Low Density Lipoproteinemia in Obese and Type-2 Diabetics from Association of IR and LDLR
Universality of association of two receptors, LDLR (LDL receptor) and IR (insulin receptor), has been explored in various cell types and intracellular compartments including caveolae vesicles in plasma membrane. The existence of association of LDLR and IR has been proved in type-2 diabetes and diabetes associated obesity. The non-functionality of LDLR in conjugation with IR has been found as of a prime reason for developing LDL related atherosclerosis in insulin defunct system. Confocal microcopy and western blots are the major techniques used to explore this study.
Gopalakrishnan Ramakrishnan1,3, Puja Beriwal1,4, Sangam2 and N.
C. Chandra2*
Present Address
Phulwarishariff, Patna, India, Email: nc1_chandra@hotmail.com
system. Confocal microcopy and western blots are the major techniques used to explore this study.
Keywords: Insulin Receptor; LDL Receptor; Diabetes; Lipoproteinemia; Atherosclerosis
Introduction
Previous reports from our laboratory have shown a direct correlation between diabetes and atherosclerosis by demonstrating non-functionality of LDL receptor (LDLR) in clearing extracellular LDL when remains in complex association with insulin receptor (IR) [1]. Insulin can dissociate this association and make LDLR free with its full functional activity [2]. We have shown a strong association of these two receptors in type-1 diabetes having inadequate level of insulin [1, 3]. Although we have shown by electron microscopy that this association is present at intracellular space [2], it is not clear from these studies how universally and specifically this association exists in intracellular locations. It is also an enigma whether this association is also existed in type-2 diabetic subjects. Type-2 diabetes is also inter-related with obesity and obesity precipitates atherosclerosis. So speculation of association of LDLR and IR cannot be avoided also to project atherosclerotic excerpts in obese subjects. Confocal microscopy and immunoprecipitates in western blot have been exploited in present study to explore intracellular locations of this association and plausibility of such association in type-2 diabetes and obesity where hyperinsulinemia exists owing to insulin resistance.
Materials
Kit(s) for insulin estimation was purchased from Mercodia Ab, Sylveniusgatan 8a, Se-754 50 Uppsala, Sweden; glucose estimation kits were from Dia Sys Diagnostic Systems Gmbh, Holzheim, Germany; kits for LDL estimation was from Giess Diagnostics snc, Via Crevinara, Rome, Italy. Regular and fluorescein (red PE and green FITC) lebeled antibodies were purchased from Santa Cruz Biotechnology, Inc. California, USA (DAKOLSAB + Kit). Ethylene diamine tetra acetate (EDTA), Antibiotic/Antifungal solu-tion (100×), DMEM and RPMI- 1640 powder were purchased from Sigma Chemical Co., St. Louis, MO, USA. Fetal Bovine Serum (FBS) was obtained from GIBCO BRL, Life Technologies, Inc. Grand Island, NY, and USA. DAB Substrate Kit for Peroxidase and Strep-tavidin Peroxidase Kit were bought from Vector, Laboratories, Inc., Burlingame, CA, U.S.A. Comassie brilliant blue R-250 was purchased from Bio-Rad laboratories, Hercules, USA. All other chemicals used were of analytical re-agent grade. Cell lines (HepG2 and THP-1) were purchased from NCCS, Ganeshkind, Pune, India. Goat liver tissue was obtained from local slaughter shop.
Methods
• Cell culture: HepG2 cells were grown in high glucose (4.5 g/L) DMEM and THP-1 cells were grown in RPMI- 1640 supplemented with 10% FBS and 100 U/mL penicillin, 100µg/mL streptomycin and 0.25µg/mL amphotericin-B.
• Tissue sample preparation: Fresh goat liver was procured from the local slaughter shop within 30 minutes of slaughter and carried in ice cold saline during transportation. The tissue was then washed thoroughly in ice cold saline to remove the associated blood. For confocal microscopy small 1 cm3 cubes were dissected out from lobular side of the liver. The cubes were immediately snap frozen in liquid nitrogen and then used for block preparation using the cryogel at - 25°C. Once the blocks were made, 5µm-10µm thick sections were cut and lifted on glass slides. The samples were then stored at -20°C and used for experiment within a week time.
• Preparation of cultured cell for confocal microscopy: Cells were seeded onto 12mm cover glasses in a 6 well plate @ 5 x 105 cells/ well and grown same way as described under cell culture in this section. 50mM PMA was used in the medium of THP-1 cells and incubated for 24h at 37°C to induce adherence.
• Confocal microscopy: The protocol for confocal microscopy was similar as reported earlier [1].
• Western blot: This was used to compare receptor protein expressions. The protocol of the procedure to precipitate respective receptors has been depicted in earlier reports from our laboratory [1, 2]. The estimation of other parameters from blood plasma were mainly Kit based following the protocol as directed in manufacturer’s manual. Results
Complex Association of IR and LDLR
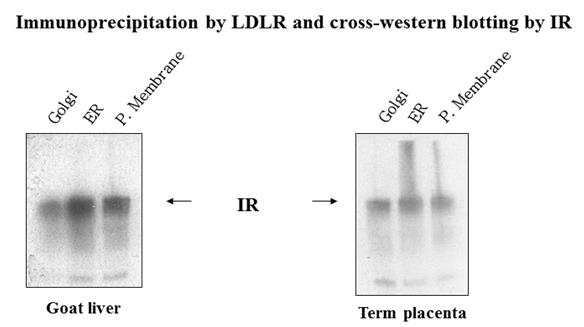
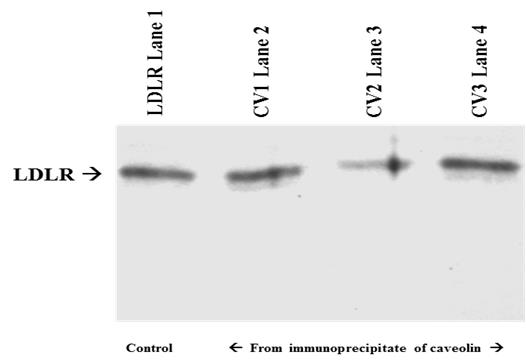
Confocal microscopy: The topographical location of two receptors, IR (green labelled) and LDLR (red labelled) were studied by confocal microscopy. Green FITC and red PE fluorescent dyes were used to identify IR and LDLR. Zooming the receptors through confocal microscope, the merged yellow colour reveals an overlap of the two receptors in the similar zone of plasma membrane. In other words, the merging of these two receptors satisfies the earlier reports of electron microscopy and immune precipitation over the association and complex adherence of IR and LDLR on plasma membrane surface [2, 3]. The universality of this association in various cell type viz. cell culture and tissue cells has been shown in Figure 1. Western blots: Universality of association of the two receptors in cell organelles e.g. in endoplasmic reticulum (ER), golgi apparatus and plasma membrane has been explored by immuno-precipitating LDLR from different tissue cells viz. goat liver and term placenta, and by performing western blot of IR from the immune- precipitate of LDLR (Figure 2). The presence of IR band in the immune-precipitate of LDLR marks a strong evidence of association and adherence of these two receptor proteins in the post translational steps through endoplasmic reticulum and golgi body to plasma membrane. Proof of evidence of association of the receptors within the caveolae of plasma membrane has been revealed by observing LDLR band in the western blot from the immune-precipitate of IR which has been immune-precipitated from caveolae protein (caveolin). The caveolin protein was immune-precipitated by anti- caveolin antibodies (CV1, CV2 and CV3) from plasma membrane fraction isolated from HepG2 cells (Figure 3). It is noteworthy to mention that insulin receptor was originally found within the caveolae of the surface membrane [4, 5].
![Figure 1: **Western blots:** Universality of association of the two receptors in cell organelles e.g. in endoplasmic reticulum (ER), golgi apparatus and plasma membrane has been explored by immuno-precipitating LDLR from different tissue cells viz. goat liver and term placenta, and by performing western blot of IR from the immune- precipitate of LDLR (Figure 2). The presence of IR band in the immune-precipitate of LDLR marks a strong evidence of association and adherence of these two receptor proteins in the post translational steps through endoplasmic reticulum and golgi body to plasma membrane. Proof of evidence of association of the receptors within the caveolae of plasma membrane has been revealed by observing LDLR band in the western blot from the immune-precipitate of IR which has been immune-precipitated from caveolae protein (caveolin). The caveolin protein was immune-precipitated by anti- caveolin antibodies (CV1, CV2 and CV3) from plasma membrane fraction isolated from HepG2 cells (Figure 3). It is noteworthy to mention that insulin receptor was originally found within the caveolae of the surface membrane [4,5].](/fulltextimages/3406/fig_1.jpeg)
Obesity
Plasma component: The relative plasma levels of insulin, glucose and LDL in normal and obese subjects are shown in table-1. It is very much transparent from table-1 that as obesity advances, the propensity of induction of hyperinsulinemia (insulin resistance), hyperglycemia (preferable type-2 diabetes) and LDL related lipoproteinemia (pro-atherosclerosis) also increases. Previous reports [1, 2] from this laboratory has shown the existence of complex association of IR and LDLR in type-1 diabetic subjects and associated hyperlipoproteinemia due to increased plasma LDL level which has been the outcome of inactivity of LDLR trapped in the IR-LDLR complex. So increase of LDL concentration laterally with increase of glucose level in the advancement of diabetes prone obesity might be a reason of complex association of IR and LDLR as depicted in previous reports. In fact it is shown in Figure 4 that insulin receptor tyrosine phosphorylation occurs at low level in obese subjects as compared to normal ones. Insulin receptor tyrosine phosphorylation is a property of activation and functionally active insulin receptor [6, 7]. Receptor tyrosine phosphorylation: Insulin receptor gets activated by phosphorylation of its receptor tyrosine residue to send extracellular signals to intracellular signalling components for gene activation and expression. Inactivation of IR receptor by its adherence with LDLR may be expected to reduce the level of receptor tyrosine phosphorylation. In fact this may happen in obesity as shown in Figure 4. The western blot with anti-tyrosine- phosphate antibody shows a decrease in the band density of IR-tyrosine-phosphate in the peripheral blood mononuclear cells (PBMC) of obese subject. Similar results were also found in other blood borne cells too (not shown) with the progress of obesity. This could be the reason of lateral inactivation of IR and LDLR in IR-LDLR complex.
Type2 diabetes
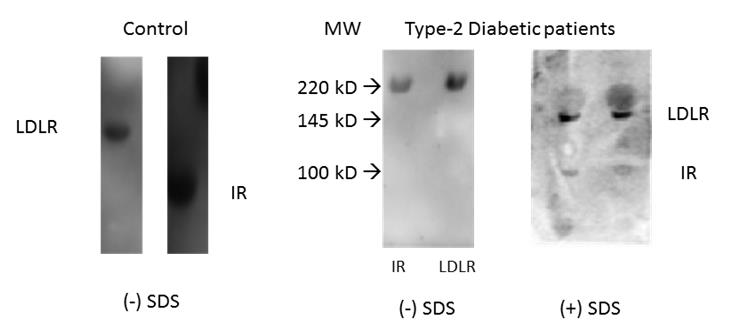
Previous report of the association of IR and LDLR in diabetic subjects has been shown only with type-1 diabetic subjects where insulin level is either very low or absent. There is so far no report of such association of IR and LDLR in type-2 diabetics where hyper insulinemia exists. Figure 5 shows the status of IR and LDLR in samples from type-2 diabetic subjects as compared to their status in control ones. In type-2 diabetic subjects both IR and LDLR bands were found at the same site of higher mol. wt. region as a single band in lanes showing without SDS. With SDS the single band breaks into two bands (LDLR and IR) and move at respective mol. wt. region. The positions of LDLR and IR as per their mol. wt. in normal subjects are shown in control lane in absence of SDS. This shows that the association of IR and LDLR also exists in type-2 diabetic subjects even in presence of hyperinsulinemia.
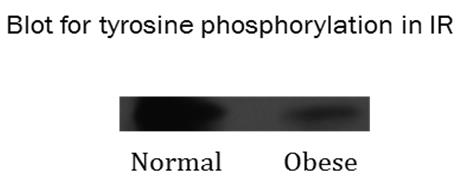
Figure 4: The picture shows western blot of insulin receptor tyrosine phosphate level in peripheral blood mononuclear cells (PBMC) of normal and obese subjects. Anti-tyrosine-phosphate antibody was used to precipitate the receptor protein tagged with tyrosine phosphate from IR immune precipitate. The band density of receptor tyrosine phosphate is found to be decreased in obese subjects as compared to normal subjects.
Discussion
One of the major reasons of atherosclerosis is deposition of LDL particles in blood vessels due to impaired clearance of LDL from blood plasma [8, 9]. The clearance of LDL particles by LDL receptor located on the surface membrane of cells around the vessel wall was a landmark discovery of Goldstein and Brown [10]. The prime reason of accumulation of LDL particles in blood vessels is therefore considered from the result of defective functioning of LDL receptor (LDLR) [11, 12]. Previous reports from our laboratory have shown an association and adherence of the two receptors (IR and LDLR) as a prime reason for their inactivity in biological cells [1, 2, 3]. Besides intracellular organelles, the association is also noticed on surface membrane and within caveole of the surface membrane (Figures 2 and 3). Functionally inactive LDL receptors (LDLRs) cannot clear LDL particles from blood vessels and gradually develop atheroma. It is also shown that insulin could separate the two receptors and make them functionally active [2]. Like type-1 diabetes, our present study shows the presence of the same adherence in type-2 diabetes too. In type-2 diabetes hyperinsulinemia exists due to insulin resistance. Presence of hyperinsulinemia even cannot dissociate the receptor complex into free IR and LDLR. This proves that the separation of the two receptors is not because of mere attachment of ligand insulin to its receptor. Some insulin mediated signalling event may be responsible for dissociation of the receptors. Insulin generated signals are transmitted via a receptor tyrosine phosphorylation [6, 7]. Many type-2 diabetics are obese also [13, 14] (Table 1).
| Normal subjects | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Obese (n=15) Mean ± | Morbid Obese (n=15 | ) | ||||||||||||
| Variable | Normal range | (n=15) | ||||||||||||
| SD | Mean ± SD | |||||||||||||
| Mean ± SD | ||||||||||||||
| Insulin | 2 – 25mU/L | 15.06 ± 12.3 | 21.04 ± 14.6 | 26.8 ± 12.6 | ||||||||||
| Glucose | 70 – 110mg/dL | 86.55 ± 14.7 | 112.45 ± 19.8 | 127.74 ± 7.14 | ||||||||||
| LDL | < 120 mg/dL | 100.09 ± 7.1 | 111.29 ± 2.27 | 136 ± 8.7 |
Table 1: Comparison of fasting plasma parameters in normal and obese subjects.
Acknowledgement
The authors are thankful to Department of Biotechnology, Govt. of India and Indian Council of
References
-
Suneja S, Ramakrishnan G, Tandon N, Chandra NC (2011) Modulation by insulin of the co-localized LDL receptor in normal and Type-I Diabetic subjects. International Journal of Clinical Medicine 2(3): 231- 245.
-
Ramakrishnan G, Arjuman A, Suneja S, Das C, Chandra NC (2012) The association between insulin and low- density lipoprotein receptors. Diabetes & Vascular Disease Research 9(3): 196-204.
-
Suneja S, Christian Y, Chandra NC (2018) Milieu of Diabetes in the 2nd Decade of 21st Century. J Diabetes Metab 9(9): 804.
-
Gustavsson J, Parpal S, Karlsson M, Ramsing C, Thorn H, et al. (1999) Localization of the insulin receptor in caveolae of adipocyte plasma membrane. FASEB J 13(14): 1961-1971.
-
Cohen AW, Combs TP, Scherer PE, Lisanti MP (2003) Role of caveolin and caveolae in insulin signaling and diabetes. Am J Physiol Endocrinol Metab 285(6): E1151-E1160.
-
Ullrich A, Bell JR, Chen EY, Herrera R, Petruzelli LM, et al. (1985) Human insulin receptor and its relationship to the tyrosine kinase family of oncogenes. Nature 313: 765-761.
-
Petruzzelli LM, Ganguly S, Smith CJ, Cobb MH, Rubin CS, et al. (1982) Insulin activates a tyrosine-specific protein kinase in extracts of 3T3-L1 adipocytes and human placenta. Proc Natl Acad Sci USA 79(22): 6792-6796.
-
Borén, J, Williams KJ (2016) The central role of arterial retention of cholesterol-rich apolipoprotein- B-containing lipoproteins in the pathogenesis of atherosclerosis: a triumph of simplicity. Current Opinion in Lipidology 27(5): 473-483.
-
Linton MRF, Yancey PG, Davies SS, Jerome WG, Linton EF, et al. (2015) The Role of Lipids and Lipoproteins in Atherosclerosis. Endotext.
-
Goldstein JL, Brown MS (1986) A receptor mediated pathway for cholesterol homeostasis. Science 232(4746): 34-47.
-
Goldstein JL, Brown MS (1977) The low density lipoprotein pathway and its relation to atherosclerosis. Ann Rev Biochem 46: 897-930.
-
Ross (1995) Cell biology of atherosclerosis. Ann Rev Physiol 57: 791-804.
-
Al-Goblan AS, Al-Alfi MA, Khan MZ (2014) Mechanism linking diabetes mellitus and obesity. Diabetes Metab Syndr Obes 7: 587-591.
-
Eckel RH, Kahn SE, Ferrannini E, Goldfine AB, Nathan DM, et al. (2011) Obesity and Type 2 Diabetes: What Can Be Unified and What Needs to Be Individualized? J Clin Endocrinol Metab 96(6): 1654-1663.
- Investigation of Polymorphisms in PPAR-Ɣ and TRHR Genes and their Impact on Turkish Diabetic and Obese Individuals
- The Impact of Aircraft Noise Exposure on the Efficacy of Empagliflozin Therapy in an Animal Model of Obesity
- Rooibos Mitigates Metabolic and Inflammatory Dysfunctions in Mice Fed a High-Carbohydrate Diet
- Synergistic Effect of Combined Leaf Extract of Vernonia amygdalina, Ocimum gratissimum, and Zingiber officinale Tuber on Phytochemical Profile, Antioxidant Activity, Serum Insulin, and Biochemical Parameters in Streptozotocin-Induced Diabetic Rats
- Investigation of Cardiovascular Responses to Aerobic Exercise in Obese University Students
- A Look at the Phase Angle Obtained by Electrical Bioimpedance