KLF4 Protects Endothelial Cells against Senescence by Activating PDGF-BB/PDGFRA Pathway
Aims: To explore the role KLF4 plays in regulating senescence of endothelial cells and the specific mechanism of it. Main Methods: Human umbilical vein endothelial cells (HUVECs) were cultured with glucose or angiotensin (Ang) II to induce senescence. Expressions of KLF4, PDGF-BB, PDGFRA and PDGFRB in normal and senescent HUVECs were assessed by mRNA-seq and protein microarray. KLF4/PDGF-BB/PDGFRA axis then was affected using sh-RNA. Fluorescent dual luciferase reporting system and EMSA were used to determine if KLF4 directly promoted transcription of PDGF-BB/PDGFRA. Senescence of aortic wall in wild type C57BL/6 mice, STZ-induced diabetic mice fed with high-fat diet (HFD) and endothelial cells (ECs)- specific KLF4 knockout mice was assessed by SA-β-Gal staining. Key Findings: STZ-induced diabetes with high-fat diet and absence of endothelial KLF4 led to accelerated senescence of aortic vessel wall in mice. Whole mRNA-seq and protein microarray analysis revealed decreased expression of KLF4, PDGFBB, PDGFRA and PDGFRB in senescent HUVECs. Knock-in of KFL4, PDGF-BB or PDGFRA, but not PDGFRB, led to impaired senescence in glucose treated HUVECs, whereas knock-down of KLF4, PDGF-BB or PDGFRA directly accelerated senescence as revealed by greater proportion of positive SA-β-Gal staining, higher inflammatory factors level, more frequent G2/M cell cycle arrest and typical cellular nuclear senescent morphology. KLF4 directly promoted transcription activity of PDGF-BB/PDGFRA as revealed by fluorescent dual luciferase reporting system and EMSA. Incubation of HUVECs with PDGF-BB and PDGFRs vectors increased KLF4 expression. Significance: In diabetic mice models and glucose/Ang II-induced HUVECs, KLF4 protects endothelial cells against senescence by activating PDGF-BB/PDGFRA pathway.
Introduction
The vascular endothelium, composed of an endothelial cell (EC) monolayer, is the primary barrier against atherosclerosis promoting factors, such as lipids, glucose, nicotine, angiotensin II (Ang II) and altered blood flow. Damage or senescence of ECs contributes to the formation of atherosclerotic plaques. Senescent ECs cease to proliferate while continuing to release inflammatory mediators such as interleukin (IL)-1, intracellular adhesion molecule 1 (ICAM-1), tumor necrosis factor (TNF)-α and monocyte chemoattractant protein (MCP)-1, all of which contribute to atherosclerosis progression by promoting monocyte chemotaxis, foam cell formation, fibrous cap erosion or rupture, and thrombosis [1].
Platelet-derived growth factor-BB (PDGF-BB), a known potent mitogen and chemoattractant for vascular smooth muscle cells (VSMCs) and other inflammatory cells, is expressed at high level in almost all different types of cells in atherosclerotic aortic walls [2] and is a crucial regulator of neointimal enlargement [3]. Recently, researchers found that PDGF-BB and its downstream factors play pivotal roles to maintain ECs proliferation [4], induce ECs immature angiogenesis [5], keep the self-renewal and stemness function of endothelial progenitor cells [6] and improve mitochondrial energy biogenesis to repair LPS-induced endothelial injury [7]. Suppression of PDGF-BB activation induced cellular senescence and apoptosis in radiation-, oxidative stress- and inflammation-mediated injuries [8, 9, 10]. Therefore, we suspect that PDGF-BB is a potential suppressor of ECs senescence.
Krüpper-like factor 4 (KLF4), also known as gut- enriched Krüpper -like factor (GKLF) or epithelial zinc- finger protein (EZF), belongs to the SP/KLF family of transcription factors [11]. Characterized by a C-terminal three zinc-finger domain [12] and both a transactivation (TAD) and a repressor domain, KLF4 is a bifunctional transcriptional regulator in cell cycle progression, including maintenance of pluripotency of stem cells, regulation of cellular proliferation, and activation of autophagy [13]. In cardiovascular system, KLF4, as a key housekeeper gene in VSMCs, controls phenotypic switching [14], down-regulates expressions of surface markers [15], and represses growth [16], while promotes eNOS activity [17], promotes dilation of resistant arteries and increases blood flows [18]. In ApoE-/- mice, deletion of KLF4 in endothelial cells increased atherosclerosis while over-expression of KLF4 has the opposite effect [19] but the specific mechanism remains to be defined. Beyond shear-stress induced KLF4 expression and sequential cell transcriptome transcription [20], there remains much to be learned about how KLF4 regulates activities and functions of ECs. Recent evidence suggested that activation of KLF4 may induce angiogenesis of pulmonary artery [17]. Because senescent ECs lose angiogenic activity [21], we posited that KLF4 may restrict endothelial cells senescence.
Although PDGF-BB has been suggested to exert functions with the activation of KLF4 [17, 22, 23], in most cases, it does so via binding to PDGFR, including PDGFRA (PDGFR a) and PDGFRB, and downstream genes [24]. So, the relationship between KLF4 and PDGF-BB/PDGFR warrants further elucidation, and therefore the present study aims to assess whether KLF4 plays a critical role in senescence of ECs via the PDGF-BB/PDGFRs pathway.
Materials and Methods
Animals
All animal experimental procedures were performed in accordance with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH revised 2011) and approved by the Institutional Animal Care and Use Committee of Tongji University. Eight-week-old C57BL/6J male mice were purchased from the Experimental Animal Centre, Chinese Academy of Sciences (Shanghai, China). The conditional ECs-specific KLF4 knockout (KLF4EC-/-) mice were established by Saiye Corp. (Suzhou, Jiangsu, China). Briefly, we crossbred Tek-Cre: KLF4 and flox/ flox mice. Genotyping was performed at 2 weeks after birth. The Tek-Cre: KLF4 flox/flox mice had ECs-specific KLF4 deletion called KLF4EC-/- mice.
For animal models, a total of 12 eight-week-old C57BL/6J mice were randomly divided into two groups, the wild type (WT) group and the STZ-induced high-fat fed (STZ+HFD) diabetic mice group, 6 mice in each group. After 12 h of fasting, STZ+HFD group mice were injected intraperitoneally with STZ dissolved in citrate buffer (pH 4.5) at the dose of 70 mg/kg and then fed with high-fat diet for 8 weeks and the WT group mice were injected intraperitoneally with the citrate buffer vehicle and fed with normal diet for 8 weeks. The mice were anesthetized using 4% isoflurane inhalation and then sacrificed using cervical dislocation. After carefully cleaning the thrombus inside the lumen, the thoracic and abdominal aortas were prepared to execute the following operations.
Protein Obtaining From Different Aspects of The Aortic Vessel Wall
As described in previous document [7], different aspects of the aortic vessel wall were obtained by the following procedure. First, we used 0.25% trypsin to perfuse the aortic lumen for 1 min, the trypsin after perfusion was collected and centrifuged, and the precipitations were considered to be aortic endothelial cells. Then the residual vessel walls were undergoing continual digestion in 0.25% trypsin for 45 min, then the trypsin was collected and centrifuged, and the precipitations were considered to be aortic smooth muscle cells. The residual tissue was digested in 0.25% trypsin for 60 min, the trypsin was filtering and centrifugation, and the residuum was identified to be aortic adventitia cells. The cells were transferred to protein lysate.
Aortic Vessel Walls Preparation For Sa-Β-Gal Staining
The complete aortic vessel walls were carefully washed by PBS, and then were cut open along the vertical axis. The inner aspects without adipose tissue around were usually known as luminal aspects. After staining with SA-β-gal, the luminal aspects were used to take photos.
Reagents
PDGF-BB was purchased from PeproTech (Rocky Hill, U.S.). Adv-vectors (KLF4, PDGF-BB, PDGFRA and PDGFRB, VigeneBio, Shandong, China) were used to over-express target genes, and CMV-sh-RNAs (KLF4, PDGF-BB, PDGFRA and PDGFRB, VigeneBio, Shandong, China) were used to knock out target genes; all were purchased from Viogene Bio. (Jinan, Shandong, China). KLF4 luciferase reporter plasmid (pGL4-KLF4) was purchased from Yeasen Bio. (Shanghai, China). Fluorescent dual luciferase reporting system and murine leukemia virus reverse transcriptase were obtained from Promega Co. (Mannheim, Germany). The cell cycle kit was purchased from BD (Franklin Lakes, USA). The SA-β-gal kit was obtained from Beyotime (Haimen, China). Amino Allyl messageAmp II kit was purchased from Life Technologies (Massachusetts, USA). The anti-p21, anti- PDGF-BB, anti-PDGFRA, anti-PDGFRB, anti-KLF4, anti-vWF, anti-a-SMA, anti-Vimentin and anti-β-actin, and secondary antibodies were obtained from Cell Signaling (Denver, USA). Other reagents were purchased from Sigma (St. Louis, USA). The sequences of CMV-sh-RNAs are as following:
CMV-KLF4-sh-RNA: 3’-GGCTGGATATTATCAACTCACTG-5’ CMV-PDGFRA-sh-RNA: 3’- AAACTGTAGAGCTTCGATGTAAA-5’ CMV-PDGFRB-sh-RNA: 3’- AATTAGAATGTTACTCAATCAGC-5’ CMV-PDGF-BB-sh-RNA: 3’- CCAACGCCAACTTCCTGGTGTGG-5’
Cell Culture
The human umbilical vein endothelial cells (HUVECs) line and endothelial cell media (ECM) were obtained from ScienceCell (Carlsbad, California, USA). The HUVECs were grown in ECM (glucose: 5.5mmol/L). After 5 times generation, the cells were starved by deprivation of FBS for 12h. Then the cells were then exposed to Ang II (1umol/L) or Glucose (33mmol/L) for 72 h to induce HUVECs senescence.
Adenovirus and cmv-sh-rna Transfection
Adv-vectors (KLF4, PDGF-BB, PDGFRA and PDGFRB) were used to knock in target genes, and CMV-sh-RNAs (KLF4, PDGF-BB, PDGFRA and PDGFRB) were used to knock down target genes. Adv-empty vector, CMV-control-sh-RNA, Adv- KLF4, PDGF-BB, PDGFRA and PDGFRB –vectors and CMV- KLF4, PDGF-BB, PDGFRA and PDGFRB -sh-RNA were directly added to cultured cells (50MOI) and incubated for 48 h. For co-transfection, Adv-KLF4-vector/CMV-KLF4-sh-RNA was added at the same time with CMV-PDGF-BB/PDGFRA-sh- RNA or Adv-PDGF-BB/PDGFRA-vector, while HUVECs in the control group were also added with CMV-control-sh-RNA and Adv-empty vector.
Real-Time Reverse Transcription-pcr (Rt-Pcr)
Total RNA was converted to cDNA using the murine leukemia virus reverse transcriptase system. RT-PCR was performed using an ABI 7500 system and TB GREEN Premix EX TaqTM II (Takara, Shiga, Japan) following manufacturer’s instructions.
Western Blot
Proteins were obtained from cultured HUVECs or mice aortic wall. Equal amounts of proteins were subjected to SDS-PAGE and then blotted on the PVDF membranes, which then were incubated with primary antibodies at 4℃ overnight. After incubation with secondary antibodies at room temperature for 1 h, the membranes were visualized using enhanced ECL agent.
Cell Cycle Analysis
After fixation with 70% ethanol at 4℃, the cells were incubated with 50 μg/ml propidium iodide (PI), 100 μg/mL DNAase-free RNAase and 2‰ Triton-X-100 for 45 min. Flow cytometry was used for cell cycle analysis.
Sa-Β-Gal Analysis
SA-β-gal analysis was performed according to the SA- β-gal kit manufacturer’s instructions. Briefly, after fixation for 15min, fresh aortic walls and cells were incubated with 5-bromo-4-chloro-3-inolyl-b-D-galactoside (β-gal) substrate for 24 h at 37 °C and then pictured under an optical microscope. Qualified pictures were chosen to be presented.
Sahf Detection
After fixation with 90% ice-cold methanol for 2 min, cells were stained with 0.1ug/mL anti-fade DAPI for 2 min, and then analyzed using a laser confocal microscope.
msa
The probes (1000 fmol) used were: PDGF-BB, 5’-gcttgttaccacacccagctccag-3’, 3’-cgaacaatggtgtgggtcgaggtc-5’; PDGFRA, 5’-caggcgcaaccaggcccaggtggc-3’, 3’-gtccgcgttggtccgggtccaccg-5’; PDGFRB, 5’-gttgaggctgggtgcggtggctcaag-3’, 3-caactccgacccacgccaccgagttc-5’. The samples were resolved on a TBE-PAGE and then visualized using X-ray.
Fluorescent Dual Luciferase Reporting Analysis
For luciferase analysis, the pGL4-KLF4 vector and PDGF- BB, PDGFRA and PDGFRB vectors were co-transfected into 105 HEK 293T cells and then incubated for 48 h. The samples were collected and subjected to 3 freeze-thaw cycles. The results were obtained using a Lumat LB9507 luminometer (EG&G Berthold). Transfection efficiency was normalized to Renilla luciferase.
Mrna-Seq Array
The chip used in this analysis is synthesized by Riobio- tech. Inc. (Guangzhou, China). RNA was extracted from pre-treated HUVECs using Trizol, and then the extracted RNA was inspected by UV spectrophotometer, agarose gel electrophoresis and Agilent 2200 Bioanalyzer (A260/A280 ≥ 1.5, A260/A230 ≥ 1, RIN value ≥ 7; If contamination was found by electrophoresis, DNAase was used to digest). The purified RNA was stored at -80℃. 0.5-2ug of the purified RNA was used to generate antisense RNA by Amino Allyl messageAmp II kit, then the antisense RNA was labeled with Cy3/Cy5, and underwent measurement of A260, A550 by UV spectrophotometer, after that, the labeling rate DOL was calculated according to the formula listed in the experimental report (the best range of DOL is 1.0-3.6). For the pre- hybridization, the chip was incubated in sterile deionized water at 65°C for 10 minutes, and then changed into pre- hybridization solution at 37°C and incubated for 1 hour. The fluorescently labeled antisense RNA was quantified; 4-8ug was taken to prepare the hybridization solution, denatured at 95°C for 3min, incubated on ice for 1min. The hybridization solution then was added to replace the pre-hybridization solution for the chip and hybridized at 37℃for 24h. The chip was then washed by 6X SSPET, 3X SSPET, 0.5X SSPET, PBST in turn to remove non-specific hybridization background. Cover slip was added to the hybridization area and the chip was then underwent scan.
Inflammatory Factors Spectrum Array
Inflammatory factors spectrum array from total lysis proteins of HUVECs was performed by Riobio-tech. Inc. (Guangzhou, China)
Statistical Analysis
Statistical analysis was performed using SPSS22.0 software. All data are presented as mean ± SEM, and were compared using one-way ANOVA followed by Tukey’s HSD test. A value of p< 0.05 was considered to be statistically significant.
Results
Endothelial KLF4 is A Major Anti-Senescence Factor of Aortic Wall Senescence
The development of aortic vessel wall senescence is a complex process of what the initiation and progression involve key molecular and structural elements. We generated a STZ- induced diabetic/high-fat diet (HFD)-induced senescence mice model (STZ+HFD mice) using C57BL/6J mice that first received STZ (70 mg/kg) i.p. injection and then were fed with HFD for 8 weeks, the blood glucose level of mice in these two different groups were tested to show that the STZ+HFD mice models were successfully built (Figure 1A). The level of SA-β- gal positive staining burden was much higher in the luminal aspects of the aortic vessel walls (without adipose tissue around aspects) in STZ+HFD mice than in WT mice (Figure 1B). To explore in which cell type KLF4 expression has the most obvious change in the senescent aortic vessel walls, we separated KLF4 from different substructures of the aortic vessel walls, tested the sample purity by flow cytometry with corresponding markers and found that the KLF4 expression in endothelial cells (marked by vWF) decreased substantially compared with that in aortic SMCs (marked by a-SMA) and adventitia fibroblasts (marked by Vimentin) (Figure 1C- 1D), suggesting that endothelial KLF4 might be involved in aortic wall senescence. To further verify this possibility, we abrogated KLF4 expression in aortic endothelium by Cre-flox transgenetic mice and got KLF4EC-/- mice (Figure 1E). Even without HFD and STZ treatment, SA-β-gal positive stained burden was significantly higher in KLF4EC-/- mice compared with wild-type (WT) mice (Figure 1F). Also, we detected the expression of KLF4 in glucose- or Ang II-induced senescent HUVECs. Western blot analysis showed that KLF4 protein expression also reduced in glucose- or Ang II-induced senescent HUVECs (Figure 1G).
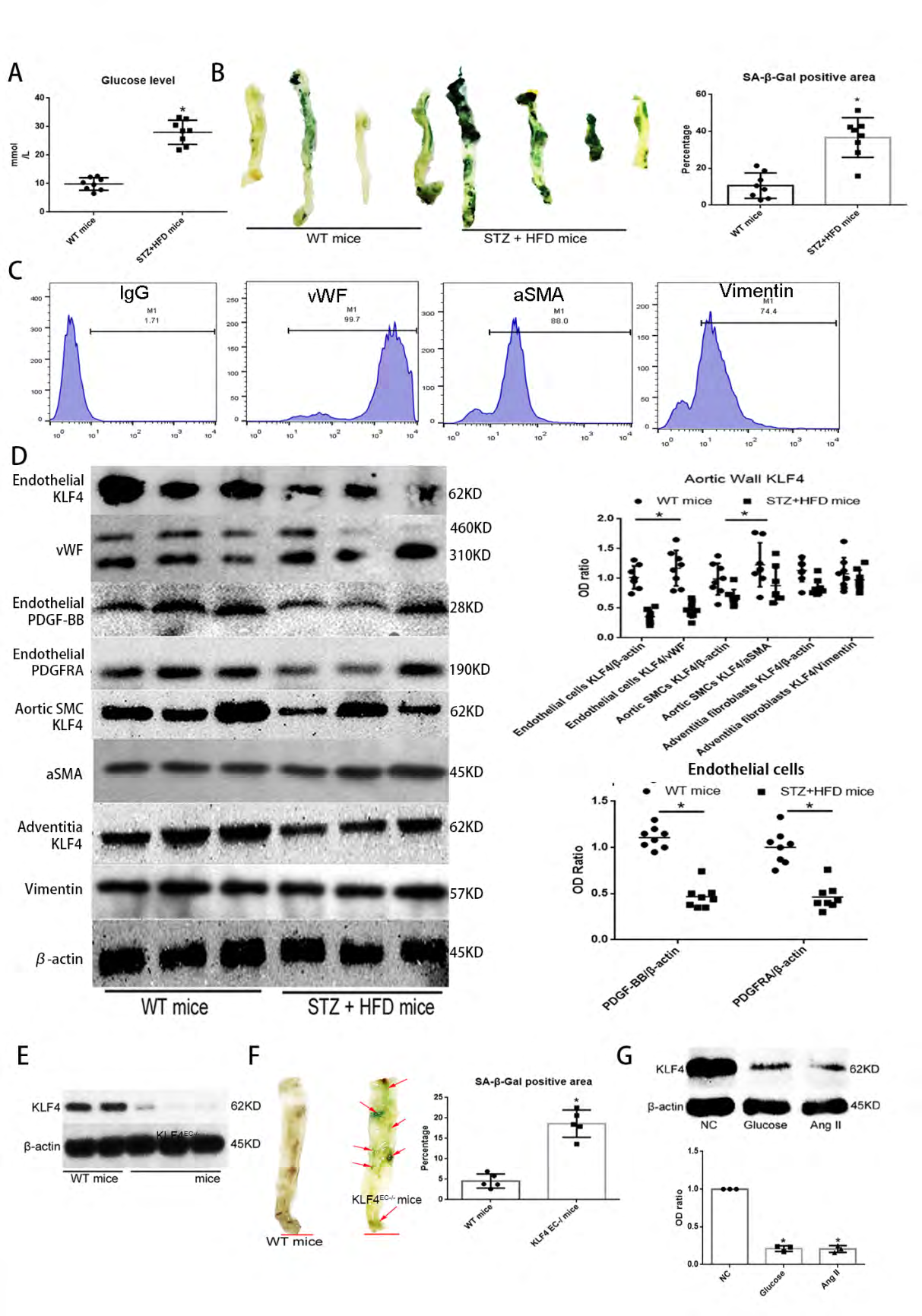
Figure 1: KLF4 inhibits aortic vessel wall senescence: (A) Blood glucose level between WT mice group and STZ induced diabetic mice with HFD (STZ+HFD) mice group. WT, wild type; STZ, streptozocin; HFD, high fat diet. (B) SA-β-gal detection and quantification of aortic vessel walls. STZ+HFD mice were used as pro-senescence models. Green, SA-β-gal positive area; N=8; *p<0.05 versus WT mice. (C) Flow cytometry for purity of samples from different aortic vessel wall structural elements. (D) Western blot analysis and quantification for KLF4 in different aortic wall structural elements, PDGF-BB and PDGFRA in endothelium of aortic vessel walls between WT mice group and STZ+HFD mice group. N=8; *p<0.05 versus WT mice. (E) Western blot analysis for KLF4 in endothelium of aortic vessel walls between WT mice group and KLF4EC-/- mice group. N=5. (F) SA-β-gal detection and quantification of aortic vessel walls between WT mice group and KLF4EC-/- mice group. Green, SA-β- gal positive; N=5; *p<0.05 versus WT mice; Scale bar, 2mm. (G) Western blot analysis and quantification for KLF4 among NC ECs, high glucose- (33mM) or Ang II- (1uM) treated ECs. N=3; *p<0.05 versus NC ECs. ECs, HUVECs; NC, non-specific control; Ang II, angiotensin II; Glu, glucose.
KLF4 Restricts Senescence of Glucose-Induced HUVECs
To further confirm the essential role of KLF4 in senescence of endothelial cells, we used Adv-KLF4-vector and CMV-KLF4-sh-RNA to build KLF4 knock-in HUVECs (KLF4+/ECs) and KLF4 knock-down HUVECs (KLF4-/ECs) and non-specific control HUVECs (NC ECs) were transfected with empty vectors as a control group, the knock-in and knock-out efficiency of KLF in HUVECs were checked by qPCR (Figure 2A). We validated senescence by SA-β-gal staining, cell cycle arrest, inflammatory factors secretion and senescence related heterochromatin foci (SAHF). SA-β-gal staining (Figure 2B) positive cells percentage and SAHF (Fig. 2C) level, G1 and G2/M phase arrest (Figure 2D), p21 expression (Figure 2E) and inflammatory factors levels (Figure 2F) were all increased in KLF4-/ECs compared with NC ECs. In contrast, KLF4+/ECs showed lower levels of glucose-induced SA-β-gal staining cell percentage (Figure 2A), G0/G1 and G2/M phase arrest (Figure 2D), p21 expression (Figure 2E) and secretion of inflammatory factors (Figure 2F). The latter findings suggested that KLF4 might be an inhibitor of ECs senescence.
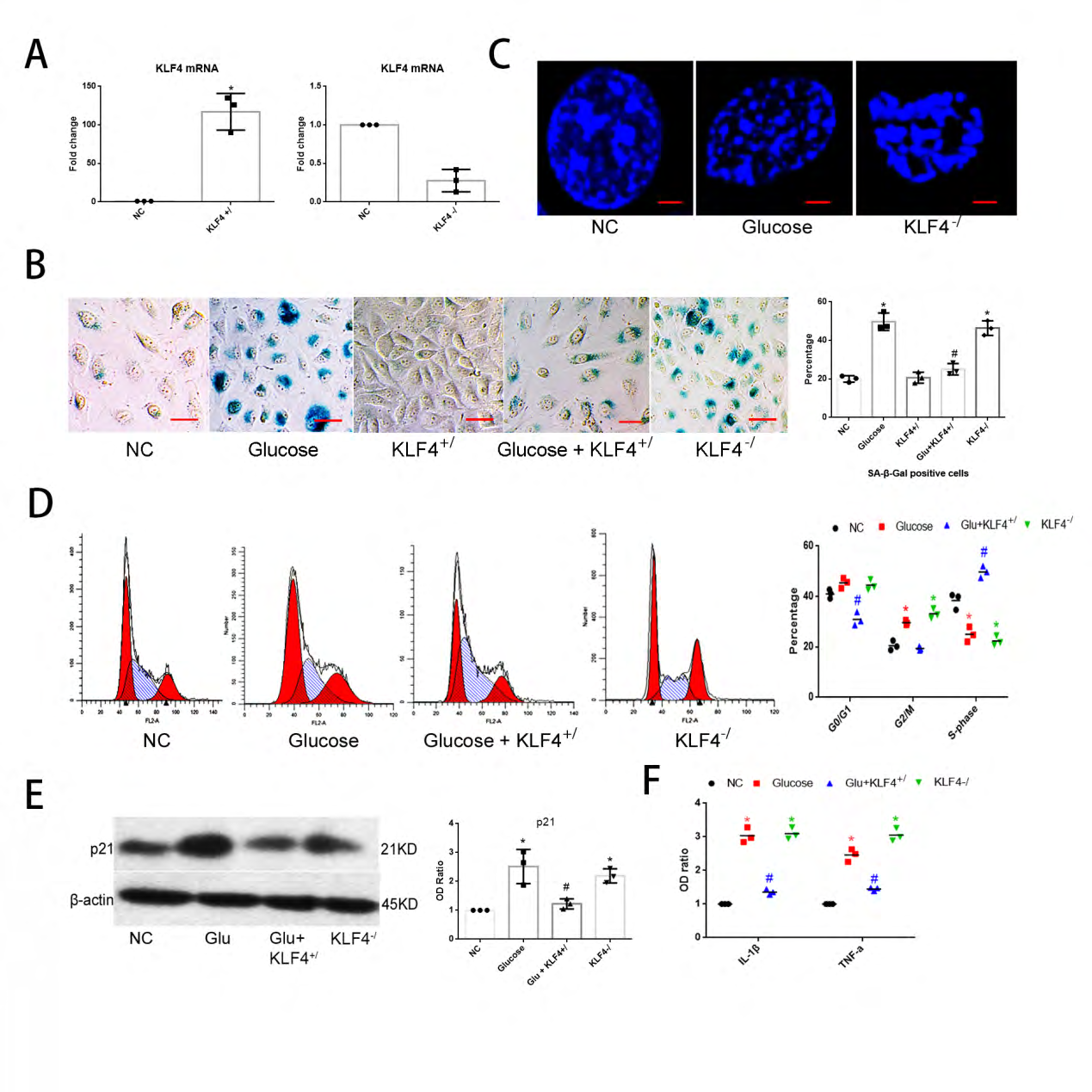
Figure 2: KLF4 inhibits ECs senescence: (A) qPCR for KLF4 mRNA expression to test the efficiency of KLF4+/ or KLF4-/ vectors. Non-specific control HUVECs (NC) transfected with empty vectors were used as a control group. (B) SA-β-gal detection and quantification of glucose-induced KLF4+/ or KLF4-/ HUVECs. Non-specific control HUVECs (NC) transfected with empty vectors were used as a control group. Green, SA-β-gal positive cells; N=3; *p<0.05 versus NC; #p<0.05 versus glucose-induced HUVECs. Scale bar, 100um. (C) Typical SAHF structure under confocal microscope in glucose- or KLF4-/ induced HUVECs. Scale bar, 10um. (D) Flow cytometry for cell cycle phase. N=3; *p<0.05 versus NC; #p<0.05 versus glucose-induced HUVECs. (E) Western blot analysis and quantification for p21. N=3, *p<0.05 versus NC ECs; #p<0.05 versus glucose-induced HUVECs. (F) qPCR for IL-1β and TNF-α mRNA expression. N=3; *p<0.05 versus NC; #p<0.05 versus glucose-induced HUVECs. NC, non-specific control; Glu, glucose; KLF4+/, KLF4 knock-in; KLF4-/, KLF4 knock-down.
MRNA-Seq Analysis Reveals That KLF4 Regulates Senescence-Related Pathways
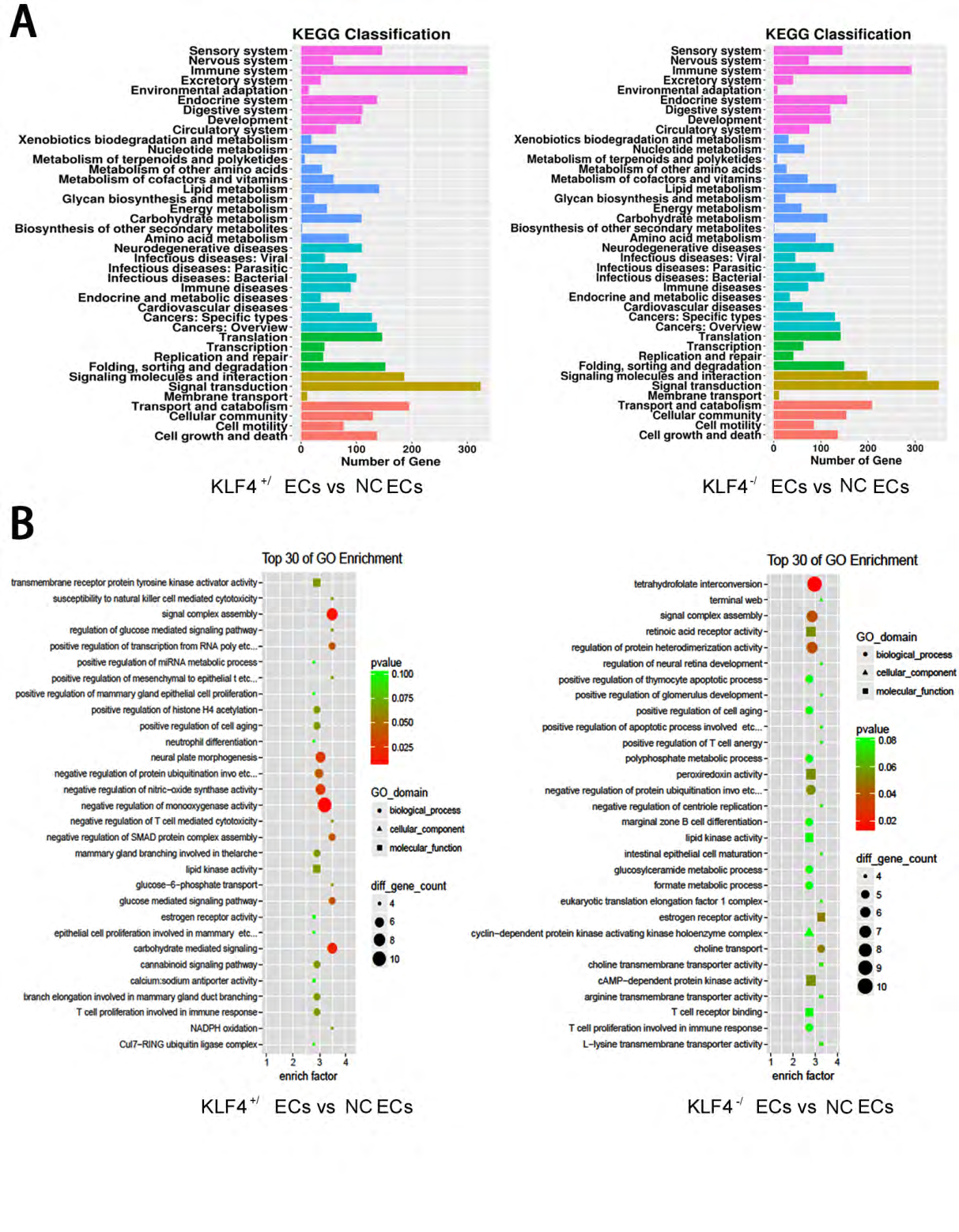
To verify the possible association between KLF4 and endothelial cell senescence, we first exerted mRNA-seq analysis among NC ECs, KLF4+/ECs and KLF4-/ECs. We chose genes with an increase of more than 2 folds and with p-value <0.05 or a decrease of more than 50% and with p-value <0.05 for KEGG (Figure 3A) analysis and then we picked up TOP30 genes with up-regulation or down-regulation changes for GO analysis (Figure 3B). The GO and KEGG analysis revealed that in KLF4+/ECs and KLF4-/ECs , the expression of cell cycle processes, NO synthase, CDK complex, immune system and cell aging process, which are all related with cellular senescence, were significantly different from those in NC ECs.
PDGF-BB/PDGFRA Pathway Inhibits Senescence in HUVECs
Because senescent cells secrete cytokines and in turn these cytokines regulate cellular senescence [25], we examined cytokine expressions in NC ECs, KLF4+/ECs and KLF4-/ECs using protein chip technology. We found that the expressions of many cytokines, including PDGF-BB, MCP-1, IL-1β and RANTES, were significantly changed (Figure 4A, Table 1). Then we analyzed mRNA-seq results again. We found that PDGF-BB and its receptor PDGFR were obviously different in KLF4-/ECs and KLF4+/ECs compared with NC ECs by KEGG pathway analysis (Figure 4B). Next, we validated the protein chip and mRNA-seq results by Western blot. The expression of PDGF-BB and its receptor PDGFRA, but not PDGFRB, were significantly higher in KLF4+/ECs compared with in NC ECs (Figure 4C). All the results suggested that KLF4 may regulate ECs senescence through the PDGF-BB/ PDGFRA pathway.
To determine the role of PDGF-BB/PDGFRA in KLF4- regulated senescence of HUVECs, we first studied the expression level of PDGF-BB/PDGFR pathway in glucose- induced senescent HUVECs. Expression of PDGF-BB and PDGFRA were lower in glucose-induced senescent HUVECs, while PDGFRB expression was similar to that in NC ECs (Figure 4D). Also, we found the protein level of PDGF-BB and PDGFRA decreased in the endothelium of aortic vessel walls in STZ+HFD mice compared to WT mice (Figure 1D). Then we examined the senescent phenotype of PDGF-BB knock-down HUVECs (PDGF-BB-/ECs) and PDGFRA knock- down HUVECs (PDGFRA-/ECs). Results showed that SA-β-gal
staining (Figure 4E) and p21 expression (Figure 4F), SAHF positive cells percentage (Figure 4G), and G0/G1 and G2/M phase arrest (Figure 4H) were all increased in PDGF-BB-/ECs and PDGFRA-/ECs, compared with NC ECs.
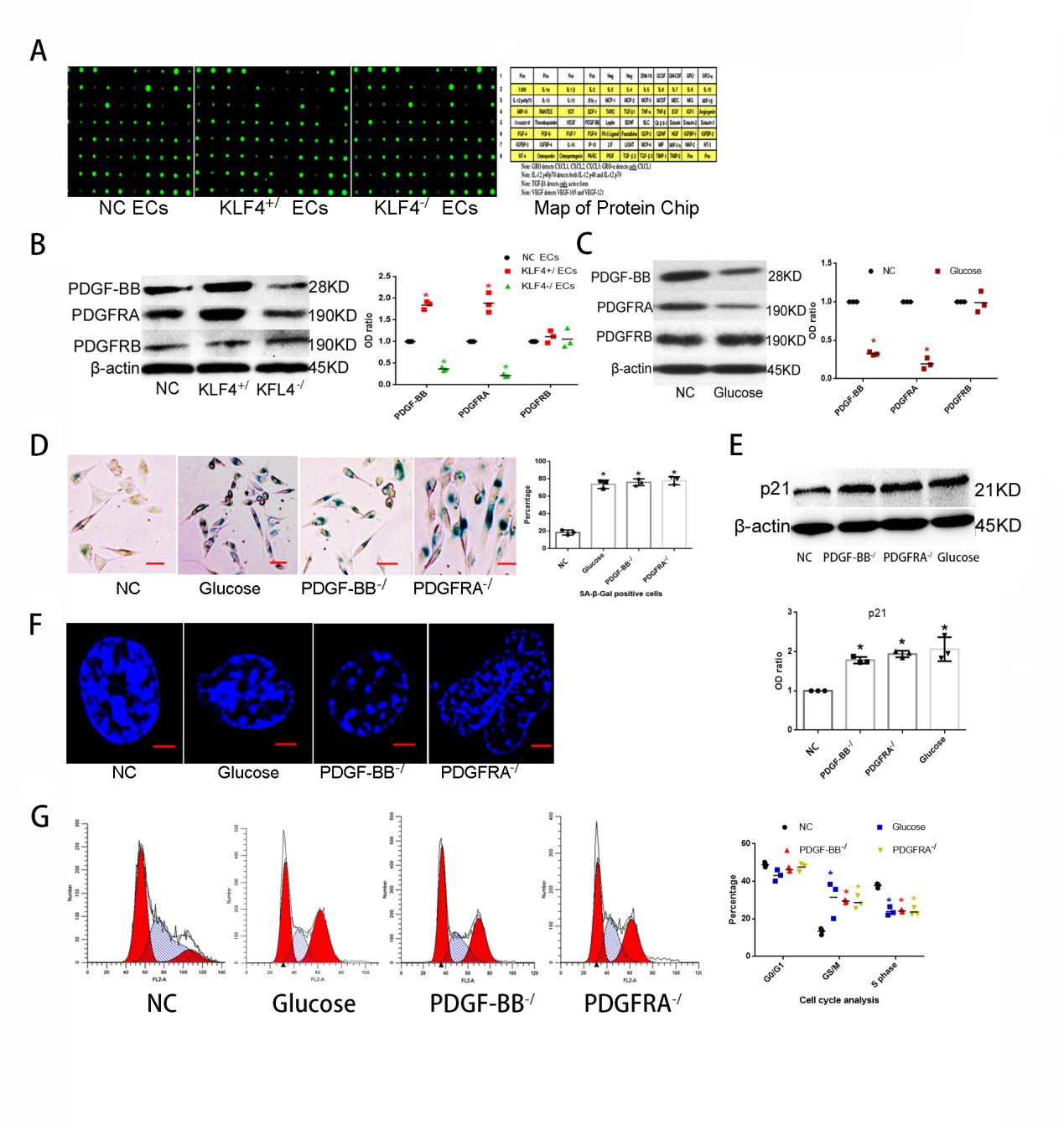
Figure 4: KLF4/PDGF-BB/PDGFRA participates in ECs senescence: (A) Cytokines protein chip analysis in NC ECs, KLF4+/ECs and KLF4-/ECs. N=3. (B) KEGG pathway analysis in KLF4+/ECs and KLF4-/ECs. N=3. (C) Western blot analysis and quantification for PDGF-BB/PDGFRs protein expression in NC ECs, KLF4+/ECs and KLF4-/ECs. N=3; *p<0.05 versus NC ECs. (D) Western blot analysis and quantification for PDGF-BB/PDGFRs protein expression in NC ECs and glucose-induced ECs. N=3; *p<0.05 versus NC ECs. (E) SA-β-gal detection and quantification in glucose-induced ECs, PDGF-BB-/ ECs and PDGFRA-/ ECs. Green, SA- β-gal positive cells; N=3; *p<0.05 versus NC ECs. Scale bar, 100um. (F) Western blot analysis and quantification for p21. N=3, *p<0.05 versus NC ECs. (G) Typical SAHF structure under confocal microscope. Scale bar, 10um. (H) Flow cytometry for cell cycle phase. N=3. ECs, HUVECs; NC, non-specific control; KLF4+/, KLF4 knock-in; KLF4-/, KLF4 knock-down; PDGF-BB-/, PDGF- BB knock-down; PDGFRA-/, PDGFRA knock-down.
| KLF4+/ECs vs NC ECs | KLF4-/ECs vs NC ECs | |
|---|---|---|
| Inflammatory factor | OD ratio | OD ratio |
| GRO-α | 1.724* | 0.499* |
| IL-1α | 0.968 | 0.623* |
| IL-1β | 0.548* | 1.625* |
| IL-6 | 0.276* | 0.716 |
| MCP-1 | 0.23* | 0.938 |
| RANTES | 0.956 | 1.684* |
| PDGF-BB | 1.829* | 0.246* |
Table 1: Quantification of cytokines protein chip analysis
KLF4 Protects HUVECs Against Senescence Through Activating PDGF-BB/ PDGFRA Pathway
To further clarify the role of PDGF-BB/PDGFRA pathway in KLF4-restricted endothelial cells senescence, we built PDGF-BB knock-in HUVECs (PDGF-BB+/ECs) and PDGFRA knock-in HUVECs (PDGFRA+/ECs) by transfection. We also knocked down PDGF-BB or PDGFRA expression in KLF4+/ECs and knocked in PDGF-BB or PDGFRA expression in KLF4-/ECs separately by co-transfection. Transfection efficiency was tested by qPCR (Figure 5A). Neither PDGF-BB nor PDGFRA deficiency could sustain the young phenotype of glucose- treated HUVECs caused by KLF4 enrichment, and manifested with higher p21 expression (Figure 5B) and greater number of SA-β-gal staining positive cells (Figure 5C). PDGF-BB or PDGFRA enrichment efficiently suppressed EC senescence caused by glucose or KLF4 absence, showing less number of SA-β-gal staining positive cells (Figure 5D,5E) and lower expression level of p21 expression (Figure 5F,5G). The latter results indicated that KLF4 inhibits EC senescence at least partly via the PDGF-BB/PDGFRA pathway.
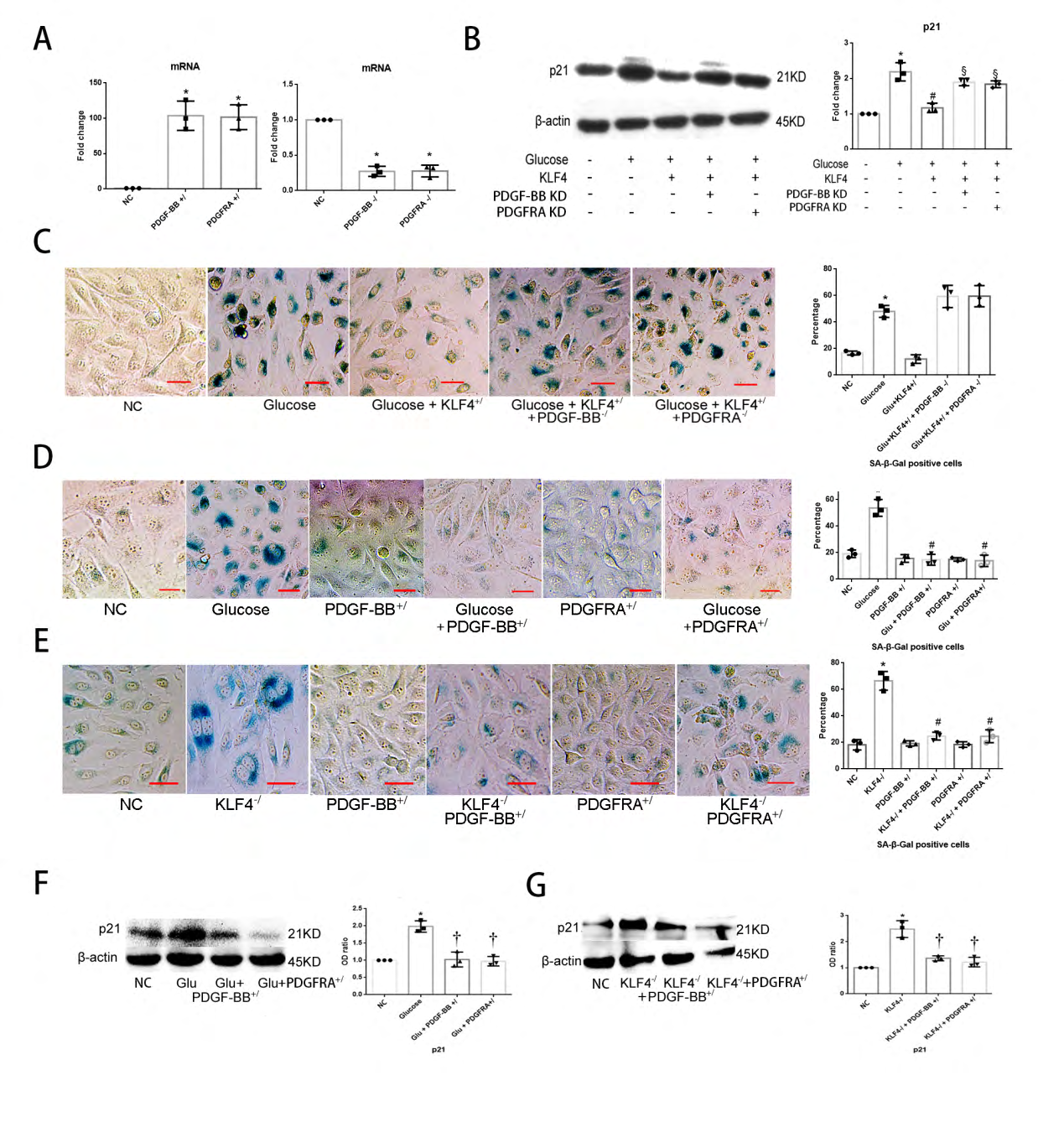
Figure 5: PDGF-BB/PDGFRA regulates ECs senescence: (A) qPCR for PDGF-BB and PDGFRA mRNA to test the efficiency of knock-in and knockdown vectors. Non-specific control HUVECs (NC) transfected with empty vectors were used as a control group. *p<0.05 versus NC. (B) Deficiency of PDGF-BB or PDGFRA increases p21 expression in KLF4+/ ECs. (C) Deficiency of PDGF-BB or PDGFRA increases SA-β-gal positive cells number in KLF4+/ ECs. Green, SA-β-gal positive cells. N=3; *p<0.05 versus NC; Scale bar, 100um. (D) Enrichment of PDGF-BB/PDGFRA decreases SA-β-gal positive cells percentage in glucose-incubated ECs. Green, SA-β-gal positive cells. N=3; *p<0.05 versus NC ECs; #p<0.05 versus glucose-incubated ECs. Scale bar, 100um. (E) Enrichment of PDGF-BB/PDGFRA decreases SA-β-gal positive cells percentage in KLF4-/ ECs. Green, SA-β-gal positive cells. N=3; *p<0.05 versus NC ECs; #p<0.05 versus KLF4-/ ECs. Scale bar, 100um. (F) Enrichment of PDGF-BB/PDGFRA decreases p21 expression in glucose-incubated ECs. N=3; *p<0.05 versus NC ECs; †p<0.05 versus glucose-incubated ECs. (G) Enrichment of PDGF-BB/PDGFRA decreases p21 expression in KLF4-/ ECs. N=3; *p<0.05 versus NC ECs; †p<0.05 versus KLF4-/ ECs. ECs, endothelial cells; Glu, glucose; NC, non-specific control; KLF4+/, KLF4 knock-in; KLF4-/, KLF4 knock-down; PDGF-BB-/, PDGF-BB knock-down; PDGFRA-/, PDGFRA knock-down; PDGF-BB+/, PDGF-BB enrichment; PDGFRA+/, PDGFRA enrichment.
KLF4 Initiates Transcription of PDGF-BB and PDGFRA
Because KLF4 has the 5’-T(A)GGGT(C)GG(T)G(A)G(T) C(T)-3’ transcriptional domain (Figure 6A), we posited that KLF4 may directly initiate PDGF-BB/PDGFRA transcription in ECs. To explore this hypothesis, we transfected pGL4- KLF4 and PDGF-BB, PDGFRA, PDGFRB vectors into HEK293T cells. After pGL4-KLF4 transfection, PDGF-BB, PDGFRA and PDGFRB effectively excited luciferase activity, however, the efficiency of transfection was much higher in PDGF-BB or PDGFRA compared with PDGFRB (Figure 6B). To better understand this connection, we exerted EMSA in HEK293T cells after KLF4 knock-in vector transfections. Strikingly, KLF4 directly bound to the promoter of PDGF-BB, PDGFRA and PDGFRB, whereas the efficiency was much more obvious in PDGF-BB or PDGFRA probes compared with PDGFRB probes (Figures 6C-6D). The latter results indicated that KLF4 directly initiated transcription of PDGF-BB and PDGFRA.
PDGF-BB/PDGFRA Induces KLF4 Expression
PDGF-BB activates KLF4 [8, 15, 16], but in our examination, KLF4 also directly promoted the transcription of PDGF-BB, suggesting a feedback loop between KLF4 and PDGF-BB. Addition of PDGF-BB peptide (30ng/mL) or PDGF- BB, PDGFRA knock-in vectors, but not PDGFRB knock-in vector, in HUVECs (Fig. 6F) resulted in increased expression level of KLF4, indicating that PDGF-BB/PDGFRA also may affect KLF4 activity.
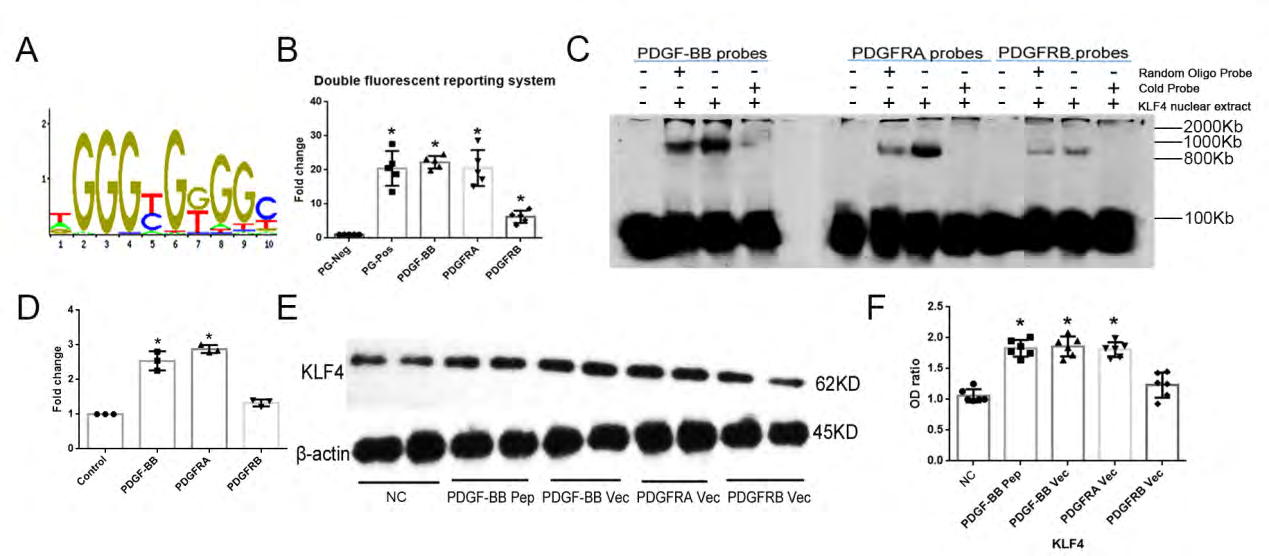
Figure 6: Activation of the KLF4/PDGF-BB/PDGFRA pathway: (A) The transcriptional domain of KLF4. (B) pGL4-KLF4 transfection excites luciferase activity in PDGF-BB and PDGFRA conditions. N=5; *p<0.05 versus PG negative control HEK293T cells. (C) EMSA shows KLF4 directly binds to PDGF-BB or PDGFRA transcriptional domain. N=3. (D) Quantifitation of EMSA blots. *p<0.05 versus control(containing only the specific-binding labeled PDGF-BB/PDGFRA/ PDGFRB probe). (E)PDGF-BB peptide or PDGFRs induces KLF4 expression. Vec, knock-in vector. N=3; *p<0.05 versus NC ECs. NC, non-specific control.
Discussion
Senescent endothelial cells lead to endothelial inflammation, high permeability, and coagulation, and thus favoring the development of atherosclerotic lesions. This study demonstrated a PDGF-BB/KLF4/PDGF-BB/PDGFRA feedback loop in senescent endothelial cells. The first major finding in this study was reduced KLF4 expression could induce accelerated senescence as evidenced by β-galactosidase activation, inflammatory factors secretion, cell cycle arrest and SAHF formation. Conversely, KLF4 enrichment strongly inhibited EC senescence. Previous analyses of various KLFs family members on maintaining endothelial homeostasis yielded poorly characterized results in mice or cultured cells [26]. ECs express KLFs 2, 4, 5, 6 and 15 [20, 27, 28, 29]. KLF2 in ECs promotes transcription of genes like endothelial nitric oxide synthase (eNOS), thrombomodulin (THBD), while inhibits multiple proinflammatory, prothrombotic, and vasoconstrictive factors, such as MCP-1, VCAM-1, E-selectin, PAI1 and ET1; in contrast, KLF2 deficiency accelerates atherosclerosis in ApoE-/- mice [30, 31]. KLF4 is considered to have the same vascular endothelium homeostasis-associated molecular patterns in regulating inflammation, oxidant stress and thrombosis, however, proof is relatively scarce. KLF4- target genes partially overlap with those of KLF2, such as eNOS, THBD, PAI1, VCAM-1 [19] and TNF-α [32]. Abrogation of KLF4 accelerated atherosclerosis [19] and acute kidney injury [33] by modulating the infiltration of lymphocytes and neutrophils, and the expression of inflammatory factors and cell adhesion molecules. However, the results either lacked sensitivity because there was KLF4 deficiency throughout the entire arterial wall [19], where endothelial cells are quantitatively negligible, or lack of assessment of the inflammatory factors and cell adhesion molecules secreted by senescent cells. Our results fill this gap in knowledge, with KLF4 expression being most obviously decreased in aortas from senescent mice, and KLF4 deficiency in aortic endothelium promoting vascular senescence. The differential secretion of inflammatory factors and cytokines between KLF4 deficient and non-specific control settings is reflected by inflammatory factors protein Chip and mRNA sequencing in cultured endothelial cells.
The second major finding in this study was that PDGF- BB deficiency enhances ECs senescence development induced by KLF4 deficiency. In previous reports, endothelial PDGF-BB was associated with ECs proliferation, migration, adhesion, angiogenesis and endothelial progenitor cells (EPCs) differentiation [33]. PDGF-BB binding to PDGFRB was lower in senescent VSMCs and fibroblasts [34, 35], and PDGF-DD bound to PDGFRB to reduce EPCs senescence [33], suggesting that PDGFRB may play a more important role in cellular senescence. Interestingly, we found that PDGFRA but not PDGFRB participate in EC senescence. PDGFRs have three isoforms, αα (PDGFRA), ββ (PDGFRB) and aβ dimer [9], which bind to different PDGF isoforms. The PDGFRA binds to all five isoforms of PDGF with high affinity, PDGFRaβ binds to other four isoforms except for PDGF-BB, whereas PDGFRB only binds to PDGF-BB or PDGF-DD [36]. Differential PDGFs binding to PDGFRs may underlie different functions through different signal transduction pathways. In the present study, PDGF-BB mainly bound to PDGFRA but not PDGFRB to inhibit EC senescence, possibly because: 1) PDGFRB expression was relatively rare compared to that of PDGFRA in the cell type studied; and 2) PDGF-BB and PDGF- DD had different excitation efficiencies of PDGFRs in cells senescent signal transduction, which may rely on the cell type and stress condition.
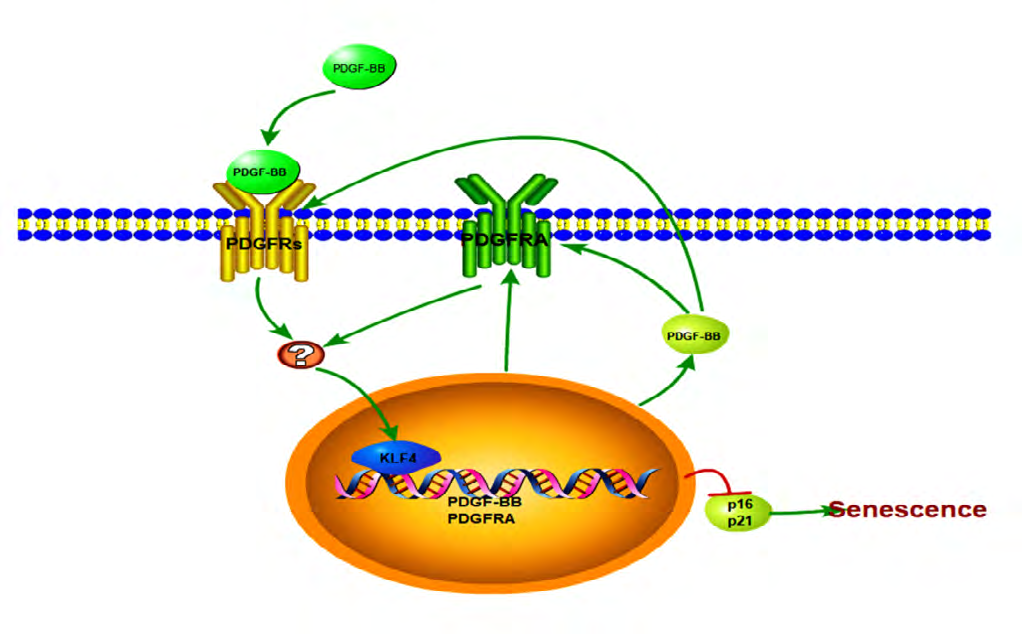
A third major finding in this study was that KLF4 directly promoted PDGF-BB/PDGFRA transcription, with a KLF4/PDGFBB/PDGFRA pathway maintaining the EC young phenotype. PDGFRs are trans membrane tyrosine kinase receptors, which could act as scaffold molecules to trigger signaling pathways, such as Ras/Raf/MEK, PI3K/Akt/mTOR, Grb2/Sos1/ Erk1/2, Rho/ROCK and NF-Κb [37], and then activate transcription factors, including KLF4, to bind to specific genes [9]. Therefore, KLF4 may serve as a key link point in PDGF-BB/PDGFRs pathway [3, 22, 24, 37]. However, whether KLF4 regulates PDGFs/PDGFRs remains unclear. Although Haskins R.M. had documented that regulation of the PDGF signaling pathway involving Myc, Pik3r1, Erf, Srgap2 and Raf1, differed between SMC KLF4 KO and wild-type animals using ChIP-seq analyses [18]. In the present study, we firstly confirmed that there was multi-motif domain of KLF4 [38] in PDGF-BB and PDGFRA promoter seed region. Then, use of the fluorescent dual luciferase reporting system and EMSA evinced that KLF4 directly triggers PDGF-BB and PDGFRA transcription. Moreover, PDGF-BB peptide and PDGFRs vectors induced KLF4 expression. Taken together, these results point to the existence of a KLF4/PDGF-BB/ PDGFRA feedback loop (Figure 7).
Conclusion
Enrichment atheroprone senescent ECs is characterized by low and inefficient KLF4, PDGF-BB and PDGFRA competence. This study highlights the involvement of PDGF- BB/PDGFRA in KLF4-deficiency induced ECs senescence, and establishes a feedback loop of PDGFBB/ KLF4/ PDGF- BB/PDGFRA signaling transduction. Further study is warranted to understand the PDGF-BB/ KLF4/PDGF-BB/ PDGFRA feedback loop, and the key points between PDGF- BB and KLF4 activity.
- Conflict of Interest statement The authors declare that there are no conflicts of interest.
- Author Contributions Jing Tong: Conceptualization, Methodology, Investigation, Writing—original draft preparation. Bing Ji: Conceptualization, Methodology, Investigation, Writing—original draft preparation. Yan-Hua Gao: Methodology, Investigation, Validation, Formal analysis. Hao Lin: Validation, Investigation. Fan Ping: Investigation, Formal analysis. Jia-Ni Tang: Formal analysis, Data Curation. Fei Chen: Writing - Review & Editing, Visualization. Xue-Bo Liu: Supervision, Project administration, Funding acquisition.
- Funding This work was supported by the China National Natural Science Foundation, No’s: 81500381 and 81670403; Shanghai science and Technology Commission medical guidance project, No: 18411950300 and 2019LJ10.
- Acknowledgments We thank Prof. Fei Zheng for help with the western blot experiments.
References
-
Ma S, Fan L, Cao F (2019) Combating cellular senescence by sirtuins: Implications for atherosclerosis. Biochim Biophys Acta Mol Basis Dis 1865(7): 1822-1830.
-
Zhao Y, Lv M, Lin H, Hong Y, Yang F, et al. (2012) ROCK1 induces ERK nuclear translocation in PDGF-BB- stimulated migration of rat vascular smooth muscle cells. IUBMB Life 64(2): 194-202.
-
Myllärniemi M, Calderon L, Lemström K, Buchdunger E, Häyry P, et al. (1997) Inhibition of platelet-derived growth factor receptor tyrosine kinase inhibits vascular smooth muscle cell migration and proliferation. FASEB J 11(13): 1119-1126.
-
Nazari M, Ni NC, Lüdke A, Li SH, Guo J, et al. (2016) Mast cells promote proliferation and migration and inhibit differentiation of mesenchymal stem cells through PDGF. J Mol Cell Cardiol 94: 32-42.
-
Díaz Flores L, Gutiérrez R, Gayoso S, García MP, González Gómez M, et al. (2020) Intussusceptive angiogenesis and its counterpart intussusceptive lymphangiogenesis. Histol Histopathol 35(10): 1083-1103.
-
Fang J, Huang X, Han X, Zheng Z, Hu C, et al. (2020) Endothelial progenitor cells promote viability and nerve regenerative ability of mesenchymal stem cells through PDGF-BB/PDGFR-beta signaling. Aging (Albany NY) 12(1): 106-121.
-
Zheng X, Zhang W, Wang Z (2019) Simvastatin preparations promote PDGF-BB secretion to repair LPS- induced endothelial injury through the PDGFRbeta/ PI3K/Akt/IQGAP1 signalling pathway. J Cell Mol Med 23(12): 8314-8327.
-
Zhang JM, Feng FE, Wang QM, Zhu XL, Fu HX, et al. (2016) Platelet-Derived Growth Factor-BB Protects Mesenchymal Stem Cells (MSCs) Derived From Immune Thrombocytopenia Patients Against Apoptosis and Senescence and Maintains MSC-Mediated Immunosuppression. Stem Cells Transl Med 5(12): 1631-1643.
-
Pan CH, Chen CJ, Shih CM, Wang MF, Wang JY, et al. (2019) Oxidative stress-induced cellular senescence desensitizes cell growth and migration of vascular smooth muscle cells through down-regulation of platelet-derived growth factor receptor-beta. Aging (Albany NY) 11(19): 8085-8102.
-
Yu YC, Yang PM, Chuah QY, Huang YH, Peng CW, et al. (2013) Radiation-induced senescence in securin- deficient cancer cells promotes cell invasion involving the IL-6/STAT3 and PDGF-BB/PDGFR pathways. Sci Rep 3: 1675.
-
Yu T, Chen X, Zhang W, Liu J, Avdiushko R, et al. (2016) KLF4 regulates adult lung tumor-initiating cells and represses K-Ras-mediated lung cancer. Cell Death Differ 23(2): 207-215.
-
Panatta E, Lena AM, Mancini M, Affinati M, Smirnov A, et al. (2018) Kruppel-like factor 4 regulates keratinocyte senescence. Biochem Biophys Res Commun 499(2): 389-395.
-
Yet SF, McA’Nulty MM, Folta SC, Yen HW, Yoshizumi M, et al. (1998) Human EZF, a Kruppel-like zinc finger protein, is expressed in vascular endothelial cells and contains transcriptional activation and repression domains. J Biol Chem 273(2): 1026-1031.
-
Liu Y, Sinha S, McDonald OG, Shang Y, Hoofnagle MH, et al. (2005) Kruppel-like factor 4 abrogates myocardin- induced activation of smooth muscle gene expression. J Biol Chem 280(10): 9719-9727.
-
Salmon M, Gomez D, Greene E, Shankman L, Owens GK, et al. (2012) Cooperative binding of KLF4, pELK-1, and HDAC2 to a G/C repressor element in the SM22alpha promoter mediates transcriptional silencing during SMC phenotypic switching in vivo. Circ Res 111(6): 685-696.
-
Yoshida T, Kaestner KH, Owens GK (2008) Conditional deletion of Kruppel-like factor 4 delays downregulation of smooth muscle cell differentiation markers but accelerates neointimal formation following vascular injury. Circ Res 102(12): 1548-1557.
-
Liang S, Yu H, Chen X, Shen T, Cui Z, et al. (2017) PDGF- BB/KLF4/VEGF Signaling Axis in Pulmonary Artery Endothelial Cell Angiogenesis. Cell Physiol Biochem 41(6): 2333-2349.
-
Haskins RM, Nguyen AT, Alencar GF, Billaud M, Kelly Goss MR, et al. (2018) Klf4 has an unexpected protective role in perivascular cells within the microvasculature. Am J Physiol Heart Circ Physiol 315(2): 402-414.
-
Zhou G, Hamik A, Nayak L, Tian H, Shi H, et al. (2012) Endothelial Kruppel-like factor 4 protects against atherothrombosis in mice. J Clin Invest 122(12): 4727- 4731.
-
Sangwung P, Zhou G, Nayak L, Chan ER, Kumar S, et al. (2017) KLF2 and KLF4 control endothelial identity and vascular integrity. JCI Insight 2(4): e91700.
-
Niu N, Xu S, Xu Y, Little PJ, Jin ZG, et al. (2019) Targeting Mechanosensitive Transcription Factors in Atherosclerosis. Trends Pharmacol Sci 40(4): 253-266.
-
Deaton RA, Gan Q, Owens GK (2009) Sp1-dependent activation of KLF4 is required for PDGF-BB-induced phenotypic modulation of smooth muscle. Am J Physiol Heart Circ Physiol 296(4): 1027-1037.
-
Yu K, Zheng B, Han M, Wen JK (2011) ATRA activates and PDGF-BB represses the SM22alpha promoter through KLF4 binding to, or dissociating from, its cis-DNA elements. Cardiovasc Res 90(3): 464-474.
-
Zhao Y, Lv W, Piao H, Chu X, Wang H, et al. (2014) Role of platelet-derived growth factor-BB (PDGF-BB) in human pulmonary artery smooth muscle cell proliferation. J Recept Signal Transduct Res 34(4): 254-260.
-
Khosla S, Farr JN, Tchkonia T, Kirkland JL (2020) The role of cellular senescence in ageing and endocrine disease. Nat Rev Endocrinol 16(5): 263-275.
-
Rane MJ, Zhao Y, Cai L (2019) Krupsilonppel-like factors (KLFs) in renal physiology and disease. EBioMedicine 40: 743-50.
-
Kumekawa M, Fukuda G, Shimizu S, Konno K, Odawara M, et al. (2008) Inhibition of monocyte chemoattractant protein-1 by Kruppel-like factor 5 small interfering RNA in the tumor necrosis factor- alpha-activated human umbilical vein endothelial cells. Biol Pharm Bull 31(8): 1609-1613.
-
Botella LM, Sánchez Elsner T, Sanz Rodriguez F, Kojima S, Shimada J, et al. (2002) Transcriptional activation of endoglin and transforming growth factor-beta signaling components by cooperative interaction between Sp1 and KLF6: their potential role in the response to vascular injury. Blood 100(12): 4001-4010.
-
Helbing T, Volkmar F, Goebel U, Heinke J, Diehl P, et al. (2010) Kruppel-like factor 15 regulates BMPER in endothelial cells. Cardiovasc Res 85(3): 551-559.
-
Simmons RD, Kumar S, Jo H (2016) The role of endothelial mechanosensitive genes in atherosclerosis and omics approaches. Arch Biochem Biophys 591: 111-131.
-
Atkins GB, Wang Y, Mahabeleshwar GH, Shi H, Gao H, et al. (2008) Hemizygous deficiency of Kruppel-like factor 2 augments experimental atherosclerosis. Circ Res 103(7): 690-693.
-
Sweet DR, Fan L, Hsieh PN, Jain MK (2018) Kruppel-Like Factors in Vascular Inflammation: Mechanistic Insights and Therapeutic Potential. Front Cardiovasc Med 5: 6.
-
Yoshida T, Yamashita M, Iwai M, Hayashi M (2016) Endothelial Kruppel-Like Factor 4 Mediates the Protective Effect of Statins against Ischemic AKI. J Am Soc Nephrol 27(5): 1379-1388.
-
Zhang J, Zhang H, Chen Y, Fu J, Lei Y, et al. (2019) Plateletderived growth factor D promotes the angiogenic capacity of endothelial progenitor cells. Mol Med Rep 19(1): 125-132.
-
Aoyagi M, Fukai N, Ogami K, Yamamoto M, Yamamoto K, et al. (1995) Kinetics of 125I-PDGF binding and down- regulation of PDGF receptor in human arterial smooth muscle cell strains during cellular senescence in vitro. J Cell Physiol 164(2): 376-84.
-
Vion AC, Kheloufi M, Hammoutene A, Poisson J, Lasselin J, et al. (2017) Autophagy is required for endothelial cell alignment and atheroprotection under physiological blood flow. Proc Natl Acad Sci U S A 114(41): 8675-8684.
-
Ying HZ, Chen Q, Zhang WY, Zhang HH, Ma Y, et al. (2017) PDGF signaling pathway in hepatic fibrosis pathogenesis and therapeutics (Review). Mol Med Rep 16(6): 7879- 7889.
-
Shankman LS, Gomez D, Cherepanova OA, Salmon M, Alencar GF, et al. (2015) KLF4-dependent phenotypic modulation of smooth muscle cells has a key role in atherosclerotic plaque pathogenesis. Nat Med 21(6): 628-637.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions