Interaction of Inflammatory Molecules in Tuberculosis and Sars-Cov-2 Infection
The Hyper-inflammatory immune response is a chief reason for disease severity and mortality in patients. Generally, macrophages and dendritic cells sense and react to microbial or viral invasion by making inflammatory molecules that remove pathogens and help in tissue repair. During tuberculosis infection, alveolar macrophages activate alveolar dendritic cells, which move to lymph nodes. In lymph nodes, the proliferation of CD4+ T cells, CD8+ cells and γδ T cells occur. Mycobacterium tuberculosis (MTB) bacteria further modify the host's immune system for their long survival period. Meanwhile, Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) infects alveolar macrophages and releases inflammatory cytokine to produce and activate T-cells. MTB and SARS-CoV-2 infections together lead to an increase in the rate of pathogenesis. This type of co-infection in macrophages causes the production of pro- and anti-inflammatory cytokines, which further play an important role in immune-pathogenesis. However, the abnormal or dysregulated response of macrophages leads to harmful effects of the host, as observed in the macrophage activation syndrome induced by severe infections, including the virus SARSCoV-2. Unlike macrophages, dendritic cells (DCs) act as antigen presenting cells. They connect innate and adaptive immunity cells. They are susceptible to cytokine-mediated activation and lead to cytokine production. The cytokine Interleukin-6 (IL6) is an important and unique molecule that can act pro- as well as anti-inflammatory and helps in the development and differentiation of macrophages associated with numerous inflammatory diseases. In this review paper, we have emphasized the vital pathological role of macrophages, dendritic cells and IL-6 in tuberculosis and SARS-CoV-2 infection and prospective therapeutic strategies based upon IL-6 as the main target for preventing the cytokine storm and associated organ failure.
Introduction
Tuberculosis (TB) is already presented as a pandemic over many years. World Health Organization (WHO) declared it as a global health emergency in 1993 [1]. Tuberculosis is among the top 10 causes of death worldwide because of the single infectious agent Mycobacterium tuberculosis. Annually, millions of people continue to fall sick with TB. Around 10 million people were affected by this disease in 2020, and 1.6 million died worldwide [2]. Out of 10 million new cases, 2.74 million cases were from India alone, making it the highest TB burden country. India accounts for more than 27% of total global incidence, the highest among the top 30 high TB burden countries globally. Despite numerous TB elimination programs, around one-fourth of total global cases have been reported from India alone.
Recently, Coronavirus (COVID-19) has emerged as a highly destructive disease prevalent worldwide. It was first reported from Wuhan City of Hubei Province of China in December 2019 [3]. The causative agent responsible for COVID-19 is a virus named Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), a member of the Coronaviridae family, which also includes Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) and the Middle East Respiratory Syndrome Coronavirus (MERS- CoV) [4]. The WHO has declared COVID-19 as a pandemic on March 11, 2020. It is the most damaging disease in recent history. Old age people are more susceptible to this disease due to their weakened immune system in terms of adaptive immunity. There have been 274,555,585 confirmed cases of COVID-19 and 5,367,175 deaths reported (as of December 19, 2021). A total of 8,337,664,456 vaccine doses have been given globally (as of December 15, 2021) [2]. India reported its first case on January 30, 2020. Since then, there have been 34,740,275 confirmed cases of COVID-19 with 477,422 deaths (as of December 19, 2021), and a total of 1,294,608,045 vaccine doses have been given (as of December 6, 2021).
Common and different features of Tuberculosis and COVID-19
Tuberculosis and COVID-19 share common immunopathology [5]. Even though COVID-19 and TB are quite similar in some aspects, some differences are also present in (Table 1). For example, their causative agents are different. Still, they show similar symptoms like fever, cough, breathlessness and weakness with varying severity in both cases [6]. Both are contagious and infectious diseases transmitted from one individual to another through aerosol or close contact and mainly affect the lungs. TB is transmitted through droplet nuclei of aerosols transferred by an infected person with MTB. After inhalation of MTB, the contaminated aerosol host becomes infected. Depending upon the host, bacterial and environmental factors, MTB infection may lead to the development of active TB in the form of pulmonary TB (PTB) or extra-pulmonary TB (EPTB). However, the incubation period of TB is longer with the slow onset of the disease. Depending upon drug resistance, active TB can also advance to multidrug-resistant (MDR) or extensively drug-resistant (XDR) TB forms causing additional threats to mankind. It has been observed that among MTB infected patients, about 10% lead towards active TB disease and roughly 90% of infected individuals maintain latent TB infection [7, 8]. The vulnerability of that 10% of individuals to TB mainly depends upon complex associations between MTB and the host immune system [9]. After going through the current situation of TB, the theme of world TB day 2021 is “the clock is ticking”, which means the COVID-19 pandemic has delayed the curing progress rate of TB. So this dreadful situation has to be overcome by effective and preventive measures.
| TB | COVID-19 | |
|---|---|---|
| Exposure | Ancient | Recent |
| Global status | Around 10 million cases | Around 43 million cases |
| Mode of transmission | Aerosols | Droplets, aerosols, fomites and surface contact |
| Mode of transmission | Contagious (slow) | Contagious (rapid) |
| Incubation period | Longer with a slow onset of disease | Ranges from 1-14 days most commonly being around 5 days |
| Symptoms | Cough with mucous or blood | Fever or nippy |
| Symptoms | Cough that lasts more than 2 months | Shortness of breath |
| Symptoms | Chest pain and fatigue | Fatigue, headache, nausea, and congestion |
| Symptoms | Weight loss and loss of appetite | Loss of taste and smell |
| Most affected organ | Lungs | Lungs (leads to organ failure with the severity of disease) |
| Latent form | Present | Absent |
| Comorbidities | Cancer | Cancer |
| Comorbidities | Chronic lung disease | Chronic kidney disease |
| Comorbidities | Smoking | Chronic lung disease |
| Comorbidities | Alcohol use | Obesity |
| Comorbidities | HIV | Sickle cell disease |
| Comorbidities | Immunocompromised system | Immunocompromised system |
| Comorbidities | Type 2 diabetes mellitus | Type 2 diabetes mellitus |
| Available vaccine | Yes (BCG) | Yes (COVAXIN, COVISHIELD, PFIZER-BioNTech, SPUTNIK V etc.) |
| Availability of cure | Yes | Yes |
| Drug resistance | Yes (MDR, XDR) | The mutant form of the virus is there but data is not available on, drug resistance |
| Economic impact and stress on the health system | Huge but slow | Huge but rapid |
Table 1: Similarity and differences between TB and COVID-19.
On the other side, SARS-CoV-2 can transmit disease mainly through droplets and fomites but can also be through aerosols, and it also spreads through surface contamination [10]. Symptoms may appear 2-14 days after exposure, with a median incubation period of 5 days and a latent period is not present in COVID-19. An infected person with SARS-CoV-2 can further transmit the infection to persons in close contact. Still, an infected TB person can infect further 1-4 individuals. Risk factors responsible for both COVID-19 and TB are advanced age, diabetes, smoking, chronic kidney disease, chronic respiratory disorders and immune-compromised state. If treatment of TB is disrupted in any case, it leads to poorer treatment outcomes and chances of COVID-19 disease severity also increase [11, 12]. Although research is going on to find an association between these two comorbidities, full- proof data is not available to tell whether active or previously treated TB can enhance the severity of COVID-19 or not. However, the various epidemiological studies are also going on a large scale to find out the association between TB and COVID-19 [13].
Host immune responses in TB and COVID-19
With respect to host immune response, macrophages and dendritic cells (DCs) provide the first line of defence by their recruitment at the very first site of infection and have multiple roles in the innate immune system. They can phagocytose bacteria and viruses to initiate and trigger an immune response [14]. They interact with bacteria and viruses by employing various cell surfaces and nuclear receptors. Interactions of bacterial pathogen-associated molecular patterns (PAMPs) with these host-cell receptors result in bacteria uptake and entanglement in the phagosomes of macrophages and DCs [15, 16, 17, 18, 19]. Airway epithelial cells (AEC), macrophages, natural killer cells (NK), neutrophils, dendritic cells (DCs), mast cells and the complement system are the vital performer of immunity tasks. When breathing in, MTB comes across through the first line of defence comprising air route epithelial cells (AECs), neutrophils, monocytes, and dendritic cells [20, 8]. If it succeeds in abolishing MTB rapidly, the infection terminates. Otherwise, phagocytes get infected, and MTB proliferate inside the cells, then they initially cause clinical indications and progression of disease depending on the complex relationship between bacterial and host factors [21] (Figure 1).
Based on several pieces of evidence, it has been confirmed that people affected with COVID-19 could lead to cytokine storm syndrome where there is an increase in inflammatory cytokines [3, 22]. This syndrome causes lung damage or injury in severely ill-defined COVID-19 patients. During SARS-CoV-2 infection, there is the secretion of IL-6 in macrophages induces apoptosis in lymphocytes. Along with IL-6 other inflammatory cytokines such as IL-12, IL-8, TGF-β, TNF-α, IL-1β, IFN-γ and various chemokines such as CCL2, CXCL10 and CXCL9 are associated with macrophage activation syndrome and adult respiratory distress syndrome [23]. It has been observed that there is a decrease in T-cells in severe cases and an increase in neutrophil and monocyte- lymphocyte ratio [24] (Figure 2).
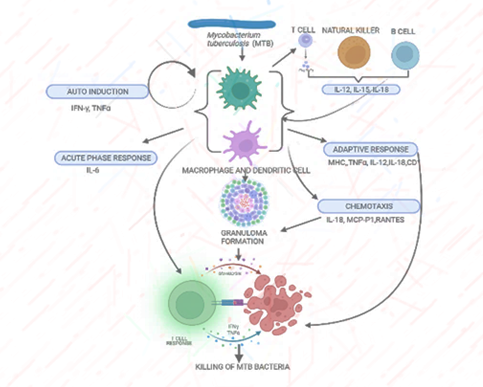
Figure 1: Phagocytic cells upon activation by bacterial infection show several Inflammatory responses. Immune recognition of Mycobacterium tuberculosis by macrophages and dendritic cells is followed by an inflammatory response with cytokine production. Initial events in this cellular response include nonspecific host defense mechanisms, which may lead to early killing or containment of infection.
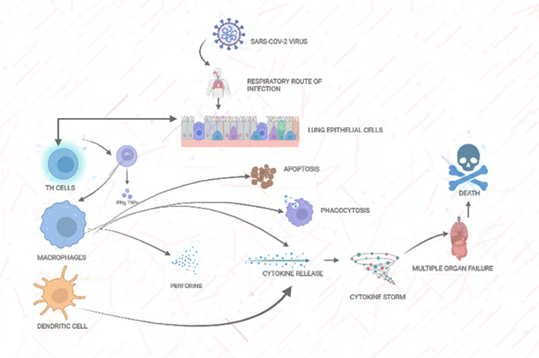
Figure 2: Phagocytic cells upon activation by viral infection show several Inflammatory responses. Immune recognition of SARS-COV-2 by macrophages and dendritic cells is followed by an inflammatory response with the cytokine release and cytokine storm production. Initial events in this cellular response include nonspecific host defense mechanisms, which may lead to early killing or containment of infection.
Role of macrophages in TB and COVID-19
Macrophages are innate immune cells that are the part of first line of defense. Still, they are only capable of this if the ratio of cells and invading microbes favour the situation, i.e. if the number of cells is greater than the load of microbes, they can prevent the infection [21, 25]. Otherwise, they promote pathogenesis because the gradual proliferation of the MTB carried out as macrophages serve as the initial niche for microbes. During the latent phase of infection, the number increase to hundreds inside the phagosomes. Different categories of death-related factors that resist macrophages’ attempts in suppressing the pathogen expressed by MTB. Various activities inhibited by MTB in the host are apoptosis, autophagy, intracellular trafficking, reactive oxygen species (ROS) and toxic metal [26]. Macrophages have IFN-γ as the main player to restrict MTB, which is controlled by vitamin D to perform its functions [27].
A variety of genes like hCAP-18 encodes cathelicidinsiscationic antibacterial peptide, which stimulates gene transcription related to autophagy. Credit for this phenomenon goes to vitamin D, which enhances macrophage maturation [28]. With the help of early secretory antigenic target (ESAT-6) protein secretion (ESX), MTB can escape from the immune system by exporting anti- immunogenic transport protein. IFN-γ axis is disturbed by secretion of ESX-system, due to which IFN-γ makes so much effort to promote the phagolysosomal activity so that they can tackle with MTB. MTB proliferation occurs in the lack of vitamin D [29]. The role of Nitric oxide (NO) in humans is comparatively low with respect to animal models in macrophages [26].
Reactive oxygen species (ROS) production on the phagosome membrane is regulated by the catalytic activity of nicotinamide adenine dinucleotide phosphate (NADPH- oxidase 2). Mutation in the catalytic subunit of NADPH- oxidase 2 has been seen in TB susceptible patients, showing impaired ROS production [30, 31]. MTB controls the NADPH- oxidase component and its tasks by interacting with small guanosinetriphosphatases (GTPases)through nucleoside diphosphate kinase (Npk) [32]. There is a battleground in macrophages for the MTB to sort metals and cells [33]. The macrophages recruit an inaccurate load of zinc and copper; their high concentrations are poisonous to MTB. MTB displays protective signalling processes that counteract such situations like an inappropriate load of metals and oxidation; up-regulation of cation-transporting P-type ATPase (ctpC) gene restricts metal poisoning [34]. To counterbalance this, macrophages put efforts to prevent the entry of metal nutrients into the MTB. These nutrients are manganese and iron.
There are different strategies used by immune cells to combat the virus after its entry through viral membrane protein through Fc gamma receptors present on activated immunocytes like macrophages, neutrophils, monocytes and natural killer cells. This leads to the release of perforins and granzymes containing granules which kill the infected cells [35, 36]. This phenomenon is considered to be ADCC which is the efficient participation of immune response towards different viruses and various studies examined during RSV infection and during anti-RSV vaccine design. Antibodies that are formed against SARS coronavirus show a two-edged result on the progression of infection and characterize both favourable and objectionable events through the assistance of immune system mechanisms, including NK cells facilitated by antibody-dependent cell cytotoxicity mechanism and macrophages, specifically alveolar macrophages by Fc gamma receptor (FcγRs) containing cells, monocytes-derived infiltrating, macrophages, monocytes, and complement– antibody complex pathways [37]. Phagocytic cells through SARS-CoV specific show a major role in the clearance of virus- infected pulmonary cells by antibodies alveolar macrophages and monocyte-derived infiltrating macrophages. Tissue- damaging and immunopathological complications may cause due to extreme Fc mediated phenomenon while eliminating infected cells.
Antibody-dependent enhancement (ADE) happens when antiviral antibodies cannot neutralize the virus. ADE cause increases the infection of cells that have FCR through the interactions between kinds of receptors and the virus- antibody immune complex [22]. The exciting fact is that ADE may be tempted by both non-neutralizing and neutralizing antibodies [38]. Studies reveal that ADE might arise during viral infections with Zika, Ebola, dengue, influenza, and HIV. Few disorders are the results of novel coronavirus disease. Studies showed that the severity of the disease is caused by passive approaches after the production of antibodies due to the prior immunity [39]. It has been described in various studies that macrophages have a significant role in the escalating immunopathogenesis of SARS-CoV-2 by enhancing ADE owing to their FCR. It has been projected that alveolar macrophages might participate in worsening COVID-19 by facilitating ADE at FcγRIIA(CD32A) high level. The production of inflammatory cytokines is induced by IgG binding to the FcγRIIA [40]. Surprisingly, a rise in the number of macrophages by replication in antibodies against SARS- CoV-2 has been seen under in vitro conditions [41].
Macrophages are crucial infectious innate immune cells against any infectious agent [42, 43]. They are extremely versatile innate cells that can act differentially in function and morphologically to evade pathogenic organisms, cancerous cells, virus-infected cells, and other macrophages that help heal parasite infections [44, 45]. A balanced immune system is required to activate both kinds of macrophages, M1 and M2, through complex and diverse innate cells [46, 47]. During ageing and chronic inflammatory diseases, macrophages may shift to a more M2-like phenotype [48, 49]. Importantly, nearly all identified high-risk factors for severe COVID19 disease, like cardiovascular disease, diabetes, age, chronic obstructive pulmonary disease, and smoking, generally share a shift from more M1 to more M2 phenotype and function [50]. Classical activation of M1 macrophages is induced by LPS/IFN-γ exposure, while alternately activated M2 macrophages are stimulated by IL-4, IL-10, IL-13 and glucocorticoids [51]. The activation of innate immune cells such as macrophages can be heavily influenced by the character of the T cell response and, in particular, the cytokines produced by T cells during infection [52]. SARS-CoV replication has previously been shown in human peripheral monocytes and macrophages, with varying efficacy. Importantly, the infection efficiency was shown to be donor-dependent, with 100% infection in some and less than 5% in others [53].
During vaccination to rhesus macaques with the SARS- CoV-2 vaccine, which is constructed keeping in view anti-S IgG, it is discovered that there is crucial relation between SARS-CoV-2 development of severity of disease, immune response and release of antibodies against it. It is found that vaccine before clearing the viral load results in acute lung injury via staffing of pro-inflammatory alveolar macrophages. They lead to the production of a huge amount of MCP-1 and IL-8 chemokines, which cause swelling [54, 55]. Numerous studies confirmed similar outcomes in which lung damage was worse in mice and African green monkeys after SARS coronavirus vaccination [56, 57]. A similar provocative response outline has been observed in several phases of COVID-19 cases.
Role of dendritic cells in TB and COVID-19
Dendritic cells (DCs) are operationally active one of the main cells is connecting, innate and adaptive immune retaliation via a distinctive role in arresting, operating and antigen presenting to T cells in the context of both MHC class I and II. Macrophages get activated by T cells, secrete tumour necrosis factor (TNF) and interferon (IFN-gamma) to kill or eliminate micro-bacterium; besides this, macrophages play significant participation in mobilizing of cells by releasing pro-inflammatory cytokines IL-6 and IL-1 at the area of disease or infection caused by bacterial invasion. These cytokines have potential activity on the lymphocytes cluster of differentiation (CD4). DCs have controversial status in controlling the TB disease. Few studies reveal whether these cells nourish the immunity against MTB or sometimes MTB modulate their action, decreasing T-cells’ function. Quickly DCs act as a niche or reservoir for MTB [58, 59, 60, 61].
CD209 provides a specific route to MTB by binding with 3-grabbing non-integrin receptor (DC-SIGN) inside the DCs and acts as a specific intercellular adhesion molecule. Under normal internal eco-friendly conditions, CD209 serve as a receptor for CD54 and favours DC movements in the cell. By coupling with the lipoarabinomannan mannose (ManLAM), a component of MTB membrane, CD209 allow its penetration into the cell, which enhances IL-10 secretion and reduces IL-12 production, which leads to alternation of DCs activity, disguising the pathogen and disturbed stimulating phenomenon that represents antigen by T lymphocytes. MTB gets benefitted from this and grows rapidly. The vaccination strategy against MTB uses the DC-SIGN mechanism as a potential target to promote immunity. It also balances inflammatory status by releasing IL-12 in the host and protecting it from infection and tissue pathogenesis. Some studies have reported the fate of DCs maturation, which depends on the type of receptors during recognition. Interactions of MTB with TLR result in DC activation characterized by high IL-12 secretion during early infections. On the other hand, interactions with DCSIGN prevent DC activation by blocking TNF-kB activation, resulting in increased secretion of IL-10 [62, 63, 64, 65, 66, 67].
Dendritic cells are widely distributed in the respiratory tract, and various pathogens attack them. DCs act like antigen presenting cells having a role in antiviral immune responses and have a role in SARS-CoV infection. DCs mainly express ACE2 on their receptors and lead to SARS-CoV infection. These cells played an important role in the pathogenesis of SARS. There is a significant increase in the production of RANTES and their specific receptors such as CCR-1, CCR-3 and CCR-5 mRNA in DCs infected with SARS-CoV. When there is high production of RANTES, it leads to the death of SARS patients. More studies relevant to this topic are required to reduce RANTES production [68, 69, 70, 71]. Another factor is there, which is known as TRAIL gene expression in adult and cord blood DCs after SARS-CoV infection. It leads to destroyer DCs gene expression. So based on this study, further gene expression studies are required to combat increased TRAIL gene expression in DCs to reduce the risk of lymphopaenia. Due to various complex mechanisms, the development and function of DCs need to be revealed further.
During SARS-CoV-2 infection, DC cells increase DC-SIGN and furin expressions, which ultimately further causes a high rate of infection. Advanced age can also negatively impact DCs number. So both factors, i.e. SARS-CoV infection and the ageing process, promote dendritic cell dysfunction or death, which causes high viral infection load, cytokine release syndrome, and sepsis. The diseased state of DCs in the form of dysfunction and death may be responsible for a bad outcome in COVID-19 disease leading to more complications and deaths. Patients already affected with COVID-19 lead to viral sepsis, and there is a decrease in the number of dendritic cells; moreover, the body’s functional impairment is also there [72, 73, 74, 75, 76, 77, 78, 3]. In view of this, deep knowledge and a better understanding of the immune functions mediated by DCs should be essential to combat the adverse effects of dendritic cell-mediated treatment and vaccines.
Nowadays, many drugs emerge as therapeutic agents for COVID-19. The names of the drugs are remdesivir, hydroxychloroquine, chloroquine, azithromycin, lopinavir and ritonavir. Dendritic cell-based vaccines are also widely used to cure tumours and pathogens. Vaccines for SARS-
CoV-2 infection based on DC are under clinical trial and development for future use [79, 80, 81, 82]. Although, new antiviral vaccines are now available for the treatment of this disease, such as COVAXIN and COVSHIELD, which are quite effective against it.
Role of IL-6 in TB and COVID-19
Human IL-6 consists of 212 amino acids, counting a 28-amino-acid signal peptide, and its gene is present on chromosome 7p21. Although the central protein is ∼20 kDa, natural IL-6 is 21–26kDa. The cytokine IL-6 is a unique molecule that can act as pro as well as anti-inflammatory. IL-6 may be harmful in mycobacterial infections, as it inhibits the production of TNF-α and promotes in vitro growth of M. avium. The production of anti-inflammatory cytokines helps to regulate this inflammatory process; nevertheless, this can give a chance to MTB for growth in host tissue. It can be speculated that genetically determined high and low production of cytokines may render an individual vulnerable or resistant to MTB infection depending upon the nature of cytokines. Other reports support a protective role for IL-6; IL- 6-deficient mice display increased susceptibility to infection with MTB which seems related to a deficient production of IFN-γ early in the infection before adaptive T-cell immunity has fully developed. IL-6 has shown a peculiar association with TB, where it can render the host susceptible to TB as it interferes with TNF-α production. Its deficiency has also been reported to be making mouse models vulnerable to MTB infection. IL-6 is needed for optimal T-cell development [83, 84, 85, 86, 87, 88]. Indeed, high levels of IL-6 cytokine are produced in response to MTB infection [59], and its role seems especially critical when the bacterial burden is high.
Macrophages and monocytes produce other inflammatory phenomena during Covid-19, known as cytokine storm, which is known as cytokine release syndrome (CRS) and hyper-cytokinemia, which are extremely stimulated due to infectious or non-infectious agents and produce a decent number of cytokines that participate in hyper inflammation through prompting and enrolling of leucocytes [76, 89, 90, 91, 92].
Cytokine storm has chief participation in the severity of Covid-19 and lung damage in the diseased patients [93, 94]. Series of cytokines released during the event, which include IL-17, IL-9, IL-7, βIL-2, IL-1, IL-10, TNF-α, G-CSF, GM-CSFIFN-γ, MIP1A, MCP1, MIP1B, CXCL8 and CXCL10 in the recently found new corona virus-diseased individuals which can lead to the immunopathological development of COVID-19. CRS is closely connected with macrophage activation syndrome (MAS), characterized by several clinical indications such as liver failure, hyperferritinemia, pancytopenia, neurological disorders and coagulopathy. Excessive production of differentiated macrophages leads to MAS, resulting in various clinical manifestations like hypercytokinemia and hemophagocytosis [95, 96].
The harshness of the disease Covid- 19 is connected with the elevated levels of chemokines and cytokines, which are pro-inflammatory in nature. These include IL-7, IL-10, IL-2, IL-6, TNF (tumour necrosis factor), MCP1 (monocyte chemoattractant protein-1), G-CSF (granulocyte colony- stimulating factor), CXCL10 (CXC-chemokine ligand) [97, 98, 99, 100, 101, 102]. Studies suggested that IL-6 is vastly correlated with COVID-19 severity above all cytokines, comparable IL-6 levels in the blood were noticed between COVID-19 fighters and non-survivors [82, 103]. It is one of the causes of disease mortality due to cytokine release syndrome (CRS), also known as cytokine storm [23, 104]. Cytokine storm is an exciting link in COVID-19 infectious individuals. It is found that patients with the disease severity have high IL-6 levels and show hyperactive inflammatory immune response, which causes pulmonary inflammation, lung injury, and multiple organ failure, leading to the death of the patient (Channappanavar & Perlman). A recent study disclosed that elevated levels of pro-inflammatory mediator cytokines in serum such as IL6, IL1β, IFNγ, IL12, IP10 and MCP1 were associated with clinical lung manifestation in SARS patients. These were found in severe covid patients, which is responsible for the cause of immune cell response of activated T-helper-1 (Th1) [105, 106, 107]. IL-6 is a key cytokine that helps develop and differentiate macrophages and is associated with numerous inflammatory diseases. Moreover, SARS-CoV-2 diseased people had minute levels of suppressor of cytokine signalling-3 and low count of CD8+ and CD4, which controls negative feedback mechanism of IL-6and TNF [100, 108, 109, 110]. In addition, new studies presented that higher IL-6, IL- 10 and CRP levels were weightier than other cytokines in the critical group of COVID-19 infected individuals’ patients.
Role of BCG vaccine in TB and COVID-19
There is some evidence showing that people who have latent TB infection could lead to severe SARS-CoV-2 infection. Patients with COVID-19 disease could lead to a worse infection if left untreated because they were previously also infected with TB disease, in which their lung is more affected. As we all know, the Bacillus Calmette-Guerin (BCG) vaccine is useful in TB disease. BCG vaccination protects newborns and infants from pulmonary TB. It also protects against other infectious diseases, so it is proposed that BCG vaccination could also provide immunity against COVID-19 disease. In many countries where continuous use of BCG vaccine in neonates was given in that case, the rate of reported cases of COVID-19 infections was less in number. Due to insufficient evidence, BCG is not recommended to prevent COVID-19. Some epidemiological studies show an association between the BCG vaccine and COVID-19 also irrespective of TB
disease. But these studies are not sufficient to establish causality between BCG vaccination and protection from severe COVID-19 [111, 112, 113, 11, 12].
Conclusion
According to the present situation, We should always keep in mind that “Prevention is the key to success” and rebuild our medical tools to tackle this pandemic situation to combat this menace of disease. Macrophages and dendritic cells are crucial immune cells with a broad range of defensive phenomena against numerous micro-pathogenic agents such as bacteria and viruses. In life-threatening emergencies like COVID-19, these macrophages induce pathogenic activities like elevating inflammatory cytokines and chemokines and excessive interaction with the immune cells, which cause severe tissue damage and disease severity.
The immune system’s response in relation to macrophages, dendritic cells, and IL-6 has to be determined. Proper monitoring should be done on patients with COVID-19 disease and TB patients. During the treatment of patients, the higher levels of IL-6 should be regularly monitored. IL-6 may be the new earmark for the effective treatment of COVID-19. We all know that this disease mainly causes pulmonary and extra-pulmonary organ damage associated with cytokine storms. So an effective therapeutic drug has to be designed by targeting IL-6 against COVID-19 disease.
People with TB are not expected to get COVID‐19, but pre‐existing TB has a higher chance of infection with COVID‐19 and complications associated with it. More studies are needed to better understand this and in addition to this enough epidemiological data with evidence is not available and it is not sufficient to recommend the use of BCG to curb COVID-19.
Conflict of Interest
The authors declare no conflict of interest.
Funding
NA
References
-
Gagneux S (2018) Ecology and evolution of Mycobacterium tuberculosis. Nature Reviews Microbiology 16(4): 202-213.
-
World Health Organization (2021) WHO operational handbook on tuberculosis. Module 3: diagnosis-rapid diagnostics for tuberculosis detection 2021 update.
-
Wang Q, Qiu Y, Li JY, Zhou ZJ, Liao CH, et al. (2020) A unique protease cleavage site predicted in the spike protein of the novel pneumonia coronavirus (2019- nCoV) potentially related to viral transmissibility. Virol Sin 35(3): 337-339.
-
Lu R, Zhao X, Li J, Niu P, Yang B, et al. (2020) Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. The lancet 395(10224): 565-574.
-
Mousquer GT, Peres A, Fiegenbaum M (2021) Pathology of TB/COVID-19 co-infection: The phantom menace. Tuberculosis 126: 102020.
-
Song WM, Zhao JY, Zhang QY, Liu SQ, Zhu XH, et al. (2021) COVID-19 and Tuberculosis Coinfection: An Overview of Case Reports/Case Series and Meta-Analysis. Frontiers in medicine 8: 657006.
-
Houben RMGJ, Dodd PJ (2016) The global burden of latent tuberculosis infection: a re-estimation using mathematical modelling. PLoS medicine 13(10): e1002152.
-
O’Garra A (2013) Systems approach to understand the immune response in tuberculosis: an iterative process between mouse models and Human Disease, pp: 1-5.
-
Azad AK, Sadee W, Schlesinger LS (2012) Innate immune gene polymorphisms in tuberculosis. Infect Immun 80(10): 3343-3359.
-
van Doremalen N, Bushmaker T, Morris DH, Holbrook MG, Gamble A, et al. (2020) Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1. N Engl J Med 382(16): 1564-1567.
-
World Health Organization (2020a) Guidance for the surveillance of drug resistance in tuberculosis: 6th(Edn.).
-
World Health Organization (2020b) WHO consolidated guidelines on tuberculosis. Module 3: Diagnosis-Rapid diagnostics for tuberculosis detection.
-
Tadolini M, Codecas, LR, García-García JM, Blanc FX, Borisov S, et al. (2020) Active tuberculosis, sequelae and COVID-19 co-infection: first cohort of 49 cases. European Respiratory Journal 56(1): 2001398.
-
Gordon S, Taylor PR (2005) Monocyte and macrophage heterogeneity. Nature reviews immunology 5(12): 953- 964.
-
Ernst JD (1998) Macrophage receptors for Mycobacterium tuberculosis. Infect Immun 66(4): 1277- 1281.
-
Kang PB, Azad AK, Torrelles JB, Kaufman TM, Beharka A, et al. (2005) The human macrophage mannose receptor directs Mycobacterium tuberculosis lipoarabinomannan-mediated phagosome biogenesis. The Journal of experimental medicine 202(7): 987-999.
-
Underhill DM, Ozinsky A, Smith KD, Aderem A (1999) Toll-like receptor-2 mediates mycobacteria-induced proinflammatory signaling in macrophages. Proceedings of the National Academy of Sciences 96(25): 14459- 14463.
-
Court N, Vasseur V, Vacher R, Frémond C, Shebzukhov Y, et al. (2010) Partial redundancy of the pattern recognition receptors, scavenger receptors, and C-type lectins for the long-term control of Mycobacterium tuberculosis infection. J Immunol 184(12): 7057-7070.
-
Basu J, Shin DM, Jo EK (2012) Mycobacterial signaling through toll-like receptors. Front Cell Infect Microbiol 2: 145.
-
Middleton AM, Chadwick MV, Nicholson AG, Dewar A, Groger RK, et al. (2002) Interaction of Mycobacterium tuberculosis with human respiratory mucosa. Tuberculosis 82(2-3): 69-78.
-
Martino MD, Lodi L, Galli L, Chiappini E (2019) Immune response to Mycobacterium tuberculosis: a narrative review. Front Pediatr 7: 350.
-
Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, et al. (2020) COVID-19: consider cytokine storm syndromes and immunosuppression. The Lancet 395(10229): 1033-1034.
-
McGonagle D, Sharif K, O’Regan A, Bridgewood C (2020) The role of cytokines including interleukin-6 in COVID-19 induced pneumonia and macrophage activation syndrome-like disease. Autoimmunity Rev 19(6): 102537.
-
Qin C, Zhou L, Hu Z, Zhang S, Yang S, et al. (2020) Dysregulation of immune response in patients with coronavirus 2019 (COVID-19) in Wuhan, China. Clinical infectious diseases 71(15): 762-768.
-
Queval CJ, Brosch R, Simeone R (2017) The macrophage: a disputed fortress in the battle against Mycobacterium tuberculosis. Frontiers in microbiology 8: 2284.
-
Lerner TR, Borel S, Gutierrez MG (2015) The innate immune response in human tuberculosis. Cellular microbiology 17(9): 1277-1285.
-
Gou X, Pan L, Tang F, Gao H, Xiao D (2018) The association between Vitamin D status and tuberculosis in children: a meta-analysis. Medicine 97(35).
-
Yuk JM, Shin DM, Lee HM, Yang CS, Jin HS, et al. (2009) Vitamin D3 induces autophagy in human monocytes/ macrophages via cathelicidin. Cell Host Microbe 6(3): 231-243.
-
Groeschel C, Aggarwal A, Tennakoon S, Hoebaus J, Prinz Wohlgenannt M, et al. (2016) Effect of 1, 25-dihydroxyvitamin D3 on the Wnt pathway in non-malignant colonic cells. The Journal of steroid biochemistry and molecular biology 155: 224-230.
-
Bustamante J, Arias AA, Vogt G, Picard C, Galicia LB, et al. (2011) Germline CYBB mutations that selectively affect macrophages in kindreds with X-linked predisposition to tuberculous mycobacterial disease. Nat Immunol 12(3): 213-221.
-
Sia JK, Georgieva M, Rengarajan J (2015) Innate immune defenses in human tuberculosis: an overview of the interactions between Mycobacterium tuberculosis and innate immune cells. Journal of immunology research 2015: 747543.
-
Sun L, Wu J, Du F, Chen X, Chen ZJ (2013) Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 339(6121): 786-791.
-
Neyrolles O, Wolschendorf F, Mitra A, Niederweis M (2015) Mycobacteria, metals, and the macrophage. Immunol Rev 264(1): 249-263.
-
Botella H, Peyron P, Levillain F, Poincloux R, Poquet Y, et al. (2011) Mycobacterial P1-type ATPases mediate resistance to zinc poisoning in human macrophages. Cell host & microbe 10(3): 248-259.
-
Smyth MJ, Cretney E, Kelly JM, Westwood JA, Street SEA, et al. (2005) Activation of NK cell cytotoxicity. Molecular immunology 42(4): 501-510.
-
van Erp EA, Luytjes W, Ferwerda G, van Kasteren PB (2019) Fc-mediated antibody effector functions during respiratory syncytial virus infection and disease. Front Immunol 10: 548.
-
Yasui F, Kohara M, Kitabatake M, Nishiwaki T, Fujii H, et al. (2014) Phagocytic cells contribute to the antibody- mediated elimination of pulmonary-infected SARS coronavirus. Virology 454-455: 157-68.
-
Kamran K (2020) COVID-19: an Immunopathological View. mSphere 5(2).
-
Weiss RC, Scott FW (1981) Antibody-mediated enhancement of disease in feline infectious peritonitis: comparisons with dengue hemorrhagic fever. Comp Immunol Microbiol Infect Dis 4(2): 175-189.
-
Yip MS, Leung NH, Cheung CY, Li PH, Lee HH, et al. (2014) Antibody-dependent infection of human macrophages by severe acute respiratory syndrome coronavirus. Virol J 11: 82.
-
Hohdatsu T, Yamada M, Tominaga R, Makino K, Kida K, et al. (1998) Antibody-dependent enhancement of feline infectious peritonitis virus infection in feline alveolar macrophages and human monocyte cell line U937 by serum of cats experimentally or naturally infected with feline coronavirus. Journal of veterinary medical science 60(1): 49-55.
-
Mogensen SC (1979) Role of macrophages in natural resistance to virus infections. Microbiol Rev 43(1): 1-26.
-
Mogensen SC (1988) Macrophages and natural resistance to virus infections. In Infection. Springer, pp: 207-223.
-
Gordon S, Martinez FO (2010) Alternative activation of macrophages: mechanism and functions. Immunity 32(5): 593-604.
-
Ley K (2017) M1 means kill; M2 means heal. J Immunol 199(7): 2191-2193.
-
Murray PJ, Allen JE, Biswas SK, Fisher EA, Gilroy DW, et al. (2014) Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity 41(1): 14-20.
-
Orecchioni M, Ghosheh Y, Pramod AB, Ley K (2019) Macrophage polarization: different gene signatures in M1 (LPS+) vs. classically and M2 (LPS-) vs. alternatively activated macrophages. Frontiers in immunology.
-
Costantini A, Viola N, Berretta A, Galeazzi R, Matacchione G, et al. (2018) Age-related M1/M2 phenotype changes in circulating monocytes from healthy/unhealthy individuals. Aging (Albany NY) 10(6): 1268-1280.
-
Linehan E, Fitzgerald D (2015) Ageing and the immune system: focus on macrophages. Eur J Microbiol Immunol 5(1): 14-24.
-
Eapen MS, Hansbro PM, McAlinden K, Kim RY, Ward C, et al. (2017) Abnormal M1/M2 macrophage phenotype profiles in the small airway wall and lumen in smokers and chronic obstructive pulmonary disease (COPD). Sci Rep 7(1): 13392.
-
Arora S, Dev K, Agarwal B, Das P, Syed MA (2018) Macrophages: their role, activation and polarization in pulmonary diseases. Immunobiology 223(4-5): 383-
-
Heusinkveld M, van Steenwijk PJdV, Goedemans R, Ramwadhdoebe TH, Gorter A, et al. (2011) M2 macrophages induced by prostaglandin E2 and IL-6 from cervical carcinoma are switched to activated M1 macrophages by CD4+ Th1 cells. The Journal of Immunology 187(3): 1157-1165.
-
Yilla M, Harcourt BH, Hickman CJ, McGrew M, Tamin A, et al. (2005) SARS-coronavirus replication in human peripheral monocytes/macrophages. Virus Res 107(1): 93-101.
-
Fu Y, Cheng Y, Wu Y (2020) Understanding SARS-CoV-2- mediated inflammatory responses: from mechanisms to potential therapeutic tools. Virol Sin 35(3): 266-271.
-
Liu L, Wei Q, Lin Q, Fang J, Wang H, et al. (2019) Anti– spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection. JCI Insight 4(4): e123158.
-
Bolles M, Deming D, Long K, Agnihothram S, Whitmore A, et al. (2011) A double-inactivated severe acute respiratory syndrome coronavirus vaccine provides incomplete protection in mice and induces increased eosinophilic proinflammatory pulmonary response upon challenge. J Virol 85(23): 12201-12215.
-
Tseng CT, Sbrana E, Iwata-Yoshikawa N, Newman PC, Garron T, et al. (2012) Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PloS one 7(4): e35421.
-
Flynn JL, Chan J (2001) Immunology of tuberculosis. Annual review of immunology 19(1): 93-129.
-
Giacomini E, Iona E, Ferroni L, Miettinen M, Fattorini L, et al. (2001) Infection of human macrophages and dendritic cells with Mycobacterium tuberculosis induces a differential cytokine gene expression that modulates T cell response. J Immunol 166(12): 7033-7041.
-
Khan N, Vidyarthi A, Nadeem S, Negi S, Nair G, et al. (2016) Alteration in the gut microbiota provokes susceptibility to tuberculosis. Frontiers in immunology 7: 529.
-
Mihret A (2012) The role of dendritic cells in Mycobacterium tuberculosis infection. Virulence 3(7): 654-659.
-
Wu CY, Hu HY, Pu CY, Huang N, Shen HC, et al. (2011) Pulmonary tuberculosis increases the risk of lung cancer: a population‐based cohort study. Cancer 117(3): 618-624.
-
Balboa L, Romero MM, Yokobori N, Schierloh P, Geffner L, et al. (2010) Mycobacterium tuberculosis impairs dendritic cell response by altering CD1b, DC‐SIGN and MR profile. Immunol Cell Biol 88(7): 716-726.
-
Georgieva M, Sia JK, Bizzell E, Lala RM, Rengarajan J (2018) Mycobacterium tuberculosis GroEL2 modulates dendritic cell responses. Infect Immun 86(2): e00387- 00417.
-
Velasquez LN, Stüve P, Gentilini MV, Swallow M, Bartel J, et al. (2018) Targeting Mycobacterium tuberculosis antigens to dendritic cells via the DC-specific-ICAM3- grabbing-nonintegrin receptor induces strong T-helper 1 immune responses. Front Immunol 9: 471.
-
Ehlers S (2010) DC-SIGN and mannosylated surface structures of Mycobacterium tuberculosis: a deceptive liaison. Eur J Cell Biol 89(1): 95-101.
-
Kaufmann SHE, Schaible UE (2003) A dangerous liaison between two major killers: Mycobacterium tuberculosis and HIV target dendritic cells through DC-SIGN. The Journal of experimental medicine 197(1): 1-5.
-
Guo N, Marra CA, Marra F, Moadebi S, Elwood RK, et al. (2008) Health state utilities in latent and active tuberculosis. Value in health 11(7): 1154-1161.
-
Friedman A, Turner J, Szomolay B (2008) A model on the influence of age on immunity to infection with Mycobacterium tuberculosis. Experimental Gerontology 43(4): 275-285.
-
Chen J, Subbarao K (2007) The immunobiology of SARS. Annu Rev Immunol 25: 443-472.
-
Ng N, Padmawati RS, Prabandari YS, Nichter M (2008) Smoking behavior among former tuberculosis patients in Indonesia: intervention is needed. Int J Tuberc Lung Dis 12(5): 567-572.
-
Lin HH, Murray M, Cohen T, Colijn C, Ezzati M (2008) Effects of smoking and solid-fuel use on COPD, lung cancer, and tuberculosis in China: a time-based, multiple risk factor, modelling study. The lancet 372(9648): 1473-1483.
-
Yoshikawa TT, Marrie TJ (2000) Community-acquired pneumonia in the elderly. Clinical infectious diseases 31(4): 1066-1078.
-
Ventura SPM, E Silva FA, Quental MV, Mondal D, Freire MG, et al. (2017) Ionic-liquid-mediated extraction and separation processes for bioactive compounds: past, present, and future trends. Chem Rev 117(10): 6984- 7052.
-
Agrawal A, Agrawal S, Gupta S (2017) Role of dendritic cells in inflammation and loss of tolerance in the elderly. Front Immunol 8: 896.
-
Wong JP, Viswanathan S, Wang M, Sun LQ, Clark GC, et al. (2017) Current and future developments in the treatment of virus-induced hypercytokinemia. Future Med Chem 9(2): 169-178.
-
Wong SL, Ji H, Hollenbeck BK, Morris AM, Baser O, et al. (2007) Hospital lymph node examination rates and survival after resection for colon cancer. JAMA 298(18): 2149-2154.
-
Kumar V (2018) Dendritic cells in sepsis: potential immunoregulatory cells with therapeutic potential. Molecular immunology 101: 615-626.
-
Lin Q, Zhao S, Gao D, Lou Y, Yang S, et al. (2020) A conceptual model for the coronavirus disease 2019 (COVID-19) outbreak in Wuhan, China with individual reaction and governmental action. International journal of infectious diseases 93: 211-216.
-
Pillaiyar T, Meenakshisundaram S, Manickam M (2020) Recent discovery and development of inhibitors targeting coronaviruses. Drug discovery today 25(4): 668-688.
-
Ueno K, Kinjo Y, Okubo Y, Aki K, Urai M, et al. (2015) Dendritic cell-based immunization ameliorates pulmonary infection with highly virulent Cryptococcus gattii. Infection and immunity 83(4): 1577-1586.
-
Zhou F, Yu T, Du R, Fan G, Liu Y, et al. (2020) Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet 395(10229): 1054-1062.
-
Vatansever D, Wang S, Sahakian BJ (2021) Covid-19 and promising solutions to combat symptoms of stress, anxiety and depression. Neuropsychopharmacology 46(1): 217-218.
-
Schindler R, Mancilla J, Endres S, Ghorbani R, Clark SC, et al. (1990) Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 75(1): 40-47.
-
Shiratsuchi H, Johnson JL, Ellner JJ (1991) Bidirectional effects of cytokines on the growth of Mycobacterium avium within human monocytes. The Journal of Immunology 146(9): 3165-3170.
-
Ladel CH, Blum C, Dreher A, Reifenberg K, Kopf M, et al. (1997) Lethal tuberculosis in interleukin-6-deficient mutant mice. Infection and immunity, 65(11): 4843- 4849.
-
Saunders BM, Frank AA, Orme IM, Cooper AM (2000) Interleukin-6 induces early gamma interferon production in the infected lung but is not required for generation of specific immunity to Mycobacterium tuberculosis infection. Infection and immunity 68(6): 3322-3326.
-
Leal IS, Smedegard B, Andersen P, Appelberg R (1999) Interleukin-6 and interleukin-12 participate in induction of a type 1 protective T-cell response during vaccination with a tuberculosis subunit vaccine. Infection and immunity 67(11): 5747.
-
Farsalinos K, Barbouni A, Niaura R (2020) Systematic review of the prevalence of current smoking among hospitalized COVID-19 patients in China: could nicotine be a therapeutic option? Intern Emerg Med 15(5): 845- 852.
-
Liu Q, Zhou YH, Yang ZQ (2016) The cytokine storm of severe influenza and development of immunomodulatory therapy. Cell Mol Immunol 13(1): 3-10.
-
Tisoncik JR, Korth MJ, Simmons CP, Farrar J, Martin TR, et al. (2012) Into the eye of the cytokine storm. Microbiology and Molecular Biology Reviews 76(1): 16- 32.
-
Shimabukuro Vornhagen A, Godel P, Subklewe M, Stemmler HJ, Schlosser HA, et al. (2018) Cytokine release syndrome. Journal for immunotherapy of cancer 6(1): 56.
-
İnandıklıoğlu N, Akkoc T (2020) Immune responses to SARS-CoV, MERS-CoV and SARS-CoV-2. Cell Biology and Translational Medicine 1288: 5-12.
-
Wiersinga WJ, Rhodes A, Cheng AC, Peacock SJ, Prescott HC, et al. (2020) Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID-19): A Review. JAMA 324(8): 782-793.
-
Lerkvaleekul B, Vilaiyuk S (2018) Macrophage activation syndrome: early diagnosis is key. Open Access Rheumatol 10: 117-128.
-
Ravelli A (2002) Macrophage activation syndrome. Current opinion in rheumatology 14(5): 548-552.
-
Tay MZ, Poh CM, Rénia L, MacAry PA, Ng LFP (2020) The trinity of COVID-19: immunity, inflammation and intervention. Nature reviews immunology 20(6): 363- 374.
-
Chen G, Wu DI, Guo W, Cao Y, Huang D, et al. (2020) Clinical and immunological features of severe and moderate coronavirus disease 2019. J Clin Invest 130(5): 2620-2629.
-
Kim JY, Ko JH, Kim Y, Kim YJ, Kim JM, et al. (2020) Viral load kinetics of SARS-CoV-2 infection in first two patients in Korea. Journal of Korean medical science 35(7).
-
Liu B, Li M, Zhou Z, Guan X, Xiang Y (2020) Can we use interleukin-6 (IL-6) blockade for coronavirus disease 2019 (COVID-19)-induced cytokine release syndrome (CRS)?. J Autoimmun 111: 102452.
-
Pan Y, Zhang D, Yang P, Poon LLM, Wang Q (2020) Viral load of SARS-CoV-2 in clinical samples. The Lancet infectious diseases 20(4): 411-412.
-
Phan LT, Nguyen TV, Luong QC, Nguyen TV, Nguyen HT, et al. (2020) Importation and human-to-human transmission of a novel coronavirus in Vietnam. New England journal of medicine 382(9): 872-874.
-
Liu F, Li L, Xu M, Wu J, Luo D, et al. (2020) Prognostic value of interleukin-6, C-reactive protein, and procalcitonin in patients with COVID-19. J Clin virol 127: 104370.
-
Hirano T, Murakami M (2020) COVID-19: a new virus, but a familiar receptor and cytokine release syndrome. Immunity 52(5): 731-733.
-
Wong CK, Lam CW, Wu AK, Ip WK, Lee NL, et al. (2004) Plasma inflammatory cytokines and chemokines in severe acute respiratory syndrome. Clin Exp Immunol 136(1): 95-103.
-
Huang X, Wei F, Hu L, Wen L, Chen K (2020) Epidemiology and clinical characteristics of COVID-19. Archives of Iranian medicine 23(4): 268-271.
-
Wan S, Yi Q, Fan S, Lv J, Zhang X, et al. (2020) Characteristics of lymphocyte subsets and cytokines in peripheral blood of 123 hospitalized patients with 2019 novel coronavirus pneumonia (NCP).
-
Okabayashi T, Kariwa H, Yokota SI, Iki S, Indoh T, et al. (2006) Cytokine regulation in SARS coronavirus infection compared to other respiratory virus infections. J Med Virol 78(4): 417-424.
-
Diao B, Wang C, Tan Y, Chen X, Liu Y, et al. (2020) Reduction and functional exhaustion of T cells in patients with coronavirus disease 2019 (COVID-19). Front Immunol 11: 827.
-
Han H, Ma Q, Li C, Liu R, Zhao L, et al. (2020) Profiling serum cytokines in COVID-19 patients reveals IL-6 and IL-10 are disease severity predictors. Emerging microbes & infections 9(1): 1123-1130.
-
Guan WJ, Ni ZY, Hu Y, Liang WH, Ou CQ, et al. (2020) Clinical characteristics of coronavirus disease 2019 in China. New England journal of medicine 382(18): 1708- 1720.
-
Liu Y, Yang Y, Zhang C, Huang F, Wang F, et al. (2020) Clinical and biochemical indexes from 2019-nCoV infected patients linked to viral loads and lung injury. Sci China Life Sci 63(3): 364-374.
-
Channappanavar R, Perlman S (2017) Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. Semin Immunopathol 39(5): 529-539.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions