Predictive Modelling of Peroxidase, Polyphenol Oxidase and Catalase in Post-Harvest White Trifoliate (D. Dumetorum Var.) Yam Tubers
The enzymes peroxidase (EC 1.11.1.7), polyphenol oxidase (EC 1.10.3.1) and catalase (EC 1.11.1.6) are fundamental in the development of Dioscorea dumetorum yam tissues. Enzymological studies on activities of peroxidase, polyphenol oxidase and catalase in white trifoliate yam tubers stored for three (3) days under room temperature (28±2˚C) and 72-85%RH, and at refrigeration (5±1˚C) and 85-100%RH conditions were fitted in a second order polynomial model. The model showed that at day three (3) of yam storage, a 750units activity of peroxidase relates to 1200units of polyphenol oxidase activity in white trifoliate yam tubers stored at 28±2˚C and 72-85%RH. Similarly, day three (3) of yam storage, a 7.5 units of catalase activity relates to 1000units of peroxidase activity in white trifoliate yam tubers stored at 28±2˚C and 72-85%RH. Also, day three (3) of yam storage, a catalase activity of 2 units corresponded to 400 units for both peroxidase and polyphenol oxidase activities in white trifoliate yam tubers at 5±1˚C and 85-100%RH. This data describes the mechanism of association of these enzymes as may be linked to the production of biomolecules that affect biochemical changes and physical properties in post-harvest white trifoliate yam tuber.
Bernard TN O*, George SA and Firibu KS
Nutrition and Food science Department, University of Ghana, Box LG 134 Legon, Ghana *Corresponding author: Bernard TN Otubuah, Nutrition and Food science Department, University of Ghana, Box LG 134 Legon, Ghana, Tel: +233(0)267642008; Email: botubuah@ gmail.com
Introduction
Large amount of trifoliate yam (D. dumetorum) tubers can be produced in the yam zones of the world. However, these tubers have demonstrated post-harvest tissue hardening in the most dramatic manner [1, 2, 3]. In stored trifoliate yam tubers, physical changes of the tubers are attributed to lignification [1, 3, 4, 5]. Peroxidases and polyphenol oxidases are known regulators of lignin biosynthesis in vegetables [6, 7, 8]. Besides, peroxidases and catalases regulate the availability of hydrogen peroxide (H2O2) in the cell wall, which serves as the basis for cell wall rigidity during stress [9, 10]. This suggests that lignification of the cell wall could be as a result of collective roles of enzymes such as peroxidase, polyphenol oxidase and catalase. Recently, activities of peroxidase, polyphenol oxidase and catalase were shown to be in phase with dramatic tissue hardening in white trifoliate yam tuber at ambient conditions [6]. Again, lignified parenchyma cell has been shown to be the cause of dramatic tissue cohesion and hardening in post-harvest trifoliate yam tubers [3]. However, how these enzymes relate during stress in bringing about the sudden changes in chemical and physical properties in white trifoliate yam tuber are unknown. Therefore, modelling to fit a second order polynomial of these enzymes shall highlight their associations as linear, square and interactive. As a result, the purpose of this study is to model and predict how enzymes such as peroxidase, polyphenol oxidase and catalase act together at post-harvest of stored white D. dumetorum yam tubers.
Materials and Methods
Plant Materials
Wholesome, mature D. dumetorum yam tubers (characterized by dry vines and withered leaves), (white cultivar) were simultaneously harvested in early/January (2013), after 11 months of planting from an Experimental farm (Plant Genetic Resource Research Institute, Bonsu) in Ghana [2]. Care was taken during harvesting, cleaning and transportation not to injure the tubers [2]. Harvesting of the tubers was done between the hours of 8.00am- 9:30 am [2]. During harvesting, soil temperatures of the yam mounds were estimated to be 22 ± 2°C and with a corresponding dry season RH (40%) [2]. Samples were randomly selected and cleaned, kept cold in an ice chest containing ice gels [2]. They were transported (within 3hrs) to the laboratories [2]. The samples were divided into the following treatments: (i) stored at room temperature (28 ± 2°C, 72-85% RH) and (ii) under refrigeration conditions (5 ± 1°C, 85-100% RH) for a period of 0, 1, 2 and 3 days [2]. Day of harvest and arrival at the laboratory was taken as day zero [2].
Enzyme Extraction and Assay
The enzymes in the tuber were extracted from three randomly selected tubers of each cultivar [2]. Samples (2g) were homogenized with 10 ml of cold phosphate buffer pH 6.8 (0.2 M) in prechilled mortar and pestle; and were centrifuged at 4°C for 15 min at 15,000rpm [2]. The clear supernatant were taken as the enzyme source and kept frozen [2].
Peroxidase Assay: The peroxidase activity was done following the method Kar M, et al. [11] with the following modifications Otubuah BTN, et al. [2] in a final volume of 5 ml of the assay mixture comprised: 1ml of 12.5 mM phosphate buffer, 2ml of 5mM pyrogallol, 1ml of 5mM H2O2, and 1 ml of the diluted enzyme extract (1ml of enzyme stock dissolved in 9 ml phosphate buffer pH 6.8 (0.2M)). This was incubated for 5 min at 25˚C after which the reaction was stopped by adding 0.5 ml of 5% (v/v) H2SO4. The amount of purpurogallin formed was determined by taking the absorbance at 420nm using a 1cm path-length cuvette in a spectrophotometer (Cecil Instruments, Cambridge- England, C≡ 3041). To the blank, the phosphate buffer pH 6.8 (0.2M) solution was added as the enzyme solution. The specific activity of peroxidase (nmoles purpurogallin/mg protein/min) was calculated using the extinction coefficient [2.47 mM-1cm-1] for purpurogallin [12].
Polyphenol Oxidase Assay: The polyphenol oxidase activity was done following the method Boerjan W, et al. [11] with the following modifications Otubuah BTN, et al. [2] in a final volume of 5 ml of the assay mixture comprised: 1ml of 12.5 mM phosphate buffer, 3ml of 5mM pyrogallol and 1ml of enzyme stock dissolved in 9 ml phosphate buffer pH 6.8 (0.2M). This was incubated for 5 min at 25°C after which the reaction was stopped by adding 0.5 ml of 5% (v/v) H2SO4. The absorbance of the purpurogallin formed was taken at 420 nm using a 1cm path-length cuvette in a spectrophotometer (Cecil Instruments, Cambridge- England, C≡ 3041). To the blank, the phosphate buffer pH 6.8 (0.2M) solution was added as the enzyme solution. The specific activity of polyphenol oxidase (nmoles purpurogallin/mg protein/min) was calculated using the extinction coefficient [2.47 mM-1cm-1] for purpurogallin [12].
Catalase Assay: Catalase activity was assayed by the method Sinha AK [13] with the following modifications [2]. The diluted enzyme (1ml) extract (1ml of enzyme stock dissolved in 9 ml phosphate buffer pH 6.8 (0.2M)) was added to the reaction mixture containing 1ml of 0.1 M phosphate buffer (pH 7.0), 4 ml of 5mM H2O2 and then incubated for 5 minutes at 25°C. The reaction was terminated by the addition of 0.5 ml of 5% (v/v) H2SO4 and the absorbance was read at 610 nm using a 1cm path-length cuvette in a spectrophotometer (Cecil Instruments, Cambridge- England, C≡ 3041). To the blank, the phosphate buffer pH 6.8 (0.2M) solution was added as the enzyme solution. Specific catalase activity was expressed in terms of nmoles H2O2 consumed/mg protein/ min using the extinction coefficient [40 mM -1cm-1] for H2O2 [11].
Protein Assay: The extraction of soluble proteins was followed as described Zhou B, et al. [14] with modifications described by Otubuah BTN, et al. [2]. Fresh sample of white cultivar (2g) was homogenized in 10 ml of cold phosphate buffer pH 6.8 (0.2M) and centrifuged at 15000 rpm for 15 min at 4°C. Protein concentrations were determined by a modified method using bovine serum albumin (BSA) as a standard [15]. 1ml of the extract (10 times diluted with buffer solution), 5ml of alkaline reagent (made of 48ml of reagent A (20g NaCO3 in 1L of 0.1M NaOH) and 2ml of reagent B (5g CuSO4.5H2O in 1L of 10g/L Na, K-tartrate) was added and properly vortexed. This was left to stand for 10 minutes. Folin-Ciocalteu reagent (0.5ml) diluted with distilled water (1:3) was added and rapidly mixed. The absorbance was read after 30 minutes (at room temperature) at 750nm. The calibration equation for BSA was y= 0.172x + 0.1655 (R2= 0.998). Proteins were expressed as mg BSA equivalent/mg dry weight.
Statistical Analysis
Response surface regression analysis was used to model the enzyme data to fit a second order polynomial (Y = b0+b1 x1+b2x2+b12x1x2+b11x2 1+b22x2 2) where b1, b2, b11, b22 and b12 are coefficients for the square, interactive and linear terms of the equation. The dependent and independent variables of the second order polynomial was Y, x1 and x2. Also, a significant difference was set at p ≤ 0.05. R2 and lack of fit was used to evaluate the adequacy of the models. All data analyses were done using Minitab (version 15) and Microsoft Office Excel (2010).
Results and Discussion
Relating Enzyme Activities Such as Peroxidase, Catalase and Polyphenol Oxidase in White Trifoliate Yam at 28±2°C and 72-85%RH
From the study, specific activity of polyphenol oxidase increased to about 400 units in day two (2) of storage when peroxidase activity was lower than 250 units (Figure A). Again, an increase in polyphenol oxidase activity was observed at 750 units of peroxidase activity (Figure 1A). These imply, increasing peroxidase activity above 250 units competitively activated polyphenol oxidase activity during the storage period. However, main effect of storage duration on specific polyphenol oxidase activity was high compared to peroxidase activity. An interactive effect of peroxidase activity was observed to act complementarily on polyphenol oxidase activity (Figure 1A) (Table 1). This suggests that, long term effect of peroxidase activity dramatically increases polyphenol oxidase activity in white trifoliate yam at room temperature. This explains the observation that lignification has a faster rate in stored trifoliate yam tubers [4]. Again, activities of enzymes were shown to be in phase with dramatic tissue hardening in white trifoliate yam tuber at ambient conditions [17]. Moreover, peroxidase and polyphenol oxidase in strawberry cultivars ‘Elsanta and Madame Moutot’ were shown to be moderately correlated (0.64 and 0.76) [16]. Also, the study showed that immediately after harvest, a unit activity of catalase increased peroxidase activity by 89 folds. Similarly, a unit activity of catalase increased polyphenol oxidase activity 62 folds. But, peroxidase activity was 0.7 faster than polyphenol oxidase activity in freshly harvested tubers. These observations were more dramatic in tubers stored for three (3) days at ambient conditions where a unit activity of catalase increased peroxidase activity by 122 folds. Again, a unit activity of catalase increased polyphenol oxidase activity by 147 folds. Once more, a 1.2 fold increase in polyphenol oxidase activity was observed to be linked to a unit peroxidase activity in three (3) days stored trifoliate tubers at ambient condition. This finding confirms report suggesting that peroxidase responded earlier to abiotic shock than polyphenol oxidase as related to tissue hardness [2].
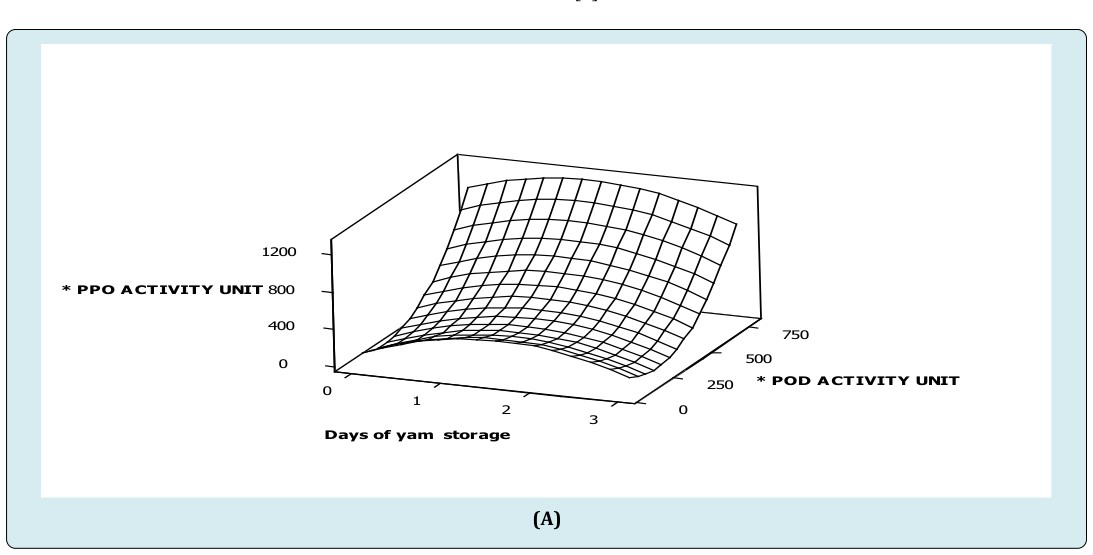
(C) Figure 1: Surface plot of (A) specific polyphenol oxidase activity versus specific peroxidase activity, storage duration of white trifoliate yam stored at room temperature (B) specific peroxidase activity versus specific catalase activity, storage duration of white trifoliate yam stored at room temperature (C) specific polyphenol oxidase activity versus specific catalase activity, storage duration of white trifoliate yam stored at room temperature. *Specific activity of peroxidase (nmoles purpurogallin produced/mg protein/min) *Specific activity of polyphenol oxidase (nmoles purpurogallin produced/mg protein/min) *Specific activity of catalase (nmoles H2O2 consumed/mg protein/min)
| Regression Coefficients | PPO4 Activity | PPO5 Activity | POD6 Activity |
|---|---|---|---|
| B : constant O | -37.707 | -130.052 | -147.1 |
| B : time A | -103.457* | 121.959 | 254.27* |
| B : CAT Activity B | (-) | 281.461 | 370.67 |
| B : POD Activity C | 1.365* | (-) | (-) |
| B : PPO Activity D | (-) | (-) | (-) |
| B AA | 13.31 | -8.758 | -66.61* |
| B AB | (-) | -5.982 | 14.42 |
| B AC | 0.543 | (-) | (-) |
| B AD | (-) | (-) | (-) |
| B BB | (-) | -79.88 | -118.22 |
| B CC | -0.003* | (-) | (-) |
| B DD | (-) | (-) | (-) |
| R2 (%) | 97.57 | 85.93 | 63.88 |
| Lack of fit (p) | 0.095 | 0 | 0.117 |
Table 1: Regression coefficients, coefficient of determination (R2), Lack of fit and analysis of variance of regression models fi
Table 1: Regression coefficients, coefficient of determination (R2), Lack of fit and analysis of variance of regression models fitted for enzymatic profile of stored white trifoliate yam at refrigeration using response surface plot. *Significant at p≤ 0.05. (-) means such parameter was not included in the model for predicting the activation and inhibition of enzymes as enzyme activity increases. PPO4 was based on peroxidase (POD) activity (Figure 1A), PPO5 was based on catalase (CAT) activity (Figure 1B) and POD6 was based on catalase (CAT) activity (Figure 2C).
Furthermore, interactive effects of storage duration by peroxidase activity influenced polyphenol oxidase activity negatively (Figure 1A, Table 1), implying that at a particular storage duration and peroxidase activity, polyphenol oxidase activity was hindered slightly. Additionally, the highest specific peroxidase activity also corresponded to the highest specific activity of catalase in day three (3) of storage, suggesting that peroxidase and catalase activities competitively act together (Figure 1B). This explains why catalase and peroxidase competed for hydrogen peroxide when plants were stressed [17]. Subsequently, the coefficients of the main effects of catalase activity and storage duration revealed that storage duration influences peroxidase more than catalase activity (Figure 1B) (Table 2). In addition, interactive effects of storage duration by catalase activity acted positively on peroxidase activity in white trifoliate yam stored at room temperature storage (28±2°C). It also been observed that the specific activity of polyphenol oxidase increases as storage duration increases at lower catalase activity (≤ 2.5unit) suggesting competitive increase in polyphenol oxidase activity when catalase activity was insignificant (≤ 2.5unit). However, as catalase activity (≥ 2.5unit) increased in day three (3) of storage, polyphenol oxidase activity decreased gradually in white trifoliate yam stored at room temperature (Figure 1C). Subsequently, coefficients of the main effects of catalase activity and storage duration revealed that storage duration influences polyphenol oxidase activity more than catalase activity (Figure 1C, Table 1). In addition, interactive effect of time (storage duration) by catalase activity acted positively on polyphenol oxidase activity in white trifoliate yam stored at room temperature (28±2°C); implying that at a particular storage duration and catalase activity, polyphenol oxidase activity might either increase steadily or remain unchanged (Figure 1C, Table 1). The sharp increases in enzymic activities such as peroxidase, polyphenol oxidase and catalase due to their working together as demonstrated suggest correspondingly high increase in synthetic potential for the production of biomolecules that can affect biochemical changes and physical properties of white D. dumetorum yam tissues shortly after harvest
| Regression Coefficients | POD1 Activity | PPO2 Activity | PPO3 Activity |
|---|---|---|---|
| B : constant O | 70.113* | 88.16 | 154.7 |
| B : time A | 46.058 | 252.83* | 366.6* |
| B : CAT Activity B | 38.013 | -38.1 | (-) |
| B : POD Activity C | (-) | (-) | -1.321 |
| B : PPO Activity D | (-) | (-) | (-) |
| B AA | 3.41 | 196.46* | -116.98 |
| B AB | 52.235* | -100.15 | (-) |
| B AC | (-) | (-) | -0.057 |
| B AD | (-) | (-) | (-) |
| B BB | -17.163* | 12.15 | (-) |
| B CC | (-) | (-) | 0.003 |
| B DD | (-) | (-) | (-) |
| R2 (%) | 99.83 | 99.56 | 99.45 |
| Lack of fit (p) | 0.014 | 0.399 | 0.542 |
Table 2: Regression coefficients, coefficient of determination (R2), Lack of fit and analysis of variance of regression models fi
Table 2: Regression coefficients, coefficient of determination (R2), Lack of fit and analysis of variance of regression models fitted for enzymatic profile of stored white trifoliate yam at room temperature using response surface plot. *Significant at p≤ 0.05. (-) means such parameter was not included in the model for predicting the activation and inhibition of enzymes as enzyme activity increases. POD1 was based on catalase (CAT) activity (Figure 1A), PPO2 was based on catalase (CAT) activity (Figure 1B) and PPO3 was based on peroxidase (POD) activity (Figure 1C)
Relating Enzyme Activities Such as Peroxidase, Catalase and Polyphenol Oxidase in White Trifoliate Yam at 5±1°C and 85-100%RH
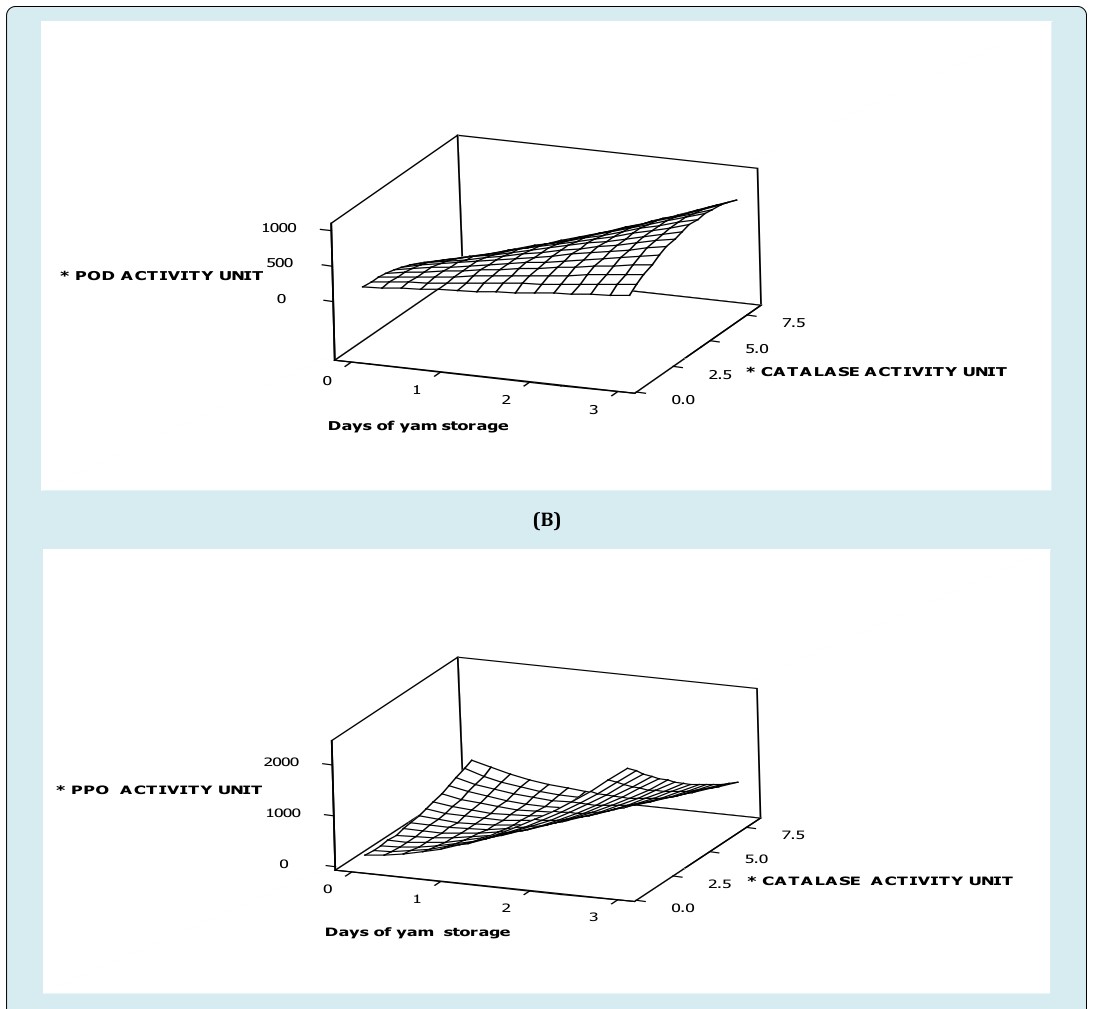
Once more, when white trifoliate yam was stored at 5±1°C and 85-100%RH, specific activity of polyphenol oxidase decreased gradually to about 90 units at day three (3) of storage when peroxidase activity was below 100units (Figure 2). Conversely, polyphenol oxidase activity was observed to rise sharply during storage period (day 0-3) when peroxidase activity increased from 100 units to 400 units (Figure 2A). However, main effect of storage duration on specific polyphenol oxidase activity was low compared peroxidase activity. Besides, these effects on polyphenol oxidase activity were significant (p≤0.05). Again, interactive effect of peroxidase activity significantly (p≤0.05) acted negatively on polyphenol oxidase activity during refrigeration in white trifoliate yam (Figure 2A, Table 2), suggesting that long term effect of peroxidase activity less than 100 units gradually reduces polyphenol oxidase activity in refrigerated white trifoliate yam. On the contrary, interactive effects of storage duration by peroxidase activity influenced polyphenol oxidase activity positively (Figure 2A, Table 2); implying that at a particular storage duration and peroxidase activity, polyphenol oxidase activity might be increased steadily. Hence, the link between peroxidase and polyphenol oxidase could be argued that peroxidases produce hydrogen peroxide or use hydrogen peroxide as a co-substrate for which the end product of diaoxygen is also a co-substrate for polyphenol oxidase and this explains the biochemical basis for the enzymes acting together during storage. Similarly, the biochemical bases for competiveness of peroxidase and catalase are due to hydrogen peroxide bioavailability [10, 17, 18]. Furthermore, the potency of hydrogen peroxide formation and breakdown by peroxidases and the subsequent hydrogen peroxide breakdown by catalases influences diaoxygen gradient which directly determines polyphenol oxidase activities, and indirectly determines catalase and peroxidase activities respectively. Again, specific polyphenol oxidase activity increases gently during refrigeration period (0-3 days) as catalase activity increases (0-2 units) simultaneously, implying that activities of catalase and polyphenol oxidase are competitively increased (Figure 2B). On the contrary, specific polyphenol oxidase activity had increased in day two (2) then it decreased gradually in day three (3) (Figure 2C). Subsequently, the coefficients of the main effects of catalase activity and storage duration revealed that storage duration influences peroxidase more than catalase activity (Figure 2B) (Table 2). In addition, interactive effects of storage duration and storage duration by catalase activity acted negatively on polyphenol oxidase activity in white trifoliate yam stored at refrigeration storage (4°C) (Table 2). On the other hand, it was evident that specific peroxidase activity increases gradually up to day two (2) to about 120 units then decreases gently to about 30 units in day three (3) when catalase activity was low (Figure 2C). Furthermore, maximum peroxidase activity was observed at day two (2) when catalase activity was at a maximum (2 units), implying that activities of catalase and peroxidase act together at a slower phase at refrigeration. Again, least peroxidase activity was observed in freshly harvested white trifoliate yam when catalase activity was low (Figure 2C). These observation explain why a decrease in enzymes’ activities particularly peroxidase and catalase in white trifoliate yam tubers at refrigeration (5±1˚C) and 85-100%RH was comparable to no changes in tissue hardness, indicating that very low enzyme profiles corresponded to no increase in tissue hardness (2). But, coefficients of the main effects of catalase activity and storage duration revealed that storage duration influences peroxidase more than catalase activity (Figure 2B, Table 2). Therefore, interactive effect of storage duration and catalase activity acted negatively on peroxidase activity in white trifoliate yam stored at refrigeration (4°C) (Figure 2B). The combined effect of the enzymes as discussed explains the prolonged dramatic hardening of the yam tissues at room temperatures than under refrigeration conditions.
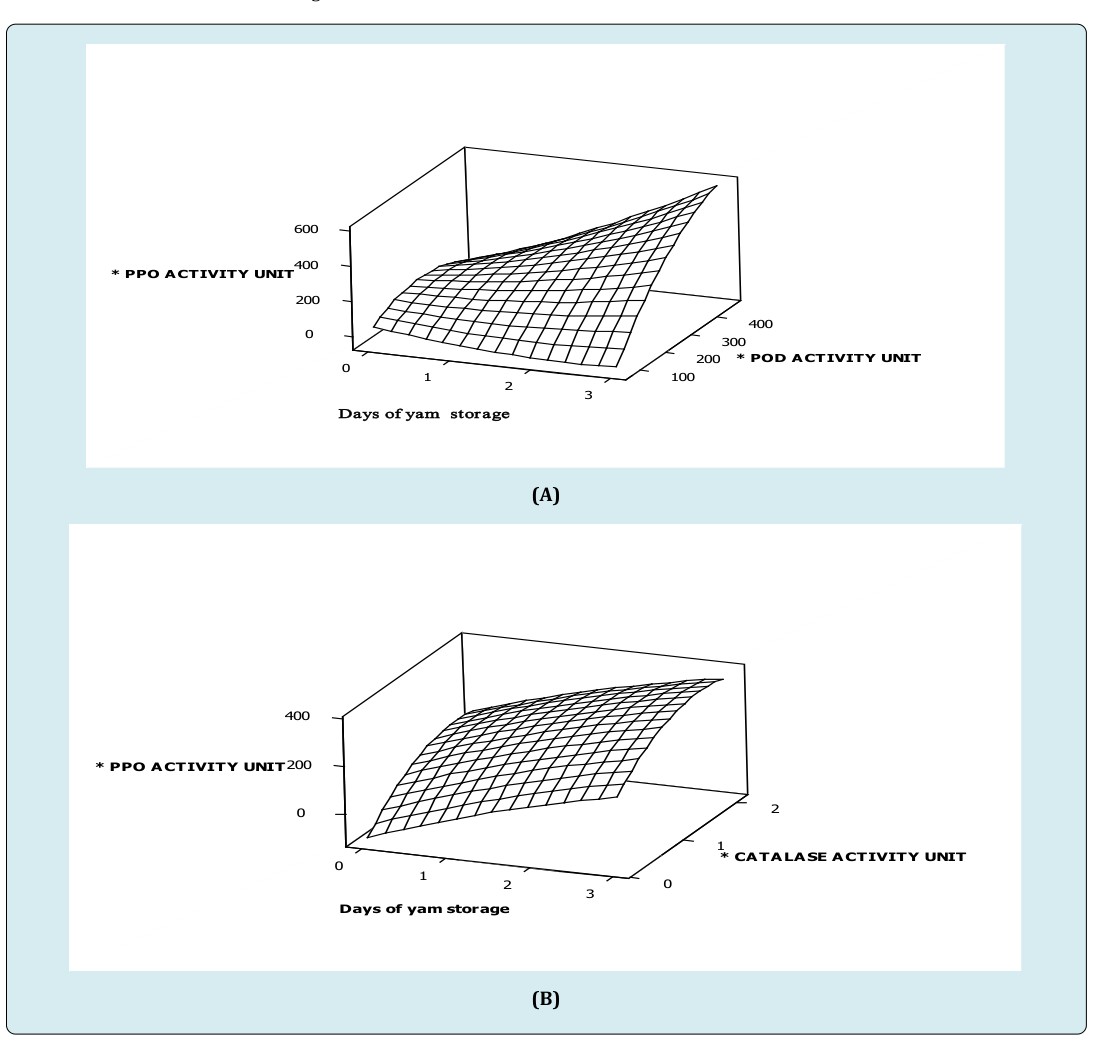
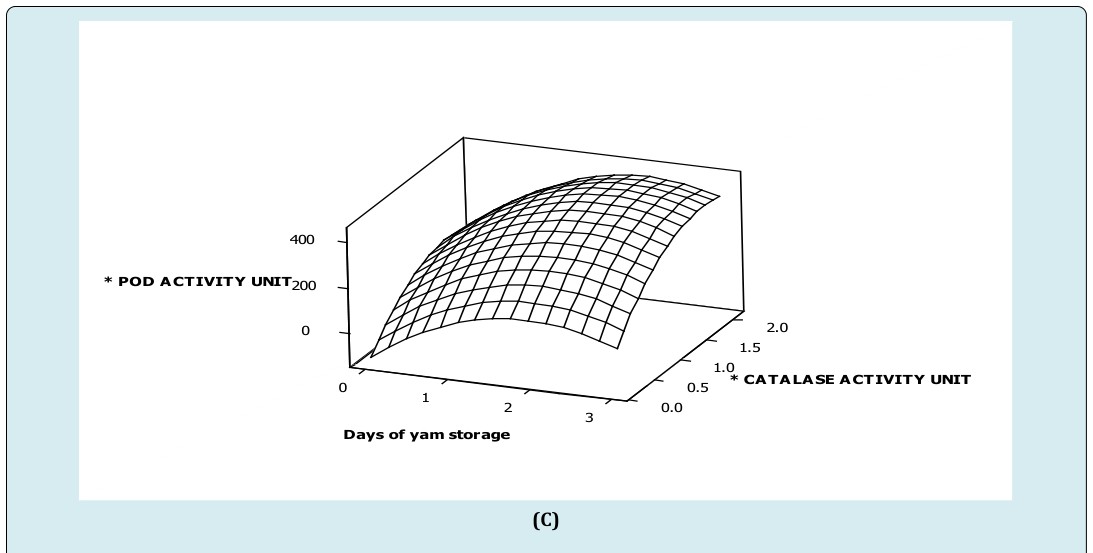
Figure 2: Surface plot of (A) specific polyphenol oxidase activity versus specific peroxidase activity, time of white trifoliate yam stored at refrigeration temperature (B) specific polyphenol oxidase activity versus specific catalase activity, time of white trifoliate yam stored at refrigeration temperature (C) specific peroxidase oxidase activity versus specific catalase activity, time of white trifoliate yam stored at refrigeration temperature. *Specific activity of catalase (nmoles H2O2 consumed/mg protein/min) *Specific activity of peroxidase (nmoles purpurogallin produced/mg protein/min) *Specific activity of polyphenol oxidase (nmoles purpurogallin produced/mg protein/min) In summary, activities of enzymes such as peroxidase, polyphenol oxidase and catalase as storage duration of trifoliate yam was prolonged, appeared to be in phase with one another suggesting dramatic working together of these enzymes at post-harvest storage of white trifoliate yam tubers at 28±2°C and 72-85%RH. This meant that as storage duration of the yam tuber was increased from day zero (0) to three (3), activity profiles of the enzymes’ such as peroxidase, polyphenol oxidase and catalase were related such that an increasing activity profile of an enzyme such as catalase can competitively initiate an increase in activity profiles of other enzymes such as peroxidase and polyphenol oxidase whiles peroxidase activities can competitively activate a dramatic increase in activity profile of polyphenol oxidase. Again, similar observations were made under refrigeration conditions but were not dramatic compared to the enzymes at ambient conditions. This explains the dramatic working together or otherwise of these enzymes at post-harvest storage as may be related to chemical and physical changes in white trifoliate yam tissues.
Acknowledgements
The authors are thankful to Mr. Owusu Danso and Mr. K. Danso (Plant Genetic Resource Research Institute, Bonsu- Ghana) for providing the trifoliate yam tubers (white) for the research work. Thanks to Mr. Daniel Tetteh Otu Amanquah for providing me with an accommodation prior to harvesting. I also thank Mr. Nelson Tsigbe for assisting with the harvesting and transportation of the tubers to the laboratory. Special thanks to Ms. Flora Ugbekile, Mrs. Joyce Duah, Mrs. Leonora Wontumi (Nutrition and Food science Department, University of Ghana) for their numerous supports to this work. This work is an excerpt from Bernard T. N. Otubuah MPhil thesis (Enzymology of Post-Harvest Hardening of Dioscorea dumetorum yam tuber during storage), supervised by Prof. G. S. Ayernor and Prof. F. K. Saalia.
Conflict of Interest
The authors declare no conflict of interest.
References
-
Brillouet JM, Treche S, Sealy L (1981) Alterations in cell wall constituents of yams _Dioscorea dumetorum_ and _Dioscorea rotundata_ with maturation and storage conditions-Relation with post- harvest hardening of _Dioscorea dumetorum_ yam tubers. Journal of food science 46(6): 1964-1967.
-
Otubuah BTN, Ayernor GS, Saalia FK (2015) Enzymic Basis of Sharp Tissue Hardening in Post-Harvest White Trifoliate (_D_. _dumetorum_ var.) Yam Tubers. American Journal of Food Science and Nutrition Research 2(3): 98- 107.
-
Otubuah BTN, Ayernor GS, Saalia FK (2015) Histochemical Evidence of Dramatic Tissue Hardening in Post-harvest Trifoliate (D. dumetorum vars.) Yam Tubers. American Journal of Food and Nutrition 3(3): 75-83.
-
Medoua GN, Mbofung CMF (2006) Hard-to-cook defect in trifoliate yam _Dioscorea dumetorum_ tubers after harvest. Food Research International 39(5): 513-518.
-
Trèche S, Agbor-Egbe T (1996) Biochemical changes occurring during growth and storage of two yam species. International Journal of Food Science and Nutrition 47(2): 93-102.
-
Boerjan W, Ralph J, Baucher M (2003) Lignin Biosynthesis. Annul Rev Plant Biol 54: 519-46.
-
Mayer AM, Staples RC (2002) Laccas: new functions for old enzyme. Phytochemistry 60: 551-565.
-
Vance CP, Kirk TK, Sherwood RT (1980) Lignification as a mechanism of disease resistance. Ann Rev Phytopothol 18: 259-88.
-
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants, Oxidative damage and oxygen deprivation stress: a Review. Ann Bot 91(2): 179-194.
-
Passardi F, Penel C, Dunand C (2004) Performing the paradoxical: how plant peroxidases modify the cell wall. Trends in Plant Science (Review) 9 (11): 534-540.
-
Kar M, Mishra D (1976) Catalase, peroxidase, and polyphenol oxidase activities during rice leaf senescence. Plant Physiol 57: 315-319.
-
Haddadchi GR, Gerivani Z (2009) Effects of Phenolic Extracts of Canola (_Brassica napuse_ L.) on Germination and Physiological Responses of Soybean (_Glycin max_ L.) Seedlings. International Journal of Plant Production 3(1): 63-74.
-
Sinha AK (1972) Colorimetric assay of catalase. Anal Biochem 47(2): 389-394.
-
Zhou B, Chen Z, Du L, Ye X, Li N (2012) Correlation between Resistance of Eggplant and Defense-Related Enzymes and Biochemical Substances of Leaves. African Journal of Biotechnology 11(74): 13896-1390.
-
Plummer DT (1987) An introduction to practical biochemistry. 3rd (Edn.), McGraw-Hill Book Company, London, pp: 159.
-
Chisari M, Barbagallo RN, Spagna G (2007) Characterization of Polyphenol Oxidase and Peroxidase and influence of browning on cold stored strawberry fruit. J Agric Food Chem 55(9): 3468-3476.
-
Rodrigo C, Rodrigo M, Alvarruiz A, Frigola A (1996) Thermal inactivation at high temperatures and regeneration of green asparagus peroxidase. J Food Prot 59: 1065-1071.
-
Tayefi-Nasrabadi H, Dehghan G, Daeihassani B, Movafegi A, Samadi A (2001) Some biochemical properties of catalase from safflower (_Carthamus tinctorius_ L. cv. M-CC-190). African Journal of Agricultural Research 6(23): 5221-5226.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends