Improvement of Glucose Intolerance in Asian-Type Diabetes Model Mice Using Pentadecyl, Pentadecanoic Acid-Based Triglyceride
Even-chain saturated fatty acid concentrations are associated with an increased risk of coronary heart disease, whereas odd- chain saturated fatty acid concentrations, such as pentadecanoic acid (15:0) and heptadecanoic acid (17:0), are associated with a decreased risk. Similarly, even-chain saturated fatty acids are positively associated with type 2 diabetes (T2D), whereas odd-chain saturated fatty acids are thought to be inversely associated with T2D. We purified pentadecyl, pentadecanoic acid- based triglycerides from Aurantiochytrium limacinum, a type of marine microalgae, and demonstrated the effect of pentadecyl on insulin secretion both in vitro and in vivo. Pentadecyl increased the cell viability of immortalized mouse pancreatic beta cells in a dose-dependent manner and induced high glucose-stimulated insulin secretion in the cells. Pancreatic beta cell- specific CDKAL1-deficient mice, a model of Asian-type T2D, were given pentadecyl with a low- or high-fat diet for 9 weeks, and glucose tolerance tests were performed. Pentadecyl improved the glucose intolerance of the mice and decreased the expressions of ER stress-related genes in the islets of the mice. These results suggest that pentadecyl can improve insulin secretion through the reduction of ER stress in patients with Asian-type T2D.
Introduction
Saturated fatty acids (SFAs) are thought to be associated with many chronic conditions, including cardiovascular disease, cancers, and systemic inflammation [1], and are capable of stimulating an inflammatory response through the TLR4 signaling pathway [1, 2, 3]. SFAs are divided into two classes, even-chain saturated fatty acids (ECFAs), such as lauric acid (C12:0) and palmitic acid (C16:0), and odd- chain saturated fatty acids (OCFAs), such as pentadecanoic acid (C15:0) and heptadecanoic acid (C17:0). Furthermore, the functions of ECFAs and OCFAs are thought to be mutually exclusive. Dietary ECFAs have been associated with an increased risk of inflammation, heart disease, and type 2 diabetes (T2D) in humans [3, 4, 5, 6], whereas higher dietary intake and circulating levels of OCFAs have been associated with lower risks of chronic inflammation, cardiovascular disease, metabolic syndrome, and T2D [6, 7, 8, 9]. Oral administration of pentadecanoic acid (C15:0) from dairy reduces circulating concentrations of pro-inflammatory cytokines, glucose, and cholesterol in obese C57BL/6J mice fed a high-fat diet (HFD) [6]. These results suggest that dietary intake of OCFAs, such as pentadecanoic acids (C15:0), may be useful for the prevention and improvement of glucose intolerance. However, approximately 68% of the fatty acids present in whole-fat milk are ECFAs, whereas OCFAs, namely pentadecanoic acid (C15:0) and heptadecanoic acid (C17:0), represent only 1% and 0.5% of the fatty acids in whole-fat milk, respectively [6, 10]. In general, it is difficult to take in only OCFAs from natural foods.
Aurantiochytrium is an oleaginous microorganism in the Thraustochytriaceae family and contains high contents of squalene, docosahexaenoic acid, and pentadecanoic acid (C15:0) [11, 12]. The major lipids of Aurantiochytrium comprise squalene, carotenoids, triglycerides, glycolipids, and phospholipids [12]. We extracted the triglycerides using a known fat and oil extraction method to obtain triglycerides from Aurantiochytrium. The triglyceride with the highest melting point was purified by repeated recrystallization and named pentadecyl. Pentadecyl was composed mainly of pentadecanoic acid (C15:0), tridecanoic acid (C13:0), and heptadecanoic acid (C17:0).
Cdk5 regulatory subunit associated protein 1-like 1 (CDKAL1) is one of the most reliable risk genes for T2D among all ethnic groups [13]. The ratio of the risk allele of CDKAL1 in Asians is higher than that in European populations [14]. CDKAL1 is the best-replicated susceptibility locus and the most associated with T2D among several susceptibility loci in the Japanese population [15]. The risk variants of CDKAL1 are associated with reduced insulin secretion but not obesity [16]. Asian-type T2D is characterized by a decrease in insulin secretion and less insulin resistance or obesity [17]. Thus, the variants of CDKAL1 may be implicated in T2D in Asian populations.
CDKAL1 is a mammalian methylthiotransferase that catalyzes the 2-methylthio (ms2) modification of N6-threonyl-carbamoyladenosine (t6A) to produce 2-methylthio-N6-threonyl-carbamoyladenosine (ms2t6A) at position 37 of tRNALys (UUU) [18, 19]. The ms2-modification of tRNALys(UUU) is critical for accurate decoding of the lysine codons AAA and AAG by stabilizing the codon- anticodon interaction. The precise decoding of the lysine codon by ms2-modification was particularly important for proinsulin synthesis because a lack of CDKAL1 significantly compromised the proper translation and processing of proinsulin [18]. Pancreatic beta cell-specific Cdkal1 knockout (KO) mice showed pancreatic islet hypertrophy, a decrease in insulin secretion, and impaired blood glucose control. Moreover, ER stress-related genes were upregulated in the pancreatic islets of the KO mice, and abnormal ER structure was observed. Furthermore, the KO mice were hypersensitive to HFD-induced ER stress but not to obesity [18], suggesting that pancreatic beta cell-specific Cdkal1 KO mice are an effective model for Asian-type T2D.
In the present study, we demonstrated the effect of pentadecanoic acid-based triglyceride purified from Aurantiochytrium on the glucose tolerance of pancreatic beta cell-specific CDKAL1 KO mice and the expressions of ER stress-related genes in the pancreatic islets.
Materials and Methods
Generation of Immortalized Pancreatic Beta Cells and Cell Culture
Immortalized pancreatic beta cells were generated from IT6 transgenic mice (kindly provided by Dr. Junichi Miyazaki) following a previously reported protocol (20). Briefly, an 8-week-old male mouse expressing the SV40 large antigen under the control of an insulin promoter was euthanized, and the insulinoma that had developed in the pancreas was excised under a microscope. The insulinoma tissue was then minced into small pieces and cultured in DMEM supplemented with 15% fetal bovine serum and ampicillin/ streptomycin. The cells were passaged more than 10 times to eliminate fibroblast cells and obtain immortalized pancreatic beta cells. The immortalized pancreatic beta cells were cultured and passaged in DMEM (Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific) and ampicillin/streptomycin (Thermo Fisher Scientific).
Isolation of Pentadecyl from Aurantiochytrium
Aurantiochytrium limacinum was incubated for 3 days and was then collected by centrifugation (3,000 G, 5 min). The collected Aurantiochytrium was washed with water and then added hexane under nitrogen displacement to extract the fat. The hexane solution was allowed to stand at 5°C to obtain white crystals. The solution was dissolved again by adding 5 times the amount of hexane, and the recrystallization was repeated to obtain white needle-like crystals of pentadecyl.
The main fatty acid component of pentadecyl was pentadeconic acid (C15:0) (> 73%) (Table 1).
| Content | Fatty acid composition | ||
|---|---|---|---|
| C42 | 2.10% | C15, C14, C13 | |
| C43 | 8.00% | C15, C13 | C15, C14 |
| C44 | 18.80% | C15, C14 | |
| C45 | 30.60% | C15 | |
| C46 | 22.30% | C15, C16 | |
| C47 | 11.70% | C15, C17 | C15, C16 |
| C48 | 4.50% | C15, C16, C17 |
Table 1: Fatty acid composition of pentadecyl. C13, tridecanoic acid; C14, tetradecanoic acid; C15, pentadecanoic acid; C16, hexa
Cell Viability Assay
Immortalized pancreatic beta cells were seeded in a 96-well plate at a density of 10,000 cells per well in DMEM medium supplemented with 10% fetal bovine serum. Twenty-four hours after seeding, cells were treated with 0, 1, 10, or 100 µg/ml pentadecyl for an additional 72 hr. WST- 8 reagent (Dojindo, Japan) was added to each well and the absorbance was measured at 405 nm.
Insulin Secretion Assay
Immortalized pancreatic beta cells were seeded in a 24- well plate at a density of 100,000 cells per well in DMEM medium supplemented with 10% fetal bovine serum. Twenty-four hours after seeding, the culture medium was replaced with fresh Krebs-Ringer Bicarbonate buffer (KRB buffer: 115 mM NaCl, 5 mM KCl, 10 mM NaHCO3, 2.5 mM MgCl2, 2.5 mM CaCl2, and 20 mM HEPES, pH 7.4, 0.1% bovine serum albumin) containing 2.8 mM glucose for 1 hr at 37°C. Cells were washed 3 times with 2.8 mM glucose in KRB buffer, followed by stimulation with 10 µg/ml pentadecyl in KRB buffer containing 2.8 mM glucose or 16.7 mM glucose for 30 min. As a control, cells were treated with 0.1% ethanol instead of pentadecyl. The supernatant from each well was saved and subject to ELISA assay. An ELISA kit (Shibayagi, Japan) was used to measure the insulin concentration according to the manufacturer’s instructions.
Rearing of Cdkal1 KO Mice and Intraperitoneally Glucose Tolerance Test (IPGTT)
Cdkal1flox/flox mice (18) were crossed with transgenic mice expressing Cre recombinase under the control of rat insulin 2 promoter (RIP-Cre) to generate pancreatic- specific Cdkal1 KO mice (KO). Their habitat was maintained at 22 ± 2°C with a 12-hr light/12-hr dark cycle. Mice were assigned to groups taking pentadecyl or dextrin (control). The pentadecyl group was given 6.7 µg of 10% pentadecyl dissolved with dextrin per 1 g of HFD (D12451, Research Diets) and the control group was given the same amount of dextrin instead of pentadecyl. KO mice aged 6 weeks old were given free-feeding for 9 successive weeks. A glucose tolerance test was performed after 3 and 9 weeks according to the following procedure. Mice were fasted for 12 hr and then intraperitoneally injected with 1 g/kg glucose (049- 31165, FUJIFILM Wako). Blood glucose was measured with a glucometer (Accu-Check Aviva Nano; Roche). All animal procedures were approved by the Animal Ethics Committee of Kumamoto University (approval ID; A2021-012).
Gene Expression Studies
Gene expression studies were performed as described previously [18]. Briefly, islets were isolated from KO mice or wild-type (WT) mice, which were fed an HFD for 9 weeks with pentadecyl or dextrin (control). Isolation of total RNA from islets was performed using an RNeasy mini kit (Qiagen). A PrimerScript RT reagent kit (TAKARA) was used to generate cDNA. Quantitative real-time PCRs were performed using either a Taqman Gene Expression Kit (Applied Biosystems) or SYBR Premix Ex Taq (TAKARA). The results were normalized to the level of GAPDH or beta actin.
Statistical Analysis
The data in all graphs are expressed as mean ± standard error of the mean. Comparisons between the two groups were analyzed using two-tailed unpaired Student’s t-tests. For multiple comparisons, a one-way analysis of variance was performed followed by Dunnet’s post-hoc analysis. A P-value of < 0.05 was considered significant.
Results
Assessment of the Toxicity Of Pentadecyl to Pancreatic Beta Cell
To investigate the toxicity of pentadecyl, immortalized pancreatic beta cells were treated with 1, 10, or 100 mg/ ml pentadecyl for 72 hr, and cell viability was accessed using WST-8 reagent. Compared with vehicle-treated cells, pentadecyl did not cause cell death. Notably, 10 or 100 mg/ ml pentadecyl potently induced cell viability (10 mg/ml pentadecyl, 110 ± 3.3; 100 mg/ml pentadecyl, 122 ± 2.6) (Figure 1A).
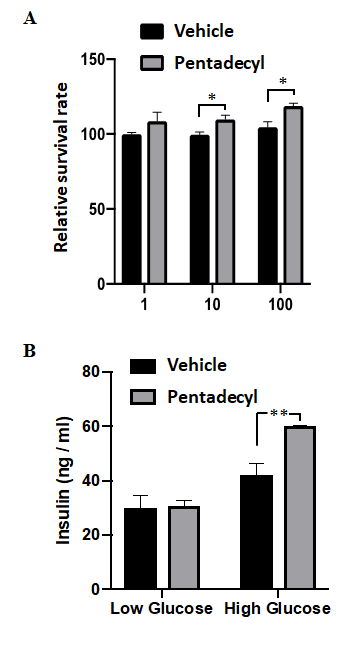
Figure 1: Demonstration of the toxicity of pentadecyl and its effect on insulin secretion in mouse pancreatic beta cells. (A) Toxicity assessment of the pentadecyl to pancreatic beta cells. Immortalized mouse pancreatic beta cells were treated with 1, 10, or 100 mg/ml pentadecyl for 72 hr. As a control, cells were treated with 0.1% ethanol (Vehicle). Then, cell viability was accessed using WST-8 reagent. n = 4 each. *P < 0.05. (B) Effect of pentadecyl on insulin secretion. Cells were treated with 10 µg/ml pentadecyl or 0.1% ethanol (Vehicle) in KRB buffer containing 2.8 mM glucose (Low Glucose) or 16.7 mM glucose (High Glucose) for 30 min. n = 4 each. **P < 0.01.
Assessment of the Effect of Pentadecyl on Insulin Secretion
We next investigated the effect of pentadecyl on insulin secretion. The immortalized mouse pancreatic beta cells were treated with 10 mg/ml pentadecyl and stimulated with either low glucose (2.8 mM glucose) or high glucose (16.7 mM glucose) for 30 min, and insulin release was examined using an ELISA kit. Pentadecyl did not affect insulin secretion when cells were stimulated with low glucose. In contrast, pentadecyl significantly enhanced insulin secretion when cells were stimulated with high glucose (vehicle, 42.3 ± 8.4 vs. pentadecyl, 59.4 ± 2.6) (Figure 1B).
Effect of Pentadecyl in Body Weight and Insulin Secretion in Pancreatic Beta Cell-Specific Cdkal1-Deficient Mice
Pancreatic beta cell-specific CDKAL1-deficient mice (KO), a model of Asian-type T2D, show glucose intolerance when fed an HFD [18]. Thus, we next investigated the effect of pentadecyl on insulin secretion in KO mice fed an HFD with pentadecyl or dextrin (control) for 9 successive weeks. There was no difference in weight gain between the pentadecyl or control group during the experimental period (Figure 2A).
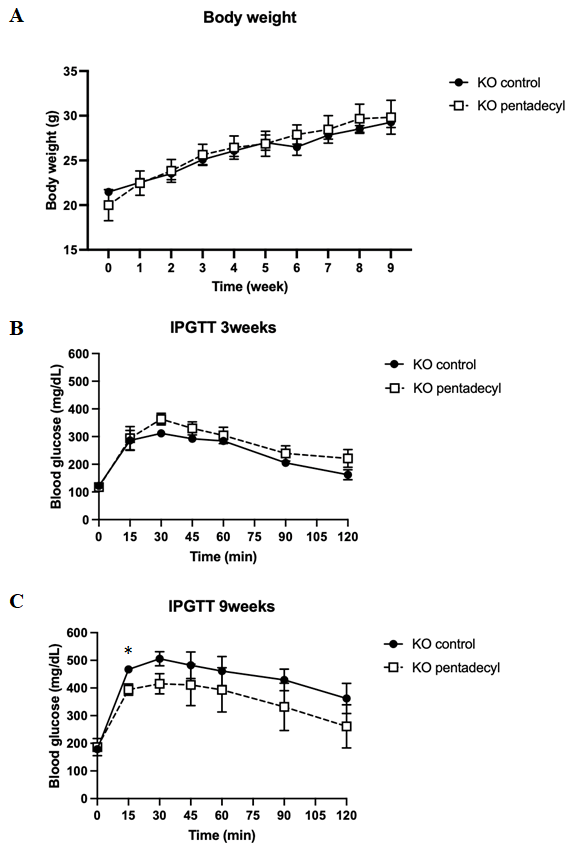
Figure 2: Effect of pentadecyl in body weight and insulin secretion in pancreatic beta cell-specific Cdkal1-deficient mice. (A) Effect of pentadecyl on body weight. The pentadecyl group was given 6.7 µg of 10% pentadecyl dissolved with dextrin per 1 g of HFD, and the control group was given the same amount of dextrin instead of pentadecyl. KO mice aged 6 weeks old were given free- feeding for 9 successive weeks. Mice body weights were measured every week. n = 4 each. Results of IPGTT at 3 (B) and 9 (C) weeks after feeding with HFD containing pentadecyl. As a control, mice were given dextrin instead of pentadecyl. n = 4–5. *P < 0.05.
The glucose tolerance in KO mice fed with pentadecyl was the same as that of control mice after 3 weeks on an HFD (Figure 2B). Both groups showed normal glucose tolerance. After 9 weeks, however, KO mice fed an HFD without pentadecyl showed glucose intolerance, whereas pentadecyl improved glucose intolerance (Figure 2C). The blood glucose level at 15 min after the intraperitoneal injection was lower in KO mice fed an HFD with pentadecyl than those fed an HFD without pentadecyl (control, 467.3 ± 9.7; pentadecyl, 394.0 ± 20.8) (Figure 2C).
Effect of Pentadecyl on the Expressions of ER Stress-Related Genes in the Islets of KO Mice Fed an HFD
In the islets of KO mice, the expressions of ER stress- related genes, namely XBP1, a spliced form of XBP1 (spliced XBP1), Bip, and Chop, increase when fed an HFD, and the elevated ER stress causes glucose intolerance in the mice [18]. Therefore, we next investigated whether pentadecyl improved the expressions of these ER stress-related genes in the islets of KO mice fed an HFD for 9 weeks. KO mice fed an HFD showed an increase in XBP1 (WT, 102.3 ± 15.8; KO, 200.3 ± 39.0), spliced XBP1 (WT, 94.0 ± 17.6; KO, 142.2 ± 15.7), Bip (WT, 101.3 ± 2.9; 128.7 ± 8.5), and Chop (WT, 96.3 ± 3.6; KO, 177.7 ± 34.1) compared with WT mice fed an HFD (Figure 3). Pentadecyl decreased the expression levels of XBP1 (KO without pentadecyl, 200.3 ± 39.0; KO with pentadecyl, 136.0 ± 8.0) and Chop (KO without pentadecyl, 177.7 ± 34.1; KO with pentadecyl, 136.0 ± 12.0) but not spliced XBP1 or Bip in the islets of KO mice (Figure 3).
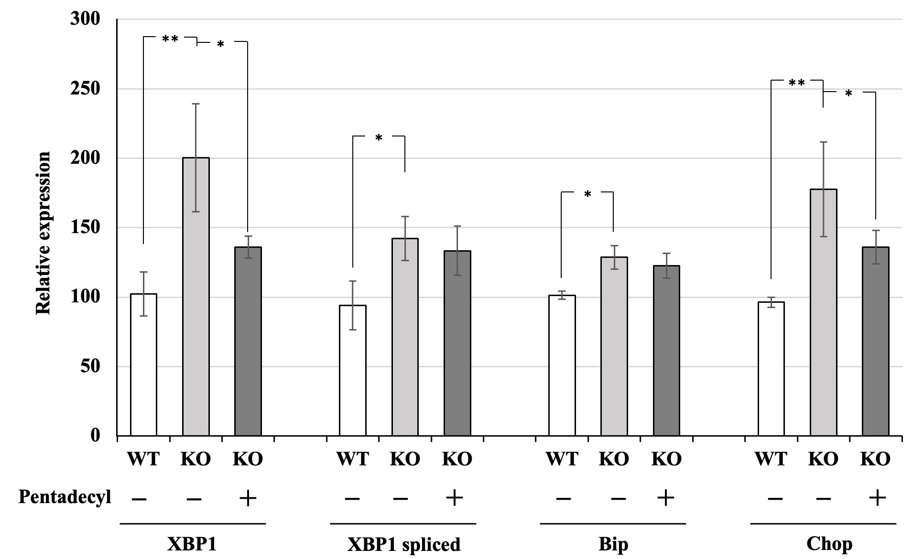
Figure 3 Effect of pentadecyl on the expressions of ER stress-related genes in the islets of KO mice fed an HFD. Relative expression of ER stress-related genes in the islets isolated from KO mice and WT mice, which were fed an HFD for 9 weeks with pentadecyl. P < 0.01; n = 4–5. As a control, mice were given dextrin instead of pentadecyl. n = 4–5. *P < 0.05, P < 0.01.
Discussion
The present study showed that pentadecyl induced high glucose-stimulated insulin secretion in immortalized mouse pancreatic beta cells and improved glucose intolerance in pancreatic beta cell-specific CDKAL1-deficient mice, a model of Asian-type T2D. Moreover, pentadecyl did not affect the development or body weight of the KO mice. Pentadecyl is a natural ingredient purified from Aurantiochytrium limacinum, a type of marine microalgae. Thus, pentadecyl may be a safe functional supplement for the improvement of glucose intolerance in Asian populations.
The fatty acid composition of pentadecyl mainly consists of pentadecanoic acid (C15:0), followed by tridecanoic acid (C13:0) and heptadecanoic acid (C17:0) [20]. Oral administration of pentadecanoic acid (C15:0) from dairy reduces circulating concentrations of the pro-inflammatory cytokines, glucose, and cholesterol in obese C57BL/6J mice fed an HFD [6]. These results suggest that the efficacy of pentadecyl is due to pentadecanoic acid (C15:0). Moreover, higher dietary intake and circulating levels of OCFAs have been associated with lower risks of chronic inflammation, cardiovascular disease, metabolic syndrome, and T2D [6, 7, 8, 9], suggesting that other OCFAs, such as tridecanoic acid (C13:0) and heptadecanoic acid (C17:0), in pentadecyl may also contribute for the improvement of glucose intolerance.
Pentadecyl decreased the expression of ER stress- related marker genes in pancreatic beta cell-specific CDKAL1-deficient mice, a model of Asian-type T2D. CDKAL1- mediated 2-methylthio modification in tRNALys(UUU) is essential for insulin synthesis in pancreatic beta cells because mistranslation frequently happens during lysine translation (18). Although proinsulin only contains two lysine residues, one of the lysine residues is located in the cleavage site between the A-chain and C-peptide. Mistranslation of these important residues can potentially generate cleavage- resistant proinsulin and lead to insulin insufficiency and accumulation of abnormal proinsulin in pancreatic beta cells, followed by ER stress [13]. However, a limitation of the present study is that we did not show a precise molecular mechanism for pentadecyl-dependent inhibition of ER stress in pancreatic beta cells of Cdkal1 KO mice. Further study is needed to demonstrate whether pentadecyl improves a decrease in the incorporation of lysine in proinsulin in the pancreatic islets of Cdkal1-KO mice and enhances pancreatic C-peptide contents. Additionally, a recent study showed that pentadecanoic acid (C15:0) improves mitochondrial dysfunction through the reduction of mitochondrial reactive oxygen species, a mediator of low insulin secretion, in stressed human cell systems [6]. These results suggest that pentadecyl enhances insulin secretion through the improvement of ER stress and mitochondrial dysfunction.
Pentadecyl did not affect the blood glucose level in the KO mice given pentadecyl for 3 weeks and decreased the blood glucose level only 15 min after the intraperitoneal injection but not after 30, 60, or 90 min in the KO mice given pentadecyl for 9 weeks. Furthermore, pentadecyl reduced the expressions of XBP1 and Chop but not spliced XBP1 or Bip in the islets of the KO mice given pentadecyl for 9 weeks. These results suggest that the effect of pentadecyl is partial. However, the present study was limited to only 9 weeks of investigation regarding the effect of pentadecyl in vivo. Thus, it is necessary to investigate the long-term effects pentadecyl application (e.g., > 12 weeks).
Conclusion
We demonstrated the effect of pentadecyl on insulin secretion both in pancreatic beta cells and pancreatic beta cell-specific Cdkal1 KO mice, a model mouse of Asian-type T2D. Pentadecyl induced high glucose-stimulated insulin secretion in the cells, improved glucose intolerance of the KO mice, and decreased the expressions of ER stress marker genes in the islets of the mice. Therefore, pentadecyl may be effective in the improvement of insulin secretion in patients with Asian-type T2D.
Conflict of Interest
KT has received research support from Refine Holdings Co., LTD.
Acknowledgments
We thank Dr. Jun-ichi Miyazaki (Osaka University) for providing IT6 transgenic mice.
Funding
This work was supported by Refine Holdings Co., LTD.
References
-
Rogero MM, Calder PC (2018) Obesity, Inflammation, Toll-Like Receptor 4 and Fatty Acids. Nutrients 10(4): 432.
-
Hwang DH, Kim JA, Lee JY (2016) Mechanisms for the activation of Toll-like receptor 2/4 by saturated fatty acids and inhibition by docosahexaenoic acid. Eur J Pharmacol 785: 24-35.
-
Rocha DM, Caldas AP, Oliveira LL, Bressan J, Hermsdorff HH (2016) Saturated fatty acids trigger TLR4-mediated inflammatory response. Atherosclerosis 244: 211-215.
-
Forouhi NG, Koulman A, Sharp SJ, Imamura F, Kröger J, et al. (2014) Differences in the prospective association between individual plasma phospholipid saturated fatty acids and incident type 2 diabetes: the EPIC-InterAct case-cohort study. Lancet Diabetes Endocrinol 2(10): 810-818.
-
Kurotani K, Sato M, Yasuda K, Kashima K, Tanaka S, et al. (2017) Even- and odd-chain saturated fatty acids in serum phospholipids are differentially associated with adipokines. PLoS One 12(5): e0178192.
-
Venn Watson S, Lumpkin R, Dennis EA (2020) Efficacy of dietary odd-chain saturated fatty acid pentadecanoic acid parallels broad associated health benefits in humans: could it be essential?. Sci Rep 10(1): 8161.
-
Huang L, Lin JS, Aris IM, Yang G, Chen WQ, et al. (2019) Circulating Saturated Fatty Acids and Incident Type 2 Diabetes: A Systematic Review and Meta-Analysis. Nutrients 11(5): 998.
-
Khaw KT, Friesen MD, Riboli E, Luben R, Wareham N (2012) Plasma phospholipid fatty acid concentration and incident coronary heart disease in men and women: the EPIC-Norfolk prospective study. PLoS Med 9(7): e1001255.
-
Krachler B, Norberg M, Eriksson JW, Hallmans G, Johansson I, et al. (2008) Fatty acid profile of the erythrocyte membrane preceding development of Type 2 diabetes mellitus. Nutr Metab Cardiovasc Dis 18(7): 503-510.
-
Månsson HL (2008) Fatty acids in bovine milk fat. Food Nutr Res 52.
-
Kaya K, Nakazawa A, Matsuura H, Honda D, Inouye I, et al. (2021) Thraustochytrid Aurantiochytrium sp. 18W-13a accummulates high amounts of squalene. Biosci Biotechnol Biochem 75(11): 2246-2248.
-
Kaya K, Shiraishi F, Iida T, Yamada M, Sano T (2021) An acidic morpholine derivative containing glyceride from thraustochytrid, Aurantiochytrium. Scientific Reports 11(1): 6868.
-
Wei FY, Tomizawa K (2018) tRNA modifications and islet function. Diabetes Obes Metab 20(2): 20-27.
-
Wei FY, Tomizawa K (2011) Functional loss of Cdkal1, a novel tRNA modification enzyme, causes the development of type 2 diabetes. Endocr J 58(10): 819- 825.
-
Takeuchi F, Serizawa M, Yamamoto K, Fujisawa T, Nakashima E, et al. (2009) Confirmation of multiple risk Loci and genetic impacts by a genome-wide association study of type 2 diabetes in the Japanese population. Diabetes 58(7): 1690-1699.
-
Steinthorsdottir V, Thorleifsson G, Reynisdottir I, Benediktsson R, Jonsdottir T, et al. (2007) A variant in CDKAL1 influences insulin response and risk of type 2 diabetes. Nat Genet 39(6): 770-775.
-
Fukushima M, Usami M, Ikeda M, Nakai Y, Taniguchi A, et al. (2004) Insulin secretion and insulin sensitivity at different stages of glucose tolerance: a cross-sectional study of Japanese type 2 diabetes. Metabolism 53(7): 831-835.
-
Wei FY, Suzuki T, Watanabe S, Kimura S, Kaitsuka T, et al. (2011) Deficit of tRNA(Lys) modification by Cdkal1 causes the development of type 2 diabetes in mice. J Clin Invest 121(9): 3598-3608.
-
Zhou B, Wei FY, Kanai N, Fujimura A, Kaitsuka T, et al. (2014) Identification of a splicing variant that regulates type 2 diabetes risk factor CDKAL1 level by a coding- independent mechanism in human. Hum Mol Genet 23(17): 4639-4650.
-
Miyazaki J, Araki K, Yamato E, Ikegami H, Asano T, et al. (1990) Establishment of a pancreatic beta cell line that retains glucose-inducible insulin secretion: special reference to expression of glucose transporter isoforms. Endocrinology 127(1): 126-132.
-
Kaya K (2020) Method for producing odd-numbered fatty acid ester, and culture composition.
- Investigation of Polymorphisms in PPAR-Ɣ and TRHR Genes and their Impact on Turkish Diabetic and Obese Individuals
- The Impact of Aircraft Noise Exposure on the Efficacy of Empagliflozin Therapy in an Animal Model of Obesity
- Rooibos Mitigates Metabolic and Inflammatory Dysfunctions in Mice Fed a High-Carbohydrate Diet
- Synergistic Effect of Combined Leaf Extract of Vernonia amygdalina, Ocimum gratissimum, and Zingiber officinale Tuber on Phytochemical Profile, Antioxidant Activity, Serum Insulin, and Biochemical Parameters in Streptozotocin-Induced Diabetic Rats
- Investigation of Cardiovascular Responses to Aerobic Exercise in Obese University Students
- A Look at the Phase Angle Obtained by Electrical Bioimpedance