J (2006) Early paracentral visual field loss in patients taking hydroxychloroquine. Archives of ophthalmology 124(12): 1729-1733.
-
Blyth C, Lane C (1998) Hydroxychloroquine retinopathy: is screening necessary?: Intensive screening is not necessary at normal doses. BMJ 316(7133): 716-717.
-
Bernstein HN (1991) Ocular safety of hydroxychloroquine. Annals of ophthalmology 23(8): 292-296.
-
Tobin DR, Krohel GB, Rynes RI (1982) Hydroxychloroquine: seven-year experience. Archives of Ophthalmology 100(1): 81-83.
-
Weiner A, Sandberg MA, Gaudio AR, Kini MM, Berson EL (1991) Hydroxychloroquine retinopathy. American journal of ophthalmology 112(5): 528-534.
-
Yam JC, Kwok AK (2006) Ocular toxicity of hydroxychloroquine. Hong Kong Medical Journal 12(4): 294-304.
-
Maturi RK, Folk JC, Nichols B, Oetting TT, Kardon RH (1999) Hydroxychloroquine retinopathy. Archives of Ophthalmology 117(9): 1262-1263.
-
Fernandez AP (2017) Updated recommendations on the use of hydroxychloroquine in dermatologic practice. Journal of the American Academy of Dermatology 76(6): 1176-182.
-
Brewer GJ, Zarafonetis CJ (1967) The haemolytic effect of various regimens of primaquine with chloroquine in American Negroes with G6PD deficiency and the lack of an effect of various antimalarial suppressive agents on erythrocyte metabolism. Bulletin of the World Health Organization 36(2): 303-308.
-
Puri PK, Lountzis NI, Tyler W, Ferringer T (2008) Hydroxychloroquine‐induced hyperpigmentation: the staining pattern. Journal of cutaneous pathology 35(12): 1134-1137.
-
Coulombe J, Boccara O (2017) Hydroxychloroquine- related skin discoloration. Canadian Medical Association Journal 189(5): E212.
-
Siddiqui AK, Huberfeld SI, Weidenheim KM, Einberg KR, Efferen LS (2007) Hydroxychloroquine-induced toxic myopathy causing respiratory failure. Chest 131(2): 588-590.
-
Granel B, Gaudy C, Serratrice J, de Roux Serratrice C, Felician O, et al. (2001) Psychological and behavioral disorders with good outcome in neurosarcoidosis. La Revue de Medecine Interne 22(2): 183-188.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions