Potential Protective Effect of Empagliflozin on αKlotho/Autophagy Protein LC3 in Diabetic Retinopathy: Evidence from Diabetic Mice Model
Introduction: To study the protective efficacy of Empagliflozin (EMPA), a novel sodium glucose transporter inhibitor antidiabetic drug, on the expression of α-klotho (αKL) protein and of the autophagy key proteins Light chain 3 (LC3) in type 2 diabetic mellitus (T2DM) mice diabetic retinopathy (DR). Materials and Methods: We used the BTBR mouse strain with the ob/ob leptin-deficiency mutation that develops spontaneously severe T2DM, C57/BL mice used as control. EMPA was administrated to the diabetic mice via drinking water for a period of 12 weeks. At the end of the experiment, mice retinas were removed and subjected to further histological analysis: Immunohistochemistry and Immunofluorescence staining for αKL, LC3, protein expression level. Results: Retinal αKL protein expression levels were lower in diabetic mice than control (11.94±4.6 vs 48.4±5.33) % ** P< 0.01), which were restored to near normal with EMPA treatment compared to DM mice (35+20.4% vs 11.94±4.6%,*P
Introduction
Diabetic retinopathy (DR) remains a leading cause of blindness, and its incidence is increasing worldwide due to the luck of achieving near normal blood glucose levels [1, 2]. Beyond glycemic control, most therapies that target the late proliferative stage of the DR are effectuated late, at a time when vision is already incurable [3, 4, 5].
DR is a very common long-term complication of diabetes mellitus (DM). Two types of DR were described, the background DR type and the serious proliferative type [6]. The latter is characterized by an abnormal growth of small blood vessels in the eye due to local hypoxia and increased synthesis of vascular endothelial growth factor ) VEGF), neo-vascularization at disc, vitreous hemorrhages that ends in retinal detachment and loss of vision [7].
Hyperglycemia triggers pro-inflammatory mediators, which activate oxidative stress pathways with increased production of free reactive oxygen species and further amplify the inflammatory signal through multiple pathways [7, 8]. The chronic activation of inflammatory cascades results in retinal microvasculature damage. Chronic hyperglycemia also triggers and increases synthesis and production of VEGF due to hypoxia, with increased retinal angiogenesis leading to retinal damage [7, 9].
The α-klotho (αKL) gene is an anti-aging gene, coding for a single-pass transmembrane protein with multiple pleiotropic effects [10, 11, 12, 13, 14], and is highly expressed in the kidneys, retina, and is affected by pathophysiological conditions including oxidative stress found in DM and its associated micro-vascular complications. The extracellular domain of αKL is cleaved by proteolytic enzymes and released into the circulation as soluble αkL [13]. Studies have shown that αKL can play role as protective against vascular disease by reducing oxidative stress and inflammatory processes [14]. It was published that klotho silencing induced oxidative stress and inflammatory response, which results in genomic instability, endoplasmic reticulum stress and retardation of the autophagy process with induction of apoptotic cell death [15, 16].
Autophagy is the primary intracellular mechanism mediating degradation and recycling of proteins and organelles and catabolic process in which cellular components are degraded by lysosomes [9]. Autophagy is essential for retinal development and vision formation [17, 18]. Autophagy, a well-coordinated multi-step process regulated by autophagy-related gene (Atg) products. Initially, a phagophore forms around cytoplasmic components that are then sequestered by a double membrane, which forms the autophagosome. The autophagosome subsequently fuses with the lysosome to form an autolysosome; the enclosed contents are then degraded.
Autophagy can be measured by microtubule-associated proteins 1A/1B light chain 3B (LC3-II) protein expression levels. In the course of autophagy, the microtubule- associated protein 1 light chain 3 (LC3-I) is transformed to LC3-II, which permits the combination of the protein to autophagosome membranes [17, 19, 20, 21]. So, light chain 3 (LC3) is a key autophagy proteins, which have a central role in the pathogenesis of DM vascular complications.
A new anti-diabetic class of drugs has been developed for patients with T2DM that targets the sodium-glucose cotransporter type 2 (SGLT2), which is mainly expressed in renal proximal convolute tubules [22]. Inhibition of the SGLT2 by empagliflozin (EMPA) is a very selective and potent SGLT2 inhibitor lowers plasma glucose levels mainly by increasing urinary glucose excretion and is associated with additional non-glycemic benefits including reduction in blood pressure and body weight, decrease urinary albumin [22, 23]. EMPA also has pleiotropic effects such as decreased the production of inflammatory markers. New therapies that target the early pathogenesis of DR are needed. Despite these recent advances, the effects of αKL and LC3 proteins on retinal health have not yet been examined extensively.
In this study, BTBR mice with the ob/ob leptin- deficiency and wild-type C57BL/6 male were purchased from the Jackson Laboratory. BTBR (ob/ob) mice have been recently described as an excellent animal model for diabetic nephropathy and other microvascular complications [24].
Materials and Methods
Animals
All animal experiments were conducted according to the institutional animal ethical committee guidelines, which conform to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (Eighth edition 2011) (Ethics 48-07-2019). The animals were maintained at the animal facility of the Galilee Medical Center, Nahariya, Israel. They were maintained at a constant temperature and relative humidity under a regular light- dark schedule (12h: 12h), fed with normal rodent diet and with tap water ad libitum.
Study Design
In the study, BTBR mice with the ob/ob leptin- deficiency and wild-type C57BL/6 male were purchased from the Jackson Laboratory. BTBR (ob/ob) mice have been recently described as an excellent animal model for diabetic nephropathy [24]. BTBR ob/ob mice develop progressive proteinuria and a renal histomorphological picture that is quite similar to that seen in human patients with advanced diabetic nephropathy.
BTBR mice of 20–25 g body wt. (~2 months of age) were randomly divided into 2 groups: 15 BTBR ob/ob mice with no treatment (DM) and 15 BTBR ob/ob mice that were treated with EMPA (DM+EMPA), 15 wild-type C57BL/6 male mice were used as control (C57/Bl). Weekly, mice were weighed and their blood glucose was measured using Glucometer (Accu-chek, Roche). Every three weeks mice were held for 24 hours, in metabolic cages (five times) for partial kidney function evaluation. All mice were sacrificed, 13 weeks after the beginning of the experiment, and selected organs were harvested and preserved for later biochemical or histological analyses.
Kidney Function
To assess partial kidney function, urine was collected by placing each mouse in a metabolic chamber (Techniplast S.p.A., Buguggiate, Italy) for 24 h. Urine output volume, 24- hrs protein and creatinine excretion water intake, as well as body weight (BW) were evaluated. Urine protein and creatinine concentrations were biochemically analyzed.
Drug Treatment
EMPA was purchased from DA-TA Biothec, USA. EMPA was supplied to mice in the dosage of 1mg/kg/day, which was dissolved in the drinking water. We have estimated daily water consumption, according to the data derived from the metabolic cages. Sacrifice, Organ Harvesting and Preservation Under deep anesthesia by overdose of Ketamine-Xylazine injection mice were killed, and an eyeball was harvested and underwent consecutive immersion in PBS then immersed in 20% sucrose-PBS for 12 h at 4°C and then frozen in O.C.T. embedding medium (Miles, Elkhart, USA) and kept in -80°C for cryosection. Retinal tissues were selected randomly from each group, and were sectioned using cryostat into 16µm sections and mounted on positively charged histological slides [24].
Histological and Immunohistochemistry Staining
Sections were incubated overnight at 4°C with primary antibodies at the following dilutions in blocking solution (CAS-block, Invitrogen): αKL (1:400; LSBio, Seattle, WA. USA), Rat anti-αKL antibodies were purchased from TransGenic.
Anti-GAPDH (1 : 20,000, EPR 16891, Abcam, UK). Rabbit monoclonal anti-LC3A/B (D3U4C) (1:100 Cell Signaling Technology Israel). Secondary antibodies used were donkey anti-rat TRITC-linked from Invitrogen; and goat anti-rabbit HRP-linked and goat anti-mouse HRP-linked (Cell Signaling Technology, Israel) [24].
Klotho Expression in the Retina
To determine whether αKL expression is detected in retina, we performed IHC simultaneously on retinal sections from C57/Bl mice, DM mice and DM+EMPA mice. Confocal microscopy with orthogonal projections to detect αKL protein expression (red) and DAPI (nucleus) in a single optical section (0.35 µm) from the GCL retina. Quantification of klotho signal in GCL was conducted across multiple retinas.
Finally, slides were covered, underwent scanning at ×400 magnification light microscope LSM 700 laser scanning confocal microscope (Carl Zeiss Meditec, Germany), and photographed using the Tecnai 12 BioTwin TEM (FEI) equipped with a SIS MegaView3 CCD camera (Surface Imaging Systems). The images were processed using Adobe Photoshop CS (Adobe Systems). For each mouse’s retina slide, five different fields of view were randomly selected, the total number of pixels with specific fluorescence of the designated antibody was counted, and then divided by the total ganglionic cell layer (GCL) tissue area to achieve the percentage of fluorescence.
Statistical Analysis
Data are presented as mean±SD. Comparisons between groups were performed by 2-way analysis of variance (ANOVA) with repeated measures, in which the treatment and the time point were the independent variables. Single measured data i.e., final measurements and histological analysis were compared between groups using 1- way ANOVA, in which the treatment was the independent variable, whenever the ANOVA was significant, a multiple comparison was performed using the Holm-Sidak as post-hoc test. P value of <0.05 was considered significant.
Results
Mice General Parameters
At baseline (BL), the average DM and DM+EMPA mice BW was 37.6±0.9 gr. and the C57/Bl mice were 23.4±0.9 gr., consistently throughout the study the C57/Bl mice were liner than the DM and the DM+EMPA mice (Figure 1A). The mice consumed 3.9±0.5 ml/day, 6.4±0.5 ml/day and 5.3±0.5 ml/day water in the DM, DM+EMPA and C57/Bl groups, respectively, though EMPA was not yet administered, higher water consumption was found in the DM+EMPA group (Figure 1B). Initial urine outputs for the DM and the DM+EMPA groups were 2.07±0.26 ml/day and 1.81±0.26 ml/day, respectively. The C57/Bl group urine output was significantly lower, 0.98±0.26 ml/day, (P<0.05 vs. both DM groups, Figure 1C).
At one and two months the DM mice (DM and DM+EMPA groups) gain weight considerably (P<0.01 vs. BL and C57/ Bl, Figure 1A). Interestingly, at two month this group was significantly, lighter than the DM group (P<0.05, Figure 1A). Water intake was preserved at 1 one month but increased significantly at 2 months (P<0.001, Figure 1B). Urine output of the DM mice was preserved at 2.32±0.26 ml/day and 2.26±0.26 ml/day at one and two months, respectively (P>0.05, Figure. 1C). Yet, For the DM+EMPA mice group, urine output increased as soon as one month of treatment reaching (3.84±0.26 ml/day and 4.45±0.26 ml/day at one and two months, respectively, P<0.001). Urine composition, i.e. urine protein, creatinine and glucose, were all preserved along the experiment in the DM and C57/Bl groups.
The DM+EMPA group’s urine composition changed dramatically at one and two month of experiment. Urine protein and urine creatinine were 72.7±9.7 mg/dl and 58.9±9.7 mg/dl and 12.8±2.8 mg/dl, and 13.2±2.8 mg/ dl (P<0.05 vs. DM and BL) at one month and two months measurements, respectively (Figure 1D and 1E). On the other hand, urine glucose levels increased to 13038.5±702.9 mg/dl and 10402±702.9 mg/dl (P<0.001, Figure 1F).
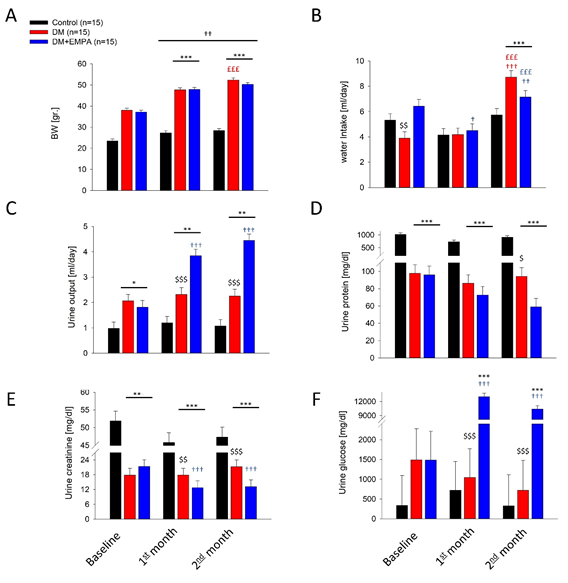
Figure 1: Mice general maintenance physiological parameters. A. mean weight, B. water intake, C. urine output, D. Urine protein exertion, E. urine creatinine, F. urine glucose level. C57 group (black columns), DM mice (red columns), DM+EMPA (blue columns). * P<0.05 vs. C57, P<0.01 vs. C57, * P<0.001 vs. C57, $ P<0.05 vs. DM+EMPA, $$ P<0.01 vs. DM+EMPA, $$$ P<0.001 vs. DM+EMPA, £££ P<0.001 vs. one month, † P<0.05 vs. BL, †† P<0.01 vs. BL, ††† P<0.001 vs. BL. Data represent the mean ± SEM.
Retinal Structure and αKL Expression
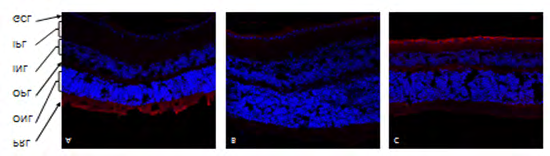
αKL expression in ganglionic cell layer (GCL) of the retina, showing representative immunofluorescence sections in C57 mice retina (Figure 2A) , DM mice retina (Figure 2B). DM+EMPA mice retina (Figure 2C). Red indicates αKL positive pixels, Blue- nuclei detected by DAPI.
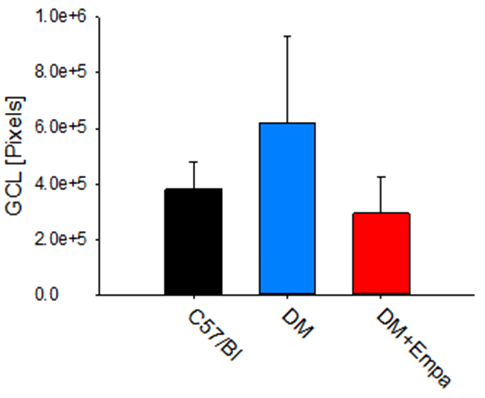
There was no significant difference between the three groups’ GCL size, as measured by their number of pixels in the microscopic photographs, indicating comparable GCL layer in all mice groups. Specifically, C57/Bl mice’s GCL layer was occupying 380055±100281 pixels, the DM’s average GCL layer was of 617963±312261 pixels, and in the DM+EMPA groups it was 293302±129155 pixels (P>0.05, Figure 3).
αKL Expression Measured by Number of Pixels in GCL Retina
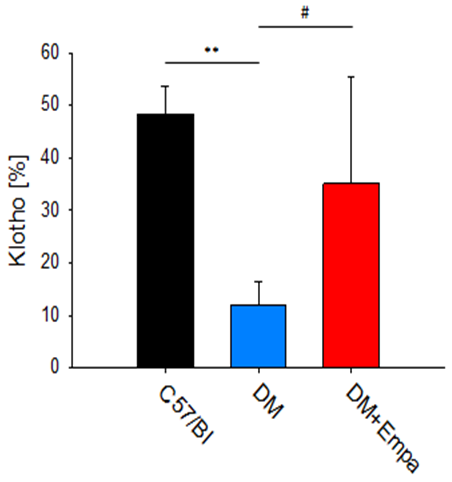
αKL was detected in C57/Bl group (Figure 2A), its expression was significantly lower in DM (Figure 2B) and restored toward control level in DM+EMPA mice (Figure 2C).
Upon stain measurements, αKL occupied, 65285±26099 pixels and 181494±42479 pixels, in the DM mice and C57/Bl, respectively (P<0.01). EMPA treatment to DM mice increased the number of αKL positive pixels yet, their number was still lower compared to the C57/Bl, 89253±62352 pixels vs. 181494±42479 pixels, respectively (P<0.05, Figure 4).
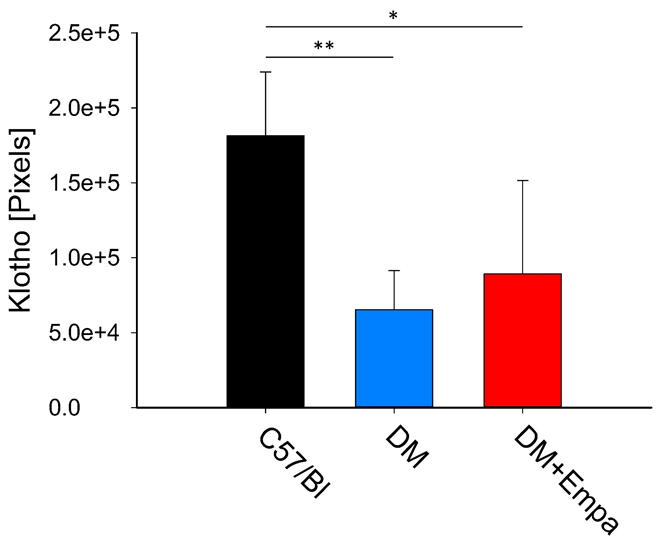
When expressed as percent of stained tissue area, a Significant decrease of αKL was found in the DM mice group vs. C57/Bl mice group 11.9±4.6% vs. 48.4±5.3% (P< 0.01), and vs. the DM+EMPA 35.0+20.4% (P<0.05, Figure
5). Collectively, indicating, that though GCL structure was comparable between the experimental groups, αKL expression was significantly affected by DM and to some extent by the EMPA treatment.
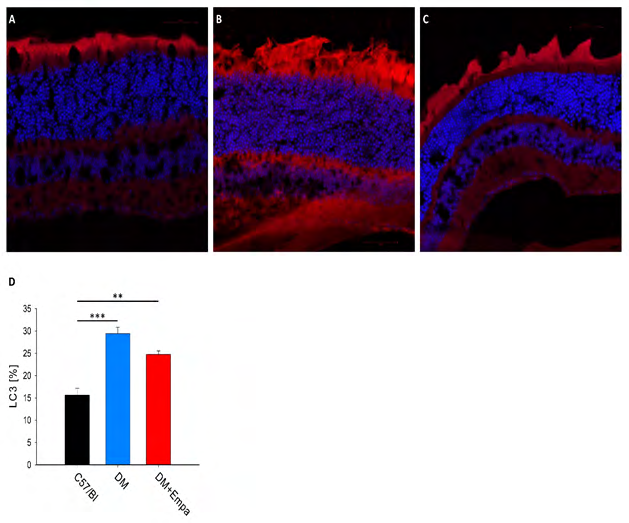
LC3 Expression % in GCL Retina
In the representative photographs LC3 seems to be highly expressed in DM mice (Figure 6A), and to lower extent in the DM+EMPA treated mice (Figure 6B), yet C57/ Bl mice showing the lower level of LC3 expression (Figure 6A). Analysis of the photographs show significant increase in the LC3 expression area in the DM group vs. the C57/Bl group (29. 4±1.4 % vs. 15.6±1.6 %, P<0.001, Figure 6D), EMPA treatment increased significantly LC3 expression vs.
the C57/Bl group 24.8±0.8% vs. 15.6±1.6%, P<0.01, Figure 6D). Nevertheless, LC3 expression was comparable among DM and DM+EMPA mice (P>0.05, Figure 6D).
Discussion
DR is the leading cause of blindness in the general population and the elderly [3, 4]. It is well known that retinal function deterioration in proliferative DR is irreversible. Under diabetic conditions, hyperglycemia and the resulting glucose-toxicity are the driving forces for inflammatory processes in the retina [6, 25]. Hyperglycemia activates oxidative stress and inflammatory pathway, concomitantly to increase of reactive oxygen species production, all lead to injury to the retinal cells.
The soluble αKL is expressed in multiple tissues, with particularly high levels in the kidney and retina [13], and acts as a humoral factor that targets multiple tissues and organs with antiaging effects, anti-inflammatory and antioxidant activity [26, 27, 28]. Different studies suggest that klotho protein, which decreased in early stages of DN, and has an important role in the pathogenesis of DM and its vascular complications such as DR [11].
Autophagy process has a central role in the maintenance of cell hemostasis, and acts as a protective mechanism against oxidative stress, that may prevent cell death, especially in DM and DR [17, 29, 30]. The key protein LC3 is one of the important autophagy proteins involved in the final formation of the auto-phagosome with a protective role.
Our Lab had demonstrated that in diabetic mice model, there is decreased expression and abundance of the defensive protein αklotho, and down regulation of the autophagy proteins LC3 and ATG5 in diabetic nephropathy in diabetic mice model [17, 29].
Different investigators had published that treatment of BTBR ob/ob mice with the new anti-diabetic drug EMPA (SGLT2i) led to reduced inflammation, as demonstrated by the significant reduction in mRNA expression of the pro- inflammatory cytokines MCP-1, RANTES, and IL-6 [29, 30, 31]. Further, studies demonstrated that SGLT2is suppress hyperglycemia, restored β cell mass, macrophage infiltration, gene expression of inflammation, oxidative stress and apoptosis in diabetic db/db mice [29, 30, 31]. Yet, less is known on the role of αKL and autophagy protein LC3 in diabetic mice with retinopathy treated with SGT2Is such as EMPA.
Few studies have investigated changes in the expression of αKL in GCL retina. Baolan, et al. (15) demonstrated that the serum αKlotho level was independently and negatively correlated with retinopathy in type 2 diabetes patients, and other showed that Klotho can ameliorate apoptosis via the ERK1/2-p38 kinase signaling pathway [11, 14]. Different pathophysiological factors in vivo could affect klotho gene expression, among them hyperglycemia, oxidative stress, inflammation and angiotensin II significantly inhibited klotho gene expression [32]. Their results showed that the serum klotho protein level was evidently reduced in diabetes, which was consistent with previous other studies [33, 34]. The data suggested that klotho protein was likely to be a new and important target for DR.
Antonella and her group [34] demonstrated that in T2DM patients, lower circulating levels of the vascular protective hormone Klotho are associated with increased risk of progression of DR. In C57BL/6 male mice, disruption of leukotriene signaling by pharmacologic blockade inhibited early DR. Before pathologic lesions develop, the administration of montelukast impeded the preceding diabetes-induced cellular dysfunction, significantly reducing retinal vascular permeability, pro-inflammatory leukotriene generation, and superoxide accumulation. Over time, these early beneficial effects correlated with long-term protection of the retinal capillaries and neurons [35].
The association between klotho, apoptosis and autophagy has been reported in various diseases, such as Alzheimer’s disease and cancer [15]. Chen and his group [36] investigated, whether the autophagy process plays a role in Klotho gene deficiency-induced arterial stiffening and hypertension. They demonstrated that Klotho gene deficiency increased autophagy, as evidenced by increased expression of LC3B-II and autophagy in klotho gene deficient mice [29], however, whether Klotho deficiency directly induces autophagy is not clear, but they conclude that Klotho directly regulates autophagy [36].
Excessively stimulated autophagy in retinal cells may cause autophagy death cells, which may lead to advanced stages of DR. Therefore, preventing excessive stimulated autophagy at early stages by new treatments may be of benefit to patients with DR.
The aim of our study was to assess the role of klotho protein and LC3 protein, in the pathophysiology of DR. Further, we looked into the protective effects of the new anti-diabetic drug, the SGLT2i, on DR. Finally, we evaluated the correlation between αKL expression in ob/ob mice with DM, and DM mice treated with SGLT2i (EMPA) to gain novel insight into the underlying functions of Klotho and autophagy in preventing or slowing the progression of DR. Our results showed that αKL in the DM mice was decrease in the retinal GCL and was restored toward normal by EMPA treatment. Concomitantly, we found a significant increase in the LC3 protein expression probably as a compensatory response to decreased αKL levels, to protect retinal cell damage, which was restored also by EMPA treatment [26, 27].
Conclusion
Our study demonstrates that the anti-aging protein αKL plays a critical role in retinal pigment epithelium (RPE), function, especially the GCL. Our study revealed significant decrease in retinal αKL protein expression in T2DM mice model, and restored by EMPA treatment. It appears that Klotho may be involved in several physiological processes such chronic hyperglycemia in T2DM vascular complications [37], that can affect the progression of DR. On the other side we found a significant increase in LC3 protein expression in the retinal GCL in the diabetic mice, and downregulated to basal levels by EMPA treatment. The effects of high glucose on decreased klotho protein levels in retinal GCL, is followed by excessively stimulated autophagy in retinal cells with increased LC3 protein levels, may cause autophagy death cells, which may lead to advanced stages of DR and loss of vision. Exogenous supplementation with Klotho may be a potential therapeutic target in the future for inhibiting the progression of DR. However, further studies are needed to elucidate the role of Klotho in the pathophysiological mechanisms of the development and progression of T2DM [38, 39, 40].
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability
The data presented in this study are available on request from the corresponding author.
Ethical Approval
The study was conducted according to the guidelines of the animal committee, Faculty of Medicine, Bar-Ilan University.
Authors’ Contributions
Nakhoul Farid and Nakhoul Nakhoul prepared the manuscript, Nakhoul Rola reviewed the manuscript, Nakhoul Nakhoul and Shaul Attar conceptualized the study; Ertracht Offir, Igbariye Anas, Ranin Saed formulated the methodology.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Ministry of development of the Negev and the Galilee
References
-
Cheung N, Mitchell P, Wong TY (2010) Diabetic retinopathy. Lancet 376(9735): 124-136.
-
Heng LZ, Comyn O, Peto T, Tadros C, Ng E, et al. (2013) Diabetic retinopathy: pathogenesis, clinical grading, management and future developments. Diabet Med 30(6): 640-650.
-
Kollias AN, Ulbi MW (2010) Diabetic retinopathy: Early diagnosis and effective treatment Dtsch Arztebl Int 107(5): 75-78.
-
Ebneter A, Zinkernagel MS (2016) Novelties in Diabetic Retinopathy. Endocr Dev 31: 84-96.
-
Ghamdi AHA (2020) Clinical Predictors of Diabetic Retinopathy Progression; A Systematic Review. Curr Diabetes Rev 16(3): 242-247.
-
Wang W, Lo ACY (2018) Diabetic Retinopathy: Pathophysiology and Treatments. Int J Mol Sci 19(6): 1816.
-
Gupta N, Mansoor S, Sharma A, Sapkal A, Sheth J, et al. (2013) Diabetic Retinopathy and VEGF. The Open Ophthalmology Journal 7: 4-10.
-
Sfi SZ, Qvist R, Kumar S, Batumalaie K, Ismail ISB (2014) Molecular Mechanisms of Diabetic Retinopathy, General Preventive Strategies, and Novel Therapeutic Targets. Biomed Res Int 2014: 801269.
-
Aiello LP, Wang JS (2000) Role of vascular endothelial growth factor in diabetic vascular complications. Kidney International 58(77): S113-S119.
-
Kokkinaki M, Abu-Asab M, Gunawardena N, Ahern G, Javidnia M, et al. (2013) Klotho Regulates Retinal Pigment Epithelial Functions and Protects Against Oxidative Stress. The Journal of Neuroscience 33(41): 16346-16359.
-
Reish NJ, Maltare A, McKeown AS, Laszczyk AM, Kraft TW, et al. (2013) The Age-Regulating Protein Klotho Is Vital to Sustain Retinal Function. Invest Ophthalmol Vis Sci 54: 6675-6685.
-
Chen CD, Sloane JA, Li H, Aytan N, Giannaris EL, et al. (2013) The antiaging protein Klotho enhances oligodendrocyte maturation and myelination of the CNS. J Neurosci 33: 1927-1939.
-
Matsumura Y, Aizawa H, Shiraki-Iida T, Nagai R, Kuro-o M, et al. (1998) Identification of the human klotho gene and its two transcripts encoding membrane and secreted klotho protein. Biochem Biophys Res Commun 242(3): 626-630.
-
Ji B, Wei H, Ding Y, Liang H, Yao L, et al. (2020) Protective potential of klotho protein on diabetic retinopathy: Evidence from clinical and in vitro studies. J Diabetes Investig 11(1): 162-169.
-
Mytych J, Solek P, Koziorowski M (2019) Klotho modulates ER mediated signaling crosstalk between pro-survival autophagy and apoptotic cell death during LPS challenge. Apoptosis 24(1-2): 95-107.
-
Herat LY, Matthews VB, Rakoczy PE, Carnagarin R, Schlaich M (2018) Focusing on Sodium Glucose Cotransporter-2 and the Sympathetic Nervous System: Potential Impact in Diabetic Retinopathy. Int J Endocrinol 2018: 9254126.
-
Gao X, Du Y, Lau WB, Li Y, Siquan Zhu S, et al. (2021) Atg16L1 as a novel biomarker and autophagy gene for diabetic retinopathy. Journal of diabetic research 2021:5398645.
-
Dehdashtian E, Mehrzadi S, Yousefi B, Hosseinzadeh A, Reiter RJ, et al. (2018) Diabetic retinopathy pathogenesis and the ameliorating effects of melatonin; involvement of autophagy, inflammation and oxidative stress. Life Sci 193: 20-33.
-
Koch EAT, Nakhoul R, Nakhoul F, Nakhoul N (2020) Autophagy in diabetic nephropathy: A Revirw. Int Urol Nephrol 52(9): 1705-1712.
-
Lu Y, Xu X, Zhonghua YK, et al. (2012) The latest advance of the relationship between autophagy and diabetic retinopathy. Zhonghua Yan Ke Za Zhi 48(7):649-52.
-
Mytych J, Sołek P, Będzińska A, Rusinek K, Warzybok A, et al. (2020) Klothomediated changes in the expression of Atg13 alter formation of ULK1 complex and thus initiation of ER and Golgistress response mediated autophagy. Apoptosis 25(1-2): 57-72.
-
Herat LY, Ward NC, Magno AL, Rakoczy EP, Kiuchi MG, et al. (2020) Sodium glucose co-transporter 2 inhibition reduces succinate levels in diabetic mice. World J Gastroenterol 26(23): 3225-3235.
-
Allon G, Mann I, Remez L, Sehn E, Rizel L, et al. (2019) PRCD is concentrated at the base of photoreceptor outer segments and is involved in outer segment disc formation. Human Molecular Genetics 28(24): 4078- 4088.
-
Gembardt F, Bartaun C, Jarzebska N, Mayoux E, Todorov VT, et al. (2014) The SGLT2 inhibitor empagliflozin ameliorates early features of diabetic nephropathy in BTBR ob/ob type 2 diabetic mice with and without hypertension. Am J Physiol Renal Physiol 307(3): F317-F325.
-
Engerman RL, Kern TS (1986) Hyperglycemia as a cause of diabetic retinopathy. Metabolism 35(4): 20-23.
-
Kurosu H, Yamamoto M, Clark JD, Pastor JV, Nandi A, et al. (2005) Suppression of aging in mice by the hormone Klotho. Science 309(5742): 1829-1833.
-
Mencke R, Hillebrands JL (2017) The role of the anti- ageing protein Klotho in vascular physiology and pathophysiology. Ageing Res Rev 35: 124-146.
-
Yassin R, Tadmor H, Farber E, Igbariye A, Armaly- Nakhoul A, et al. (2021) Alteration of autophagy-related protein 5 (ATG5) levels and Atg5 gene expression in diabetes mellitus with and without complications. Diab Vasc Dis Res 18(6).
-
Cai X, Li J, Wang M, She M, Tang Y, et al. (2017) GLP-1 Treatment Improves Diabetic Retinopathy by alleviating Autophagy through GLP-1R-ERK1/2-HDAC6 Signaling Pathway. International Journal of Medical Sciences 14(12): 1203-1212.
-
Choi AMK, Ryter SW, Levine B (2013) Autophagy in Human Health and Disease. N Engl J Med 368(7): 651- 662.
-
Winiarska A, Knysak M, Nabrdalik K, Gumprecht J, Stompór T (2021) Review Inflammation and Oxidative Stress in Diabetic Kidney Disease: The Targets for SGLT2 Inhibitors and GLP-1 Receptor Agonists. Int J Mol Sci 22(19): 10822.
-
Huang JS, Chuang CT, Liu MH, Lin S-H, Guh J-Y, et al. (2014) Klotho attenuates high glucose-induced fibronectin and cell hypertrophy via the ERK1/2-p38 kinase signaling pathway in renal interstitialfibroblasts. Mol Cell Endocrinol 390(1-2): 45-53.
-
Nie F, Wu D, Du H, Yang X, Yang M, et al. (2017) Serum klotho protein levels and their correlations with the progression of type 2 diabetes mellitus. J Diabetes Complications 31(3): 594-598.
-
Corcillo A, Fountoulakis N, Sohal A, Farrow F, Ayis S, et al. (2020) Low levels of circulating anti-ageing hormone Klotho predict the onset and progression of diabetic retinopathy. Diabetes & Vascular Disease Research 17(6).
-
Bapputty R, Talahalli R, Zarini S, Samuels I, Murphy R, et al. (2019) Montelukast Prevents Early Diabetic Retinopathy in Mice Reena Diabetes 68(10): 2004-2015.
-
Chen K, Sun Z (2019) Autophagy plays a critical role in Klotho gene deficiency-induced arterial stiffening and hypertension. Journal of Molecular Medicine 97: 1615- 1625.
-
Dahan I, Thawho N, Farber E, Nakhoul N, Asleh R, et al. (2018) The Iron-Klotho-VDR Axis Is a Major Determinant of Proximal Convoluted Tubule Injury in Haptoglobin 2-2 Genotype Diabetic Nephropathy Patients and Mice. J Diabetes Res 2018: 7163652.
-
Yamamoto M, Clark JD, Pastor JV, Gurnani P, Nandi A, et al. (20005) Regulation of oxidative stress by the anti-aging hormone klotho. J Biol Chem 280(45): 38029-38034.
-
Eshaq RS, Aldalati AMZ, Alexander JS, Harris NR (2017) Diabetic retinopathy: breaking the barrier. Pathophysiology 24(4): 229-241.
-
Di Rosaa M, Distefanoa G, Gaglianob C, Rusciano D, Malaguarnera L (2016) Autophagy in Diabetic Retinopathy. Current Neuropharmacology 14(8): 810- 825.
- Investigation of Polymorphisms in PPAR-Ɣ and TRHR Genes and their Impact on Turkish Diabetic and Obese Individuals
- The Impact of Aircraft Noise Exposure on the Efficacy of Empagliflozin Therapy in an Animal Model of Obesity
- Rooibos Mitigates Metabolic and Inflammatory Dysfunctions in Mice Fed a High-Carbohydrate Diet
- Synergistic Effect of Combined Leaf Extract of Vernonia amygdalina, Ocimum gratissimum, and Zingiber officinale Tuber on Phytochemical Profile, Antioxidant Activity, Serum Insulin, and Biochemical Parameters in Streptozotocin-Induced Diabetic Rats
- Investigation of Cardiovascular Responses to Aerobic Exercise in Obese University Students
- A Look at the Phase Angle Obtained by Electrical Bioimpedance