Nutriepigenomics: Need of the Day to Integrate Genetics, Epigenetics and Environment towards Nutritious Food for Healthy Life
Nutrition, lifestyle, behavior, and other environmental factors affect human health that may control pathogenesis of various diseases, and it is regulated through epigenetic modifications. Nutrition is an indispensable environmental factor that not only impacts our growth and development but also the health by affecting epigenetic phenomena. Furthermore, these changes might impact next generation via transgenerational inheritance of epigenetic alterations; thus, making them important health issues for the society. Certain nutrients and bioactive food components are able to alter epigenetic status at DNA methylation and histone modulation levels, which might modulate expression of the genes associated with patho-physiological processes. In addition to the genetics, health condition of an organism might be affected by the integration of environmental factors/signals acting through epigenetic changes. Therefore, understanding the molecular (epigenetic) basis of environmental factors has become a need of the day towards development of preventative strategies against various pathological conditions like obesity, type II diabetes, cardiovascular diseases, cancer etc. utilizing a personalized dietary chart.
Introduction
Unravelling the codes of about 2.9 billion nucleotides or nearly 30000 genes has provided tremendous opportunities to understand response of human body to the dietary components, other environmental factors, and how these factors affect human health. Apart from metabolic production of energy, food is now also known to provide conditioning environment that shapes the activities of the genome and physiology of the body [1]. A deeper understanding of how nutrients influence genes and their expression might help managing healthy life in a better way. Explaining the genetic variations and the continuously changing genotype due to the environmental factors, including dietary/nutritional components, using classical genetics alone has become arduous [2]. Moreover, the rate of genetic mutations and phenotypic variations are considerably different from each other which cannot be explained merely based on genetics [3]. Different tissues in an organism have different phenotype/physiology regardless of possessing the same genome, which is mainly because of differential expression of the genes regulated by the pliable epigenetic landscape of an individual (Figure 1). A better understanding of the epigenetic switching on/off gene might help explaining this enigma [4].
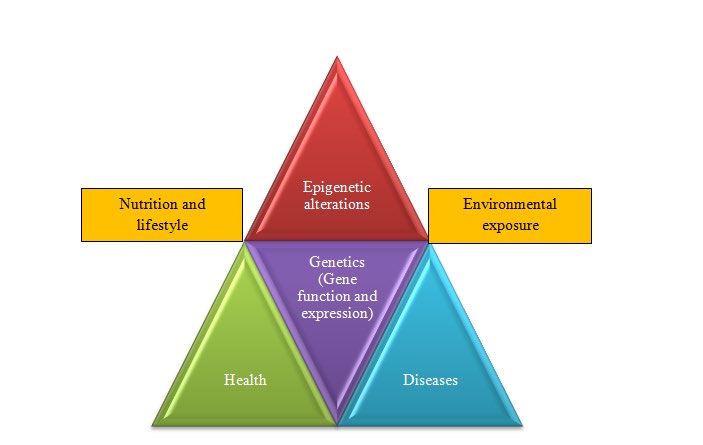
If epigenetics is contemplated as a harmonizing process with genetics, many of the phenotypic variations can be easily explained [5]. Epigenome refers to the sum total of all the changes in genetic material without altering the underlying nucleotide sequence. Studies of such changes including modification in chromatin architecture, histone protein, DNA methylation and non-coding RNA biogenesis that affect gene expression in a cell/tissue is known as epigenomics [6]. The epigenetic changes might revert back to the previous state after normalization of the environmental conditions. One of the well-known mechanisms of epigenetic change has been methylation of cytosine at the 5th carbon [5]. In addition, certain amino acids located at the tail of the histone proteins are post-translationally modified by methylation, acetylation or biotinylation [7]. Similarly, non-coding RNAs also play very important role in epigenetic modifications and gene regulation. More importantly, some of the epigenetic changes may be inherited to the next generation that might result in phenotypic variations [8]. It is becoming apparent that changes in epigenome of living organism play crucial role in tolerance to environmental stresses, nutrient availability, and the evolution processes [9].
The on-going researches on nutritional epigenetics justify the direct association between dietary nutrients and epigenetic alterations. Various nutrients possess the ability to alter the epigenetic pattern via availability of the substrate and activities of the related enzymes. In the field of nutritional science and technology, epigenetic changes have gained exceptional importance because of the ability of nutrients and bioactive food components to change the epigenetic status of gene/genome which can alter the gene expression [10]. Bioactive food compounds like folic acid, vitamin-B12, methionine and choline might affect epigenetic modifications through one-carbon metabolism [11, 12]. In principle, any nutrient/bioactive compound or condition that can affect AdoMet (S-adenosylmethionine) or AdoHcy (S-adenosylhomocysteine) level in a tissue can alter methylation of DNA/histone proteins [13]. Different vitamins play important roles in histone modification, and affect the enzymes involved in the epigenetic mechanisms [14]. The roles of epigenetic alterations in certain patho-physiological processes are summarized in Table 1.
| Dietary components | Patho- physiological state | Epigenetic modification | References |
|---|---|---|---|
| Butyrate | Stem cells | Histone acetylation, DNA | [15] |
| Methylation | |||
| Folate | Embryonic growth and development | DNA methylation, imprinting | [16,17] |
| Choline | DNA methylation | [18] | |
| Alcohol | DNA methylation | [19] | |
| Choline | Neurocognition | DNA methylation | [18] |
| Histone methylation | [20] | ||
| Fat rich diet | Obesity and insulin resistance | DNA methylation, miRNA biogenesis, Histone acetylation | [21] |
| Curcumin | |||
| Resveratrol | Inflammation | Histone acetylation | [22] |
| Methyl-deficient diet | MicroRNAs | [23] | |
| Folate | Immune system functioning | DNA methylation | [24] |
| Calorie restriction | Aging | DNA methylation | [24] |
| Histone acetylation | [25] |
Table 1: Dietary components associated with patho-physiological conditions and epigenetic modifications.
The present review discusses the influence of environmental factors (including the food/dietary components) on epigenetic behavior of living organism, and their effects on human health and diseases, and the future prospective of an emerging branch of science namely nutriepigenomics.
Nutrition and Genomics: Nutrigenomics
Nutritional genomics (nutrigenomics) deals with the identification, classification, and characterization of genetic variations in the genome that modulate metabolism of various nutrients present in the food/diet. Thus, nutrigenomics focuses on understanding the pathways that are modulated by gene-nutrient interaction for the management of patho- physiological conditions and healthy life. It is now well- understood that chronic diseases are quite complex in their etiology, and comprise of a substantial genetic component as every individual has a different response to food and its nutrients [26, 27]. This clearly indicates that the linkage between genes and environment is bidirectional, and the environment including food and nutrients can indirectly affect gene expression/response of the organism.
Role of Dietary Components in Genome Stabilization and Gene Expression
Different components of a diet may affect expression of genes and stability of the genome. For example, loss of amino acids in the extra-cellular environment enhances expression of asparagine synthetase (AS) gene [28]. AS encodes the enzyme that catalyses the reaction responsible for conversion of aspartic acid to asparagine. The diets rich in carbohydrates, primarily glucose, induce transcription of some of the glycolysis and lipogenic pathway genes [29]. It is also known that carbohydrate-rich diets are linked to patho- physiological conditions like obesity, type II diabetes and hyperlipidemia. Vitamins play significant part in repair and maintenance of DNA, and thus genome stability. Deficiencies of vitamin create hindrance in DNA repair mechanism and cellular dysfunction, such as cancer and aging. Reactive oxygen species (ROS) generated due to catabolism of the dietary components and cellular metabolic processes are very harmful for DNA and enzymes. On the other hand, carotenoids act as an antioxidant and quench the harmful free-radicals, thereby reduce the damage caused by ROS. Concentration of carotenoids in human plasma was reported to be negatively associated with oxidative lymphocyte damage [30]. Similarly, expression of interleukin 6 (IL-6), type I and type II proto-oncogenes, and growth factors like tumor growth factor-b1 are regulated by vitamin A [31].
Folic acid and vitamin B12 are required for DNA metabolism. Folate is important for the synthesis of deoxythymidine monophosphate (dTMP) from deoxyuridine monophosphate (dUMP). Deficiency of folate results in the incorporation of uracil in DNA, resulting in breaks in the DNA and increased requirements of efficient DNA repair mechanisms. Some of the major roles of vitamin B12 and folate include synthesis of methionine and S-adenosyl-methionine (SAM), which are needed to maintain the epigenomic architecture [32]. Studies indicate that single nucleotide polymorphism (SNP) in the genome of individuals might be responsible for functional variations in the proteins/enzymes necessary for synthesis, transport and metabolism of vitamins [33]. Vitamin B12 has been reported to have inducive effect on the synthesis of methionine synthase with prompt post-transcriptional modulation in mammalian cell cultures. Vitamin C also acts as an antioxidant, and plays important role in minimizing oxidative damage to DNA. Studies indicate that vitamin C affects expression of the genes for glutathione S-transferase Pi, p73, fra-1, Mut L homologue-1 (MLH1). MLH1 is specifically important in DNA repair pathways, as its up-regulated expression is essential for DNA defense mechanism [34]. Thus, optimal intake of vitamins is essential to protect the genome. It has been established now that nutrients like carbohydrates, amino acids and vitamins are required for defense, repair and maintenance of DNA, and integrity of genome; thereby, nutrients play crucial roles in genome stability and gene expression.
Epigenetics and Epigenomic Modifications
Epigenome is the sum total of the changes in the genome (without altering the nucleotide sequence), and studies of such changes are known as epigenomics. More importantly, these epigenetic changes are reversible in nature; thereby, play important roles in regulation of gene expression particularly under varying environmental conditions [6]. Epigenetic modifications may alter chromatin conformation either by making it more compact (heterochromatin), and thus suppressing transcription of the genes present in the region, or by making it (euchromatin) easily accessible to transcription machinery [4]. In addition, certain amino acids at the N-terminal tails of histone proteins are post- translationally modified that may have profound impacts on condensation/segregation of chromosomal regions [7]. Non-coding RNAs like small-RNAs (siRNA and miRNA) and lncRNA biogenesis play vital role in DNA methylation through RNA-directed DNA methylation (RdDM) pathway [35]. More importantly, some of the epigenetic changes may be inherited to the next generation that might result in phenotypic variations [8]. Evidence suggests the role of epigenetic changes in regulation of gene expression during growth, development, and environmental fluctuations; hence, it would be crucial to investigate the role of nutrients in managing epigenetic architecture of the genome/ epigenome. This may help utilizing dietary components/ nutrients to manage epigenetic landscape, which may help treating various metabolic disorders [9].
DNA Methylation is Affected by Nutrients
DNA methylation is catalyzed by DNA methyltransferase (DNMT) family of enzymes, which is responsible for modulating transcription in association of the altered chromatin state [4]. Dietary folate contains methyl group and delivers it during the formation of AdoMet. Apart from folate, other dietary components like methionine, betaine, vitamin B2 and choline are determinants of DNA methylation. A study on mouse indicated that folate concentration is significantly correlated with altered methylation at p16 promoter, and change in expression of the gene in mouse colon [11]. Choline is an essential micro-nutrient that plays a vital role in DNA methylation. Studies showed that supplementation of choline increases DNA methylation with enhanced DNMT expression in hepatic cells (HepG20) of human being [36]. When human T lymphocytes were cultured in a medium containing lower concentration of methinonine and folic acid, DNA demethylation and overexpression of certain genes associated with autoimmunity was observed [24]. An in vivo study on sheep suggested that dietary deficiency of folate, methionine and vitamin B12 trigger obesity in offspring along with inadequate immune response. The study indicates that deficiency of dietary components during the periconceptional period may have potential effects on altering DNA methylation pattern even in offspring [16].
Vitamin B12 is a vital cofactor for methionine synthase, and its availability impacts genomic DNA methylation [37]. Recent study shows that certain bioactive food compounds like polyphenols (tea), isothiocyanates and genistein (soybean) might be involved in decreasing the risks of cancer by reducing DNA methylation from the genes associated with cancer [38]. Polyphenols can have direct inhibitory effects on the catalytic site of DNA methyltransferase 1 or it can have indirect effect on DNA methylation status [39]. Alcohol has profound impacts on methyl group transfer reaction. Study on mouse model shows that expression of certain epigenetically sensitive alleles like Agouti viable yellow (Avy) is affected by the exposure of maternal parent to ethanol [19]. It was suggested that ethanol might affect transcriptional silencing, which corroborate with hypermethylation at Avy, leading to agouti-colored coat in mouse. Thus, certain nutrients and bioactive dietary components have the potential to alter DNA methylation pattern, and indirectly affect gene expression as well as various patho-physiological conditions.
Histone Modifications and Effects of Bioactive Food Components
Histone modulation works with DNA methylation to epigenetically regulate gene expression. Numerous evolutionary conserved lysine (K) residues in the histone proteins are subjected to various modifications like acetylation, methylation, ubiquitylation, biotinylation, etc [7]. Two enzymes, namely histone acetyltransferase (HAT) and histone deacetylase (HDAC), control acetylation level of the histone protein towards regulation of gene expression [40]. More importantly, certain bioactive food components impact the activity of HAT and HDAC [41]. Bioactive food compounds like sulforaphane (found in broccoli and sproutes), diallyl sulfide (from garlic) and short chain fatty acid (e.g. butyrate) act as HDAC inhibitors. In vitro melanoma cell (B16 and S91) culture study inferred that sulforaphane has the potential to reduce deacetylation activity, thereby inhibits growth and proliferation of cancerous cells [42]. Diallyl sulfide was reported to have the potential to enhance H3 and H4 acetylation in rat colon cells [43]. Another study showed that tributyrate can increase hepatic nuclear H3K9 acetylation by 4-fold in preneoplastic lesions in rat, which also increases expression of p21 protein by 1.5-fold that acts as HDAC inhibitory effect [44]. Acetylation/deacetylation of histone proteins is associated with expression of pro-inflammatory genes (e.g. interlukin-1, 8, 12) and anti-inflammatory genes (like IL-10) [45, 46, 25]. Resveratrol present in grape skin acts as activator of Sirt1, and has anti-inflammatory effects against colitis and colitis-associated colon cancer [22, 47]. Studies also indicate that resveratrol can repress NF-kB and acetylation of histone proteins. Histone methylation/ demethylation was reported to be linked with obesity in mice [48]. Loss of activity of H3K9-specific demethylase (Jhdm2a) resulted in obesity and hyperlipidemia, suggesting that H3K9 methylation plays important role in regulation of metabolism associated genes. Biotin also play important role in modification of H3 andH4 via covalent attachment of biotin to specific lysine residues. Thus, dietary deficiency of biotin might have impacts on chromatin structure and gene expression [49].
Non-coding RNAs Helps Modulating Gene Expression
Non-coding RNA (ncRNA), a functional RNA produced through transcription but not translated into proteins, is another epigenetic mark that affects gene expression, and thus many cellular processes [50]. Epigenetic modifications like DNA methylation and histone modifications are mediated through ncRNAs, these may lead to some pathological conditions like cancer [51]. Various bioactive food components affecting miRNA biogenesis and cancerous growth are listed in Table 2. Study also indicated that diet deficient in methyl group donor, cause formation of tumors in rat liver along with the changes in biogenesis of miRNAs like miR34a, miR127, and miR16a that are involved in the regulation of programmed cell death, proliferation of cells and intercellular connection [52]. Cirera, et al. [53] reported 1.4-fold reduced expression of miR122 in pigs when they were fed with high-cholesterol diet compared to those fed with the standard diet, which indicates implication of miRNA/epigenetics in obesity.
| Bioactive food component | Effect | References |
|---|---|---|
| Genistein | Represses human uveal melanoma cells and murine chronic lymphocytic leukemia cells by altering miR-16. | [54] |
| Curcumin | Represses human pancreatic cancer cells by upregulating and downregulating miR-22 and miR-199a respectively. | [55] |
| Retinoic acid | Retinoic acid receptor antagonists sufficiently effectively repress miR-10a expression and block metastasis and miR-34a functions as a potential tumor suppressor in neuroblastoma cells. | [56] |
| Polyunsaturated fatty acids | During initial stage of cancer progression several microRNAs such as let-7d, miR-15b, miR-107, miR-191 and miR-324-5p were significantly affected by diet-carcinogen interactions. It is inferred that fish-oil-fed animals differentially expressed smallest microRNAs thus, concluding the potential of fish oil in protecting the colon from carcinogen-induced microRNA dysregulation. | [57] |
| Curcumin | Upregulation of miR-15a and miR-16 expression, both of which could inhibit the expression of B-cell lymphoma 2 (Bcl-2), thereby inducing apoptosis in MCF-7 breast cancer cells. | [58] |
Table 2: Bioactive food components affecting cancer through miRNA biogenesis.
Nutriepigenomics: Integration of Epigenetics, Nutrition and Health
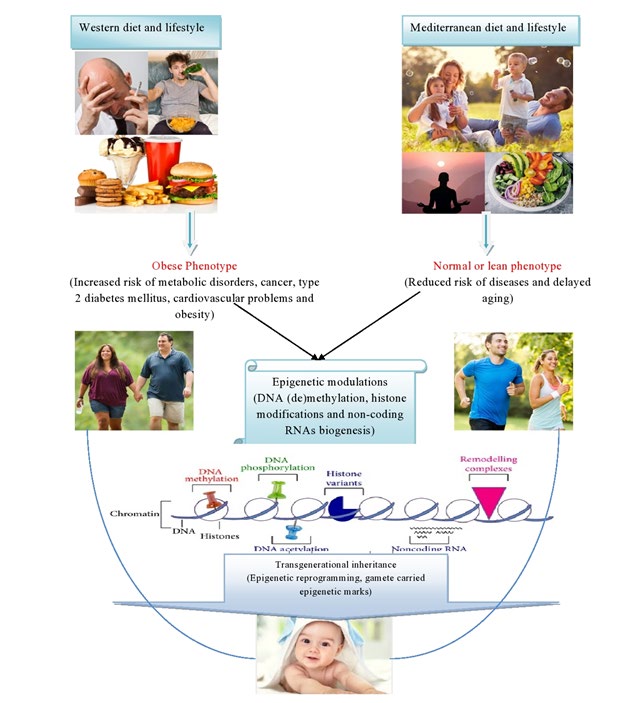
Dietary behavior not only affects nutritional requirements of the body but also have influence on epigenome and phenotype of the individual as well as its offspring. Westernized diets are mainly comprised of the food rich in saturated fats and meat, lesser carbohydrates, cereals/ grains, fruits and fresh vegetables. When westernized diet is combined with static lifestyle, severe alcohol consumption, smoking, it may lead to the occurrence of diseases like hypertension, cardiovascular disorders CVDs), obesity and cancer [59]. The impact of the diet rich in saturated fatty acid and other harmful components was assessed towards the development of non-alcoholic fatty liver disease (NAFLD) in rodent. An imbalanced ratio of n-6 polyunsaturated fatty acids (PUFA) and n-3 PUFA in Western diets increases the risk of NAFLD. Non-alcoholic steatohepatitis (NASH), a type of NAFLD, results in the accumulation of lipids in liver, fibrosis and cirrhosis/liver cancer. The risk increases due to the reduced content of linoleic acid [60]. Decreased n-6:n-3 ratio was reported to prevent NASH by attenuating oxidative stress [61]. Studies suggest strong correlation between DNA methylation level in peripheral blood and eicosapentaenoic acid/arachidonic acid content in milk-fed infants and adult men [62, 63]. Dsylipidemia in pregnant mother was reported to cause epigenetic alterations in placenta and liver of the fetus [64]. On the other hand, Mediterranean diets have been reported to reduce the risk of CVDs, obesity and other health problems. Mediterranean diet is composed of fruits, fresh vegetables, multigrain food, fish, egg, and healthy fats. Human population fed with Mediterranean diet along with olive oil and nuts showed lower risk of CVDs and cancer compared to their counterparts [65, 66]. Regular Mediterranean diet during the early pregnancy was reported to be linked with positive cognitive behavior in the offspring [67] (Figure 2).
Nutrition, being the most extensively studied environmental factor, is associated with epigenetic changes in the genome (epigenome), health of the organism, and metabolic disorders [68]. Common dietary components like folate and choline can modulate DNA methylation and impact gene expression. Methyl-enriched diet is essential during the early days of pregnancy is essential for proper growth and development of fetus, otherwise chances of diseases and cancer increase significantly [69]. An NAD+-dependent HDAC (Sirtuin 1) has been reported to be activated by several dietary components like resveratrol, and Sirtuin 1 delays some of the physiological processes related with aging via its impact on DNA methylation [70]. Balanced nutrition during the early life prompts epigenetic alterations that affect individual’s health and disorders. Nutrients directly affect the activity of enzymes (DNMT and HDAC) linked with epigenetic modifications [71]. Malnutrition during the early months of pregnancy have higher risk of diseases in fetus through epigenetic variations, and these might have impacts throughout the life. The effects of nutritional status of grandmother’s life (who faced malnutrition/famine) on the health of female grandchildren, in terms of the risks of mortality due to CVDs and body mass index (BMI), were observed to be higher compared to that in its counterpart [72, 2].
Studies report significant association between epigenetic changes in the genes involved in glucose/insulin metabolism and obesity [73, 74, 75, 76, 77]. Increased methylation of HIF3A was reported to be correlated with higher BMI [73]. Number of studies has been carried out to analyze DNA methylation and its association with obesity. DNA methylation drastically changes during ageing, and it has become clear that better nutrition during early life has the potential to modulate DNA methylation and improve longevity [71]. Thus, consumption of junk food and negligible exercise affect the risk of obesity, which might be controlled through genetic predisposition and/or epigenetic alterations [78].
Nutriepigenomics and Cancer
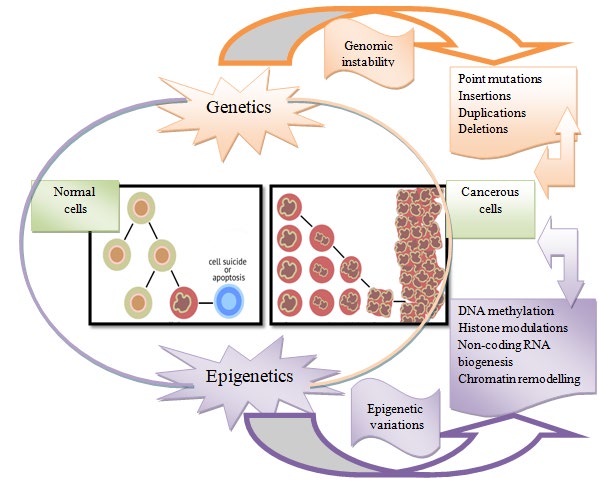
Different nutrients play important roles in epigenetic changes in the genome/epigenome which affect growth, development, and patho-physiological conditions of the organism. Epigenetic alteration in the genetic material is significantly affected by some of the nutrients like folate and vitamin B12. In vivo study indicates that presence of folate in diet is responsible for the expression of p16 tumor suppressor gene, which is associated with DNA methylation in its promoter. A tumor suppressor gene (p16INK4a) was reported to be target of epigenetic inactivation in human breast carcinogenesis [79]. Low intake of folate has been associated with hypomethylation, increased risks of colorectal and pancreatic cancers. Folate was reported for modifying anticarcinogenic properties via epigenetic alterations [1]. The potential effects of dietary components present in cereals, fruits, vegetables and other Mediterranean diets via epigenetic changes against cancer have been presented in Table 3. DNA methylation, histone modulations, and ncRNAs are involved in controlling the occurrence of cancer indirectly through epigenetic modifications. Diets deficient in methyl-group donors and folate result in aberrant expression of miRNA; thus, abnormalities in cell proliferation and apoptosis leading to cancer [80]. Genetic and epigenetic alterations causing conversion of normal cells into cancerous cells are represented in Figure 3.
| Nutrient | Dietary sources | Epigenetic alteration |
|---|---|---|
| Methionine | Spinach, Fish, Peppers, Brazil nuts, Sesame seeds, etc. | S-adenosyl methionine (SAM) synthesis |
| Folic acid | Leafy vegetables, Sunflower seeds, Baker’s yeast, Liver, etc. | Methionine synthesis. |
| Retinoic acid | Dark green, red and yellow vegetables, fish and cod liver oil, etc. | miRNA regulation |
| Vitamin B 6 | Vegetable, Nuts, Whole grain products, Meats, etc. | Methionine synthesis |
| Vitamin B 12 | Milk, Shellfish, Meat, Liver, etc. | Methionine synthesis |
| SAM-e | Dietary supplements, Supplementary pills, etc. (unstable in food) | With the help of enzymes, transfer methyl group directly to DNA. |
| Polyphenols | Green tea and other plant sources. | Inhibition of DNA methylation |
| Curcumin | Turmeric | miRNA regulation |
| Betaine | Wheat, Spinach, Sugar beet, Shellfish, etc. | Break down the toxic products of SAM synthesis. |
| Resveratrol | Red wine | Removes acetyl groups from histones. |
| Butyrate | Produced on fermentation of dietary fibre in intestine. | Increase histone acetylation. |
| Genistein | Soybean | Increase methylation. |
| Sulforaphane, Isothiocyanate | Broccoli sprouts | Increase histone acetylation. |
| (Act as HDAC inhibitor) | ||
| Diallyl sulphide | Garlic | Increase histone acetylation. |
| (Act as HDAC inhibitor) |
Table 3: Epigenetic role of nutrients and bioactive food components in cancer.
Environmental Epigenetics
In addition to the nutritional status, various other environmental conditions like exposure to certain chemicals might contribute to epigenetic changes in the genome. Exposure of organisms to some of the commonly present hazardous metals (arsenic), organic compounds, alcohol, pesticides, and endocrine disrupting chemicals (EDCs) might induce epigenetic alterations. Human beings are frequently exposed to a broad range of EDCs which interfere with pathways of hormone signaling. EDCs related exposure results in epigenetic alterations and play role in carcinogenesis [1]. EDCs have impact on the functioning of epigenetics related enzymes (DNMTs, HATs, and HDACs) [81]. Moreover, smoking and alcohol lead to emphysema (Smoker’s disease) and cancers, as smoke and alcohol exert dysregulation of cellular processes through epigenetic changes. In vitro and in vivo studies indicate that alcohol has deleterious effects on metabolism and nervous system development through down regulated epigenetic changes at early embryonic stage. This may affect learning ability, memory processes and abnormal growth and development of fetus [82]. Human being comes in contact of several chemicals, insecticides, pesticides etc. used in agriculture, society and industries, directly or indirectly. Moreover, chemical/pesticide residues in crop produce/ food, pollutants in air/water/soil cause environmental pollutions and health hazards [83]. Tobacco consumption and smoking habits in pregnant mothers have been reported to cause epigenetic alterations in buccal mucosal cells, and serious consequences on child’s health and growth with impaired neural development, asthma, allergies, and immune dysfunctions have been reported [84, 1].
Future Perspectives
Nutrition is one of the major environmental factors that affect organism’s growth, development and health conditions. Bioactive food components and dietary pattern, along with the other environmental factors, affect epigenomic landscape of the organism. Since nutriepigenomics is quite novel and an emerging branch of science, intensive scientific investigation on the impact of bioactive food components in epigenetic alterations is very much desirable. Moreover, the effects of epigenetic alterations caused by the dietary components and other environmental factors on the organism’s phenotype, as well as their inheritance, would be mandatory. In future, the extensive studies on deciphering epigenetic alterations caused by the nutrients might open the way for personalized nutritional interventions to help managing better growth, development, and healthy life. This might also help opening up new therapeutic interventions in cancer research [85, 86]. Healthy diets, including fruits and fresh vegetables, have significant effects on metabolic processes which determines cellular environment, and thus affect cellular growth and immune functions. Changes in DNA methylation in cancer causing genes is a fascinating epigenetic marker that facilitate detecting the tumor at the early stage of development and its efficient treatment [8].
Furthermore, epigenetic manipulation using CRISPR- dCas9 technology might help performing epigenome editing in future not only for sustainable food production but also for curing/preventing patho-physiological conditions [87]. However, several other basic information needs to be generated, challenges to be taken up, and considerable efforts would require to be made to decipher the pathways and epigenetic marks associated with the trait of interest. Such efforts might pave the ways for prognosis and treatment of various life-threatening diseases like cancer [88, 89]. However, it would also be essential to maintain the edited epigenetic landscape in the cell/tissue by creating the cellular environment necessary for the epigenetic state either by deploying the essential epigenetic machinery or by supplying the essential nutrient(s). More importantly, the biosafety issues of laboratory research [90] as well as those of the genetically modified organisms [91], as per the regulatory guidelines of the country, must be taken care to maximise the overall benefits [92].
Conclusions
Nutrients constitute the cellular environment necessary for epigenetic modifications in the cell. Different kinds of epigenetic modifications like DNA methylation, histone protein modifications, biogenesis of ncRNAs, chromatin remodeling are interlinked, and indirectly affected by nutritional status and environmental conditions. Nutrients play vital roles in epigenetic modifications and affect patho- physiological conditions that emergence of a new branch of science namely nutriepigenomics has initiated, which is expected to help managing some of the life-threatening diseases. However, unravelling the role of nutrients on epigenetic alterations toward controlling complex traits can be intimidating process. Moreover, healths, physiological and pathological conditions, aging related complications are the consequences of integrated action of genetics, epigenetics, nutrition and environmental factors. Lifestyle, behavior, eating patterns and various other environmental factors leave epigenetic marks on genetic material that affects expression of genes, tolerance to environmental stresses, and patho- physiological conditions. Therefore, deeper understanding of the cross-talk among nutrition, epigenomics and patho- physiology might help circumventing the health-related issues like CVDs, obesity and cancer. Future research in nutriepigenomics has unprecedented opportunity to benefit public health, and personalized nutrition. Understanding the role of nutrients, particularly in epigenetic modifications, along with their sources and metabolism in the body would be very essential for targeted discoveries and therapeutic interventions for numerous crucial/chronic diseases.
The views expressed herein are those of the authors only, and these may not necessarily be the views of the institution/ organization the authors are associated with.
Acknowledgments
Epigenomics and epitranscriptomics research work is being carried out with the financial support from National Agricultural Science Fund (NASF/ABP-70161/2018-1), and Extramural Research grants, Indian Council of Agricultural Research, New Delhi, India.
References
-
Tiffon C (2018) The Impact of Nutrition and Environmental Epigenetics on Human Health and Disease. International Journal of Molecular Sciences 19(11): 3425.
-
Aiken CE, Tarry Adkins JL, Ozanne SE (2016) Transgenerational effects of maternal diet on metabolic and reproductive ageing. Mammalian Genome 27(7-8): 430-439.
-
Singh A, Bhushan B, Gaikwad K, Yadav OP, Kumar S, et al. (2015) Induced defence responses of contrasting bread wheat genotypes under differential salt stress imposition. Indian Journals of Biochemistry and Biophysics 52(1): 75-85.
-
Kumar S (2017) Epigenetic control of apomixis: a new perspective of an old enigma. Advances in Plants and Agriculture Research 7(1): 227-233.
-
Kumar S, Chinnusamy V, Mohapatra T (2018) Epigenetics of modified DNA bases: 5-methylcytosine and beyond. Frontiers in Genetics 9: 1-14.
-
Kumar S (2018) Epigenomics of plant responses to environmental stress. Epigenomes 2(1): 6.
-
Kumar S, Singh AK, Mohapatra T (2017) Epigenetics: history, present status and future perspective. Indian Journal of Genetics and Plant Breeding 77(4): 445-463.
-
Kumar S (2018) Environmental stress, food safety, and global health: biochemical, genetic and epigenetic perspectives. Medical Safety and Global Health 7: e145.
-
Kumar S (2019) Epigenetics and epigenomics for crop improvement: current opinion. Advances in Biotechnology and Microbiology 14(1): 555879.
-
Choi SW, Friso S (2010) Epigenetics: A New Bridge between Nutrition and Health. Advances in Nutrition 1(1): 8-16
-
Keyes MK, Jang H, Mason JB, Liu Z, Crott JW, et al. (2007) Older age and dietary folate are determinants of genomic and p16-specific DNA methylation in mouse colon. Journal of Nutrition 137(7): 1713-1717.
-
Serefidou M, Venkatasubramani AV, Imhof A (2019) The impact of one carbon metabolism on histone methylation. Frontiers in Genetics 10: 764.
-
Jin SG, Kadam S, Pfeifer GP (2010) Examination of the specificity of DNA methylation profiling techniques towards 5-methylcytosine and 5-hydroxymethylcytosine. Nucleic Acids Research 38: e125.
-
Kirkland JB (2009) Niacin status impacts chromatin structure. Journal of Nutrition 139(12): 2397-2401.
-
Mali P, Chou BK, Yen J, Ye Z, Zou J, et al. (2010) Butyrate greatly enhances derivation of human induced pluripotent stem cells by promoting epigenetic remodeling and the expression of pluripotency-associated genes. Stem Cells 28(4): 713-720.
-
Sinclair KD, Allegrucci C, Singh R, Gardner DS, Sebastian S, et al. (2007) DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc Natl Acad Sci USA 104(49): 19351-19356.
-
Steegers-Theunissen RP, Obermann-Borst SA, Kremer D, Lindemans J, Siebel C, et al. (2009) Periconceptional maternal folic acid use of 400 microg per day is related to increased methylation of the _IGF2_ gene in the very young child. PLoS ONE 4: e7845.
-
Niculescu MD, Craciunescu CN, Zeisel SH (2006) Dietary choline deficiency alters global and gene-specific DNA methylation in the developing hippocampus of mouse fetal brains. FASEB J 20(1): 43-49.
-
Kaminen-Ahola N, Ahola A, Maga M, Mallitt KA, Fahey P, et al. (2010) Maternal ethanol consumption alters the epigenotype and the phenotype of offspring in a mouse model. PLoS Genetics 6: e1000811.
-
Mehedint MG, Niculescu MD, Craciunescu CN, Zeisel SH (2010) Choline deficiency alters global histone methylation and epigenetic marking at the Re1 site of the calbindin 1 gene. FASEB Journal 24(1): 184-195.
-
Chiu J, Khan ZA, Farhangkhoee H, Chakrabarti S (2009) Curcumin prevents diabetes-associated abnormalities in the kidneys by inhibiting p300 and nuclear factor-kappa B. Nutrients 25(9): 964-972.
-
Cui X, Jin Y, Hofseth AB, Pena E, Habiger J, et al. (2010) Resveratrol suppresses colitis and colon cancer associated with colitis. Cancer Prevention Research (Phila Pa) 3(4): 549-559.
-
Pogribny IP, Starlard-Davenport A, Tryndyak VP, Han T, Ross SA, et al. (2010) Difference in expression of hepatic microRNAs miR-29c, miR-34a, miR-155, and miR- 200b is associated with strain specific susceptibility to dietary nonalcoholic steatohepatitis in mice. Laboratory Investigation 90(10): 1437-1446.
-
Li Y, Liu Y, Strickland FM, Richardson B (2010) Age- dependent decreases in DNA methyltransferase levels and low transmethylation micronutrient levels synergize to promote overexpression of genes implicated in autoimmunity and acute coronary syndromes. Experimental Gerontology 45(4): 312-322.
-
Qiu X, Brown KV, Moran Y, Chen D (2010) Sirtuin regulation in calorie restriction. Biochimica et Biophysica Acta 1804(8): 1576-1583.
-
Sinclair KD, Allegrucci C, Singh R, Gardner DS, Sebastian S, et al. (2007) DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc Natl Acad Sci USA 104(49): 19351-19356.
-
Uekawa A, Katsushima K, Ogata A, Kawata T, Maeda N, et al. (2009) Change of epigenetic control of cystathionine beta-synthase gene expression through dietary vitamin B12 is not recovered by methionine supplementation. Journal of Nutrigenetics and Nutrigenomics 2(1): 29-36.
-
Pop S, Enciu AM, Tarcomnicu I, Elvira G, Cristiana T (2019) Phytochemicals in cancer prevention: modulating epigenetic alterations of DNA methylation. Phytochemistry Reviews 18: 1005-1024.
-
Li Y, Tollefsbol TO (2010) Impact on DNA methylation in cancer prevention and therapy by bioactive dietary components. Current Medicinal Chemistry 17(20): 2141-2151.
-
Kaminen-Ahola N, Ahola A, Maga M, Mallitt KA, Fahey P, et al. (2010) Maternal ethanol consumption alters the epigenotype and the phenotype of offspring in a mouse model. PLoS Genetics 6: e1000811.
-
Jambhekar A, Dhall A, Shi Y (2019) Roles and regulation of histone methylation in animal development. Nature Reviews Molecular Cell Biology 20: 625-641.
-
Nian H, Delage B, Ho E, Dashwood RH (2009) Modulation of histone deacetylase activity by dietary isothiocyanates and allyl sulfides: studies with sulforaphane and garlic organosulfur compounds. Environmental and Molecular Mutagenesis 50(3): 213-221.
-
Do DP, Pai SB, Rizvi SA, D’Souza MJ (2010) Development of sulforaphane encapsulated microspheres for cancer epigenetic therapy. International Journal of Pharmacy 386: 114-121.
-
Druesne-Pecollo N, Chaumontet C, Pagniez A, Vaugelade P, Bruneau A, et al. (2007) _In vivo_ treatment by diallyl disulfide increases histone acetylation in rat colonocytes. Biochemical and Biophysical Research Communications 354: 140-147.
-
Kuroiwa-Trzmielina J, de Conti A, Scolastici C, Pereira D, Horst MA, et al. (2009) Chemoprevention of rat hepatocarcinogenesis with histone deacetylase inhibitors: efficacy of tributyrin, a butyric acid prodrug. International Journal of Cancer 124(11): 2520-2527.
-
Villagra A, Sotomayor EM, Seto E (2010) Histone deacetylases and the immunological network: implications in cancer and inflammation. Oncogenesis 29: 157-173.
-
Coward WR, Watts K, Feghali-Bostwick CA, Knox A, Pang L (2009) Defective histone acetylation is responsible for the diminished expression of cyclooxygenase 2 in idiopathic pulmonary fibrosis. Molecular and Cell Biology 29(15): 4325-4339.
-
Qiu X, Brown KV, Moran Y, Chen D (2010) Sirtuin regulation in calorie restriction. Biochimica et Biophysica Acta 1804(8): 1576-1583.
-
Cui X, Jin Y, Hofseth AB, Pena E, Habiger J, et al. (2010) Resveratrol suppresses colitis and colon cancer associated with colitis. Cancer Prevention Research (Phila Pa) 3(4): 549-559.
-
Sanchez-Fidalgo S, Cardeno A, Villegas I, Talero E, de la Lastra CA (2010) Dietary supplementation of resveratrol attenuates chronic colonic inflammation in mice. European Journal of Pharmacology 633(1-3): 78-84.
-
Tateishi K, Okada Y, Kallin EM, Zhang Y (2009) Role of Jhdm2a in regulating metabolic gene expression and obesity resistance. Nature 458: 757-761.
-
Kuroishi T, Rios-Avila L, Pestinger V, Wijeratne SS, Zempleni J (2011) Biotinylation is a natural, albeit rare, modification of human histones. Molecular Genetics and Metabolism 104(4): 537-545.
-
Huang B, Jiang C, Zhang R (2014) Epigenetics: The language of the cell? Epigenomics 6: 73-88.
-
Iorio MV, Piovan C, Croce CM (2010) Interplay between microRNAs and the epigenetic machinery: an intricate network. Biochimica et Biophysica Acta 1799(10-12): 694-701.
-
Tryndyak VP, Ross SA, Beland FA, Pogribny IP (2009) Down-regulation of the microRNAs miR-34a, miR-127, and miR-200b in rat liver during hepatocarcinogenesis induced by a methyl-deficient diet. Molecular Carcinogenesis 48: 479-487.
-
Cirera S, Birck M, Busk PK, Fredholm M (2010) Expression profiles of miRNA-122 and its target _CAT1_ in minipigs (Sus scrofa) fed a high cholesterol diet. Comparative Medicine 60: 136-141.
-
Lytle KA, Wong CP, Jump DB (2017) Docosahexaenoic acid blocks progression of western diet-induced nonalcoholic steatohepatitis in obese _Ldlr_-mice. PLoS ONE. 12: e0173376.
-
Jensen VS, Hvid H, Damgaard J, Nygaard H, Ingvorsen C, et al. (2018) Dietary fat stimulates development of NAFLD more potently than dietary fructose in Sprague-Dawley rats. Diabetology and Metabolic Syndromre 10: 4.
-
Jeyapal S, Kona SR, Mullapudi SV, Putcha UK, Gurumurthy P, et al. (2018) Substitution of linoleic acid with α-linolenic acid or long chain n-3 polyunsaturated fatty acid prevents Western diet induced nonalcoholic steatohepatitis. Scientific Reports 8: 10953.
-
De La Rocha C, Pérez-Mojica JE, Zenteno-De LS, Cervantes- Paz B, Tristán-Flores FE, et al. (2016) Associations between whole peripheral blood fatty acids and DNA methylation in humans. Scientific Reports 6: 25867.
-
Ruegsegger GN, Grigsby KB, Kelty TJ, Zidon TM, Childs TE, et al. (2017) Maternal Western diet age-specifically alters female offspring voluntary physical activity and dopamine-and leptin-related gene expression. FASEB J 31(12): 5371-5383.
-
Ramaiyan B, Talahalli RR (2018) Dietary unsaturated fatty acids modulate maternal dyslipidemia-induced DNA methylation and histone acetylation in placenta and fetal liver in rats. Lipids 53(6): 581-588.
-
Estruch R, Ros E, Salas-Salvadó J, Covas MI, Corella D, et al. (2018) Primary prevention of cardiovascular disease with a Mediterranean diet supplemented with extra- virgin olive oil or nuts. New England Journal of Medicine 378: e34.
-
Salerno E, Scaglione BJ, Coffman FD, Brown BD, Baccarini A, et al. (2009) Correcting miR-15a/16 genetic defect in New Zealand Black mouse model of CLL enhances drug sensitivity. Molecular Cancer Therapeutics 8(9): 2684- 2692.
-
Sun M., Estrov Z, Ji Y, Coombes KR, Harris DH, et al. (2008) Curcumin (diferuloylmethane) alters the expression profiles of microRNAs in human pancreatic cancer cells. Molecular Cancer Therapeutics 7(3): 464-73.
-
Weiss FU, Marques IJ, Woltering JM, Vlecken DH, Aghdassi A, et al. (2009) Retinoic acid receptor antagonists inhibit miR-10a expression and block metastatic behavior of pancreatic cancer. Gastroenterology 137(6): 2136-2145.
-
Davidson LA, Wang N, Shah MS, Lupton JR, Ivanov I, et al. (2009) _n_-3 polyunsaturated fatty acids modulate carcinogen-directed non-coding microRNA signatures in rat colon. Carcinogenesis 30(12): 2077-2084.
-
Yang J, Cao Y, Sun J, Zhang Y (2009) Curcumin reduces the expression of _Bcl-2_ by upregulating miR-15a and miR-16 in MCF-7 cells. Medical Oncology 27(4): 1114-1118.
-
Lillycrop KA, Hoile SP, Grenfell L, Burdge GC (2014) DNA methylation, ageing and the influence of early life nutrition. Proceedings of Nutrition Society 73(3): 413- 421.
-
Rando OJ, Simmons RA (2015) I’m eating for two: parental dietary effects on offspring metabolism. Cell 161(1): 93-105.
-
Dick KJ, Nelson CP, Tsaprouni L, Sandling JK, Aissi D, et al. (2014) DNA methylation and body-mass index: a genome-wide analysis. Lancet 383: 1990-1998.
-
Wang S, Song J, Yang Y, Zhang Y, Wang H, et al. (2015) _HIF3A_ DNA methylation is associated with childhood obesity and ALT. PLoS One 10: e0145944.
-
Main AM, Gillberg L, Jacobsen AL, Nilsson E, Gjesing AP, et al. (2016) DNA methylation and gene expression of HIF3A: cross-tissue validation and associations with BMI and insulin resistance. Clinical Epigenetics 8(1): 1-7.
-
Pfeiffer S, Kruger J, Maierhofer A, Bottcher Y, Kloting N, et al. (2016) Hypoxia-inducible factor 3A gene expression and methylation in adipose tissue is related to adipose tissue dysfunction. Scientific Reports 6: 27969.
-
Richmond RC, Sharp GC, Ward ME, Fraser A, Lyttleton O, et al. (2016) DNA methylation and BMI: investigating identified methylation sites at _HIF3A_ in a causal framework. Diabetes 65(5): 1231-1244.
-
Rohde K, Keller M, Poulsenc LLC, Bluher M, Kovacs P, et al. (2019) Genetics and epigenetics in obesity. Metabolism Clinical and Experimental 92: 37-50.
-
Witcher M, Emerson BM (2009) Epigenetic silencing of the _p16__I_NK4a tumor suppressor is associated with loss of CTCF binding and a chromatin boundary. Molecular Cell 15: 271-284.
-
Larsson SC, Giovannucci E, Wolk A (2006) Folate intake, _MTHFR_ polymorphisms, and risk of esophageal, gastric, and pancreatic cancer: A meta-analysis. Gastroenterology 131: 1271-1283.
-
Alavian-Ghavanini A, Ruegg J (2018) Understanding Epigenetic Effects of Endocrine Disrupting Chemicals: From Mechanisms to Novel Test Methods. Basic and Clinical Pharmacology and Toxicology 122(1): 38-45.
-
Farris SP, Wolen AR, Miles MF (2010) Using expression genetics to study the neurobiology of ethanol and alcoholism. International Review of Neurobiology 91: 95-128.
-
Kumar S (2015) Biopesticide: an environment friendly pest management strategy. Journal of Biofertilizers and Biopesticides 6: e127.
-
Knopik VS, Maccani MA, Francazio S, McGeary JE (2012) The epigenetics of maternal cigarette smoking during pregnancy and effects on child development. Development and Psychopathology 24(4): 1377-1390.
-
Ning B, Li W, Zhao W, Wang R (2016) Targeting epigenetic regulations in cancer. Acta Biochimica et Biophysica Sinica 48(1): 97-109.
-
Nicodemus-Johnson J, Sinnott RA (2017) Fruit and juice epigenetic signatures are associated with independent immunoregulatory pathways. Nutrients 9(7): 752.
-
Kumar S (2019) Genome editing to epigenome editing: towards unravelling the enigmas in developmental biology. Trends in Developmental Biology 12: 31-38.
-
Xu X, Tao Y, Gao X, Zhang L, Li X et al. (2016) A CRISPR- based approach for targeted DNA demethylation. Cell Discovery 2: 16009.
-
Braun SMG, Kirkland JG, Chory EJ, Husmann D, Calarco JP, et al. (2017) Rapid and reversible epigenome editing by endogenous chromatin regulators. Nature Communications 8: 560.
-
Kumar S (2012) Biosafety issues in laboratory research. Biosafety 1: e116.
-
Kumar S (2014) Biosafety issues of genetically modified organisms. Biosafety 3: e150.
-
Mali P, Chou BK, Yen J, Ye Z, Zou J, et al. (2010) Butyrate greatly enhances derivation of human induced pluripotent stem cells by promoting epigenetic remodeling and the expression of pluripotency-associated genes. Stem Cells 28(4): 713-720.
-
Steegers-Theunissen RP, Obermann-Borst SA, Kremer D, Lindemans J, Siebel C, et al. (2009) Periconceptional maternal folic acid use of 400 microg per day is related to increased methylation of the _IGF2_ gene in the very young child. PLoS ONE 4: e7845.
-
Niculescu MD, Craciunescu CN, Zeisel SH (2006) Dietary choline deficiency alters global and gene-specific DNA methylation in the developing hippocampus of mouse fetal brains. FASEB J 20(1): 43-49.
-
Mehedint MG, Niculescu MD, Craciunescu CN, Zeisel SH (2010) Choline deficiency alters global histone methylation and epigenetic marking at the Re1 site of the calbindin 1 gene. FASEB Journal 24(1): 184-195.
-
Chiu J, Khan ZA, Farhangkhoee H, Chakrabarti S (2009) Curcumin prevents diabetes-associated abnormalities in the kidneys by inhibiting p300 and nuclear factor-kappa B. Nutrients 25(9): 964-972.
-
Pogribny IP, Starlard-Davenport A, Tryndyak VP, Han T, Ross SA, et al. (2010) Difference in expression of hepatic microRNAs miR-29c, miR-34a, miR-155, and miR- 200b is associated with strain specific susceptibility to dietary nonalcoholic steatohepatitis in mice. Laboratory Investigation 90(10): 1437-1446.
-
Salerno E, Scaglione BJ, Coffman FD, Brown BD, Baccarini A, et al. (2009) Correcting miR-15a/16 genetic defect in New Zealand Black mouse model of CLL enhances drug sensitivity. Molecular Cancer Therapeutics 8(9): 2684- 2692.
-
Sun M, Estrov Z, Ji Y, Coombes KR, Harris DH, et al. (2008) Curcumin (diferuloylmethane) alters the expression profiles of microRNAs in human pancreatic cancer cells. Molecular Cancer Therapeutics 7(3): 464-73.
-
Weiss FU, Marques IJ, Woltering JM, Vlecken DH, Aghdassi A, et al. (2009) Retinoic acid receptor antagonists inhibit miR-10a expression and block metastatic behavior of pancreatic cancer. Gastroenterology 137(6): 2136-2145.
-
Davidson LA, Wang N, Shah MS, Lupton JR, Ivanov I, et al. (2009) _n_-3 polyunsaturated fatty acids modulate carcinogen-directed non-coding microRNA signatures in rat colon. Carcinogenesis 30(12): 2077-2084.
-
Yang J, Cao Y, Sun J, Zhang Y (2009) Curcumin reduces the expression of _Bcl-2_ by upregulating miR-15a and miR-16 in MCF-7 cells. Medical Oncology 27(4): 1114-1118.
-
Zhang J, Zhang F, Didelot X, Bruce KD, Cagampang FR, Vatish M. et al. (2009) Maternal high fat diet during pregnancy and lactation alters hepatic expression of insulin like growth factor-2 and key microRNAs in the adult offspring. BMC Genomics 10: 478.
-
Xiao J (2015) Phytochemicals in medicine and food. Phytochemistry Reviews 14: 317-320.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends