Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
Tamoxifen (TAM), a selective estrogen receptor modulator, competes with estrogen for binding to estrogen receptor (ER) sites, thereby inhibiting estrogen-induced proliferation in ER-positive breast cancer cells. However, long-term treatment with TAM often leads to resistance, and its mechanisms, particularly involving biochemical markers, remain unclear. This study investigated the effects of TAM (10⁻⁶ M and 10⁻⁸ M) compared to 17β-estradiol (10⁻⁶ M and 10⁻⁸ M) on galectin-3 and laminin expressions in ER-positive MCF-7 breast cancer cells. The MTT assays showed that 10⁻⁶ M TAM significantly reduced cell viability and altered cellular morphology. Indirect immunofluorescence microscopy revealed that galectin-3 was present on the cell surface and in the nucleus; however, its localization was diminished after 48 hours of TAM (10⁻⁶ M) exposure. Western blot analysis showed no change in galectin-3 expression at 24 hours but a significant reduction at 48 and 72 hours. In contrast, laminin expression remained unchanged. ELISA results further confirmed reduced galectin-3 but not laminin levels after 48 hours of TAM treatment. These findings suggest that galectin-3, but not laminin, is involved in TAM-induced suppression of MCF-7 cell growth and may serve as a biochemical marker for TAM responsiveness.
Segree A¹, Flemmings S¹, Nagdas S² and Raychoudhury S*¹
¹Department of Biology, Benedict College, Columbia, USA ²Fayetteville State University, Fayetteville, North Carolina, USA *Corresponding author: Samir S Raychoudhury, Professor, Department of Biology, Benedict College, 1600 Harden Street, Columbia, SC 29204, USA, Tel: (803) 705-4648; Email: samir. raychoudhury@benedict.edu
Abbreviations
TAM: Tamoxifen; ER: Estrogen Receptor; ECM: Extra Cellular Matrix; FBS: Fetal Bovine Serum
Introduction
Galectins are a family of nonintegrin β-galactoside- binding lectins with related amino acid sequences [1]. Galectin-3, formerly known as CBP35, Mac-2, and εBP, because of its affinity for IgE and HLB31 and its relationship with laminin. It is a 31 kDa-galactoside-binding lectin and a member of the galectin family. Galectin-3 is widely involved in tumor cell migration, adhesion, differentiation, and metastasis [2, 3]. Galectin-3 has an anti-apoptotic role and high expression level in breast cancer cells [4]. Although the precise role of galectin-3 remains to be determined, several studies have shown that the expression of galectin-3 is positively correlated with the metastatic potential of several tumorigenic cell lines.
However, the generality of these findings about epithelial cell-derived human tumors is unclear, as increases and decreases galectin-3 have been reported during the malignant progression of several cancers [5]. Thus, its regulation is critical as an underlying pathway in estrogen-responsive cells when exposed to estrogens or anti-estrogens.
Therefore, we have studied the expression of galectin-3 in MCF-7 cells following treatment with 17β-estradiol (E2) and tamoxifen (TAM).
In addition, we have tested laminin expression in MCF-7 cells under the same treatment regimen because cell-matrix interaction is considered one of the most significant advances in cancer research [6]. The non-cellular extra-cellular matrix (ECM) component, laminin, is a complex structure involved in cellular adhesion and differentiation in breast tumor development by influencing the surrounding cells. Moreover, the interactions between intracellular protein, galectin-3, and ECM protein laminin are possible through their binding cascade and overall cytoskeletal architecture. This interaction mechanism facilitates signaling pathways. Therefore, we tested both galectin-3 and laminin as biochemical markers in this study.
It has long been recognized that tamoxifen can bind to the ER receptors, thereby acting as a competitive inhibitor to estradiol [7, 8]. It attaches ERs to tumors and other tissue targets, producing a nuclear complex that decreases DNA synthesis and initial estrogen effects [9]. Thus, tamoxifen reduces metastasis and lowers the risk of death from breast cancer [10]. One mechanism in which anti-estrogens such as tamoxifen are believed to function is by holding the dividing cell in the G1 phase of the cell cycle, thereby decreasing cell proliferation [11, 12]. The role of galectin-3 protein in cells in programming cell death in response to tamoxifen or other hormonal treatments is unknown. In one 72-hour analysis, it was determined that for MCF-7 cells, mRNA and protein levels associated with ER expression were slightly elevated or unregulated with very low concentrations of 10-10M and 10-11M estradiol. In contrast, higher concentrations reduced mRNA and protein expression over the 72-hour study period [13].
This study examined the effects of tamoxifen and its counterpart estradiol over 24-72 hours of exposure. We tested their results one day earlier, and the nature of these effects on the MCF-7 cell cycle regarding anti-proliferative and growth suppressive response to tamoxifen with a concomitant change in galectin-3 or ECM component laminin. Currently, there is no report examining the relevance of galectin-3 or laminin to the short-term treatment of tamoxifen in ER-positive breast cancer cells. Our study demonstrates that tamoxifen-induced growth suppression and death of MCF-7 breast cancer cells are associated with galectin-3 downregulation while laminin levels remain unaltered.
A mechanistic model depicting galectin-3 downregulation due to tamoxifen-induced cellular changes.
Materials and Methods
Cell Culture
MCF-7 human mammary adenocarcinoma cell line was obtained from the American Type Culture Collection (Rockville, MD) and routinely cultured in Dulbecco’s Modified Eagle’s/ F-12 media (DMEM; Sigma Chemical Co., St. Louis, MO) with 10% fetal bovine serum (FBS) in a 5% CO2 incubator at 37°C. After trypsinizing and counting, all cells were maintained in phenol red-free growth medium supplemented with 10% charcoal-stripped FBS to evaluate the effects of the treatments.
For western blotting, cells (1.0 × 106 cells) were cultured on eight 60 mm plates for 24 hours in phenol red-free medium supplemented with 10% charcoal-stripped FBS at 37°C in 5% CO2 and cultured in the same medium for 24 to 72 hours in the presence and absence of the treatments as mentioned below.
5 cells were plated in duplicate on eight 35mm plates. Typically, after 24 hours of incubation, growth media was replaced with 10% charcoal- striped FBS-containing phenol red-free medium (control), media containing 0.01% DMSO (as vehicle control), or media containing E2 (10 For flow cytometry, 5.0 x 10 –8 M), and TAM (10 -6 M, 10 –8 M). We have monitored the cell culture conditions routinely under a Nikon Phase-contrast microscope.
–6 M, 10
MTT Assay
Cell viability was determined with the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay [14]. This assay followed instructions in the MTT assay kit from Sigma-Aldrich (St. Louis, MO, USA).
M-PER Protein Extraction
After 24-72 hours of in vitro exposure to the MCF-7 breast cancer cell line, the media were removed from the culture dish, and cells were thoroughly washed with 1X phosphate buffer saline (PBS). 1X HALT protease inhibitor cocktail was added to the mammalian protein extraction reagent (MPER), and 200µl of the cocktail was added to the culture dish, which was then placed on ice. After 20 minutes, the cells were centrifuged for 10 minutes at 200 x g. The supernatant was collected in microfuge tubes, and the solubilized protein samples were stored at -80ºC.
Protein Assay
Cells were removed from the culture dishes, rinsed with PBS, and collected into plastic tubes. The pooled cells underwent sonication and were then collected in PBS by centrifugation (@ 200 x g for 5 min). Protein samples were stored at -80°C until assayed. The BCA protein assay kit was purchased from Pierce Chemical Company (Rockford, Ill) and used to determine cell protein concentrations as described earlier [15].
ELISA
Fifty microliters of protein samples were added to each well of a 96-well culture plate and incubated overnight. We washed three times with a wash buffer before blocking for 1 hour. Then, we added 50 µl of rabbit polyclonal galectin-3 primary antibody (1:500) and incubated for 1 hour. After washing multiple times, we added goat anti-rabbit secondary antibody (1:5,000) and incubated for 1 hour. We repeated the washing steps numerous times before adding 100 µl/ well of HRP-substrate solution (Bio-Rad Laboratories, Richmond, CA) for 1 hour. Finally, we stopped the reaction by adding 100 µl of 5% SDS solution for 30 min. The wells were read at 415 nm wavelength using the iMark Bio-Rad plate reader. All antibodies from Invitrogen were purchased from ThermoFisher Scientific, USA.
Flow Cytometry
5 cells were plated in duplicate on 35mm dishes. After 48 hours of incubation, cells were briefly trypsinized (trypsin-EDTA) and centrifuged at 3500 rpm for five minutes. The cell pellet from each treatment group was fixed in cold ethanol for 15 min and stained with 1mL Propidium Iodide (PI) solution (10 mM Tris-HCl, 10 µg/mL RNase, 10mM NaCl, and 0.1 mg PI/mL and 2.0% NP-40) before analyzing the DNA ploidy distributions at 488 nm wavelength using a Beckman Coulter FACScan flow- cytometer equipped with a 15mW air-cool laser. Nuclear DNA content was examined based on the procedure as described earlier [16].
For flow cytometry, 5.0 x 10
Fluorescence Microscopy
Cells were grown for 48 hours on a square 22 x 22 mm coverglass in the respective treatments. Cells were then washed in phosphate-buffered saline (PBS), pH 7.4, and were fixed in 3.0% paraformaldehyde solution for 10 minutes, followed by washing twice in PBS containing 0.1% Tween-20. Cells were then incubated for 5 minutes at room temperature with 5% bovine serum albumin in PBS with 0.1% Triton-X-100. Five microliters of galectin-3 rabbit polyclonal antibody were dissolved into 200µl of PBS for each coverslip for 30 minutes at room temperature. This is followed by staining with FITC-conjugated goat anti- rabbit secondary antibody for 30 min. During incubation, coverslips were kept in a covered container to prevent evaporation. The stained cells were washed twice with PBS, mounted with Prolong antifade mounting medium (Molecular Probes, Eugene, OR), and viewed under a Zeiss fluorescence microscope. All primary and secondary antibodies from Invitrogen were purchased from ThermoFisher Scientific, USA.
Gel Electrophoresis and Western Blot Analysis
Twenty microliters of protein samples from each treatment were loaded in each lane. The proteins were separated by 10 % SDS-PAGE at 100 volts (V) for about an hour, after which the gel was removed. Proteins from the gel were transferred at 200 V to Hybond-ECL nitrocellulose membranes (Pharmacia Biotech, Buckinghamshire, UK) and blocked for 1 hour at 4˚C in 1% Blotto non-fat dry milk (Santa Cruz Biotechnology, Inc., Santa Cruz, CA).
Membranes were incubated with 0.5 µg/ml rabbit polyclonal galectin-3 or laminin primary antibody for 1 hr at room temperature and subsequently in 1 µg/ml peroxidase-conjugated anti-rabbit IgG (Fab specific) at room temperature. ECL western blotting detection analysis (Amersham Biosciences, Buckinghamshire, UK) was used to visualize protein expression on X-ray film (Eastman Kodak Co., Rochester, NY). All antibodies from Invitrogen were
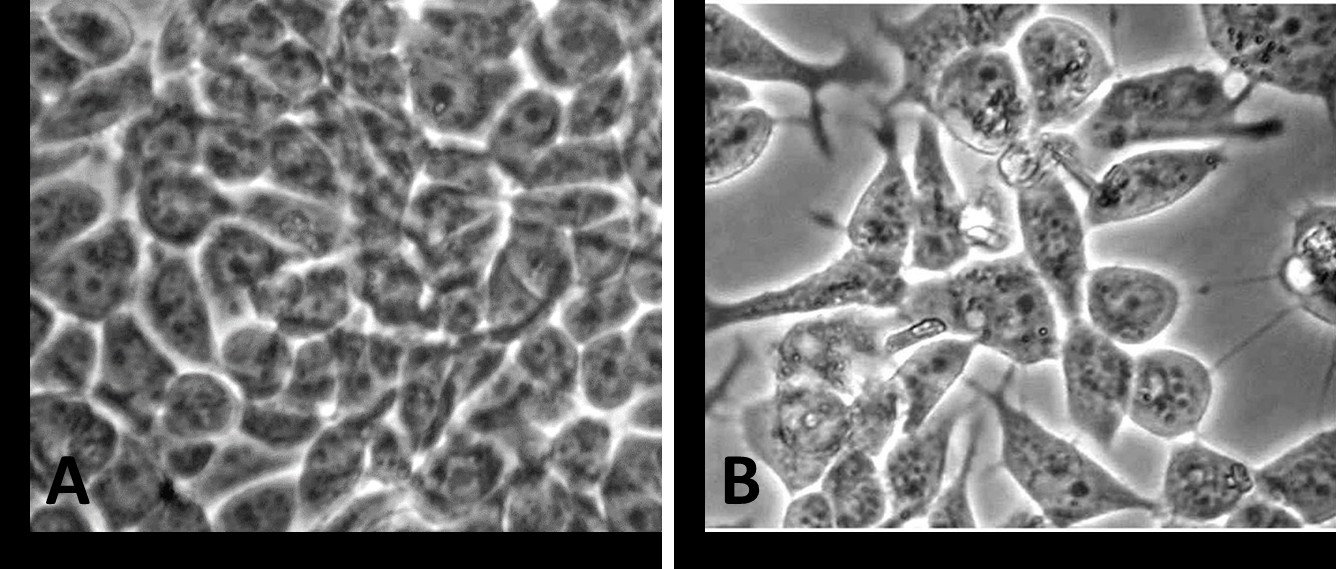
Figure 1: Phase-contrast photomicrograph of MCF-7 cells incubated in medium containing 0.01% DMSO as vehicle control (A) or in medium with the vehicle containing 10-8M tamoxifen - TAM (B). Microscopic examination revealed an alteration in shape and prominent cytoplasmic extensions in MCF-7 cells when treated with tamoxifen for 48 hours (A- Vehicle control versus B- 10-8M TAM).
Effects of Tamoxifen on Cell Viability (A) And Relative Levels of Galectin-3 (B) and Laminin (C)
purchased from ThermoFisher Scientific, USA.
Statistical Analysis
Experiments were repeated three times with two dishes per treatment, and the mean and standard error (SEM) were calculated. Differences between the various treatment groups were analyzed by one-way repeated measures analysis of variance. Multiple comparison tests analyzed the groups with significant differences (p<0.05). Prism 9.02 software (Graphpad Inc., San Diego, CA) was utilized to analyze the data.
Results
Effects of Tamoxifen on Cell Morphology
Representative phase-contrast photomicrographs of cultured MCF-7 cells are presented in (Figure 1). MCF- 7 cells incubated for 48 h in 0.01% DMSO (A) appeared morphologically normal in culture (Figure 1A). Cell-cell contacts remained undisturbed. In contrast, treatments with 10-8 M (Figure 1B) altered cellular morphology. In particular, MCF-7 cells were retracted and shrunken with prominent cytoplasmic extensions when exposed to TAM compared to the control (Figure 1A).
Table 1 (A) represents results from the cell viability assay of MCF-7 cells exposed to tamoxifen for 48 h. A significant and dose-dependent decrease in cell viability was observed with
10-6M (~50% reduction) or 10-8M TAM (~20% reduction) (P < 0.05) compared to the DMSO control. The ELISA data from a single 48-hour exposure in Table 1 (B) revealed that TAM at 10-8 M and 10-6 M concentrations decreased the relative galectin-3 levels (P < 0.05). Tamoxifen did not reduce laminin levels (C), but as expected, E2 moderately increased (~15% increase) laminin.
| Treatment | Percent viable cells (A) | Percent gal-3 (B) | Percent LN (C) |
|---|---|---|---|
| Media | 96.2 ± 4.3 | 100 | 100 |
| 0.01% DMSO | 98.7 ± 4.7 | 90.6 ± 5.8 | 100 |
| 10-6 M E2 | 98.8 ± 4.8 | 110.0 ± 5.7 | 116 ± 6.5 |
| 10-8 M E2 | 96.5 ± 4.2 | 90.3 ± 4.6 | 114 ± 5.2 |
| 10-6 M TAM | 52.5 ± 5.4* | 54.7 ± 3.2* | 95.4 ± 5.8 |
| 10-8 M TAM | 76.9 ± 6.1* | 59.5 ± 2.0* | 99.0 ± 6.5 |
Table 1: Effects of a single 48-hour exposure on the viability of cultured MCF-7 cells (A) and relative galectin-3 (gal-3, B) and
Data represent the mean and SEM from 3 separate experiments. * denotes a statistical and dose-dependent decrease (p<0.05) in cell viability (A) with both 10-6M and 10-8M tamoxifen (TAM) when compared to the Media, DMSO controls, or both 17β-estradiol (E2) exposures. The relative galectin-3 (gal-3) levels (B) were decreased (p<0.05) by both 10-6M and 10-8M tamoxifen (TAM) exposures (asterisks), whereas the relative laminin levels (C) did not change significantly.
Effect of Tamoxifen on the Cell Cycle
Flow cytometric analysis of the cell cycle data, showing a single 48-hour exposure, is presented in Table 2. TAM at 10-6M significantly (P < 0.05) reduced DNA in all phases, including the S phase, of MCF-7 cells compared to the DMSO control. TAM at 10-8M significantly decreased (P < 0.05) G2/M phase.
| Treatment | Phases Of Cell Cycle | G /M 2 | |
|---|---|---|---|
| G /G 0 1 | S | ||
| Media | 67.3+2.4 | 11.7+0.7 | 19.7+1.1 |
| 0.01% DMSO | 63.6+1.5 | 14.5 +1.5 | 20.3+1.4 |
| 10-6 M E2 | 76.7+4.7 | 15.7 + 1.7 | 20.2+1.6 |
| 10-8 M E2 | 75.5+4.8 | 16.4+0.6 | 17.1+1.4 |
| 10-6 M TAM | 18.8 + 1.0* | 6.1+0.6* | 9.8+0.3* |
| 10-8 M TAM | 59.7+2.2 | 10.7+0.6 | 11.1+1.3* |
Table 2: Effects of a single 48-hour exposure on flow-cytometric cell cycle analysis of cultured MCF-7 cells.
Data represent the mean and SEM from 3 separate experiments. * denotes statistical significance (p<0.05) compared to Media, DMSO controls, or both E2 exposures.
Immunofluorescence Microscopy
The cellular localization of galectin-3 was established by immunostaining both control and experimental (E2 and TAM-exposed) cells using anti-galectin-3 antibodies (Figure 2). Cytoplasmic and nuclear localization of galectin-3 was evident in the control groups. With E2 10-6M and 10-8M exposures, the staining was highly visualized both on the surface and in the nucleus of MCF-7 cells.
However, in 48-hour treatment with TAM 10-6M, the staining of galectin-3 was greatly diminished. Control labeling of the cells with only secondary antibodies was negative (data not shown).
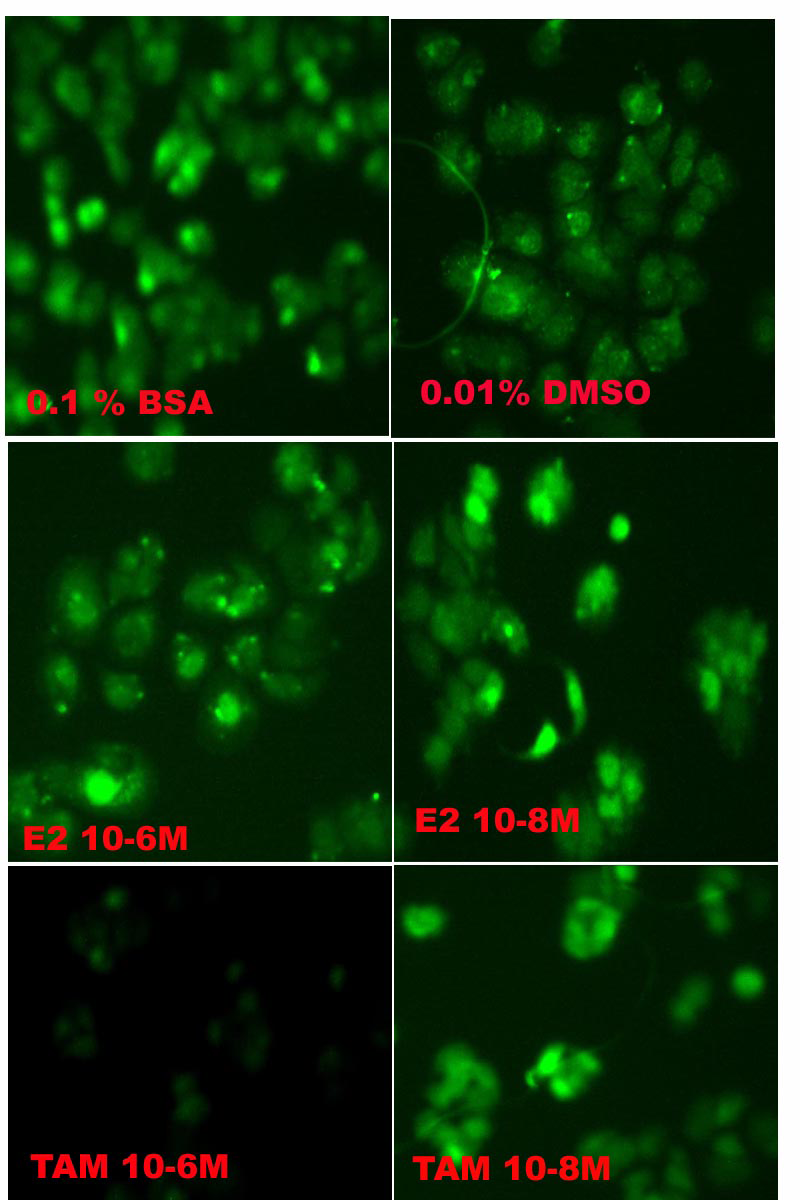
Figure 2: Photomicrograph of fluorescent-labeled galectin-3 localization in cultured MCF-7 cells. MCF-7 cells were cultured on coverslips and incubated for an additional 48 hours in a medium containing various exposures as described in the Materials and Methods. The cells treated with 10-6M tamoxifen (10-6M TAM) exhibited a shorter size with decreased galectin-3 labeling.
Galectin-3 and Laminin Immunolocalization (Western Blot)
The expression levels of galectin-3 or ECM protein laminin were determined by Western blot analysis (Figure 3). We used β-actin as a housekeeping protein for equal loading and transfer. The immunoblot analysis revealed no relative change in galectin-3 expression in the 24-hour treatment group (Figure 3A). However, in the 48-hour treatment group, the relative expression of the galectin-3 protein was significantly decreased with 10-6M TAM (Figure 3B). We have also observed a decrease in galectin-3 with 10-6M TAM in the 72-hour treatment group (Figure 3C). In comparison, the relative expression of laminin in the 48 h treatment group remained unaltered with 10-6M TAM (Figure 3D). However, we have observed an increase in laminin with both 10-6M and 10-8M E2.
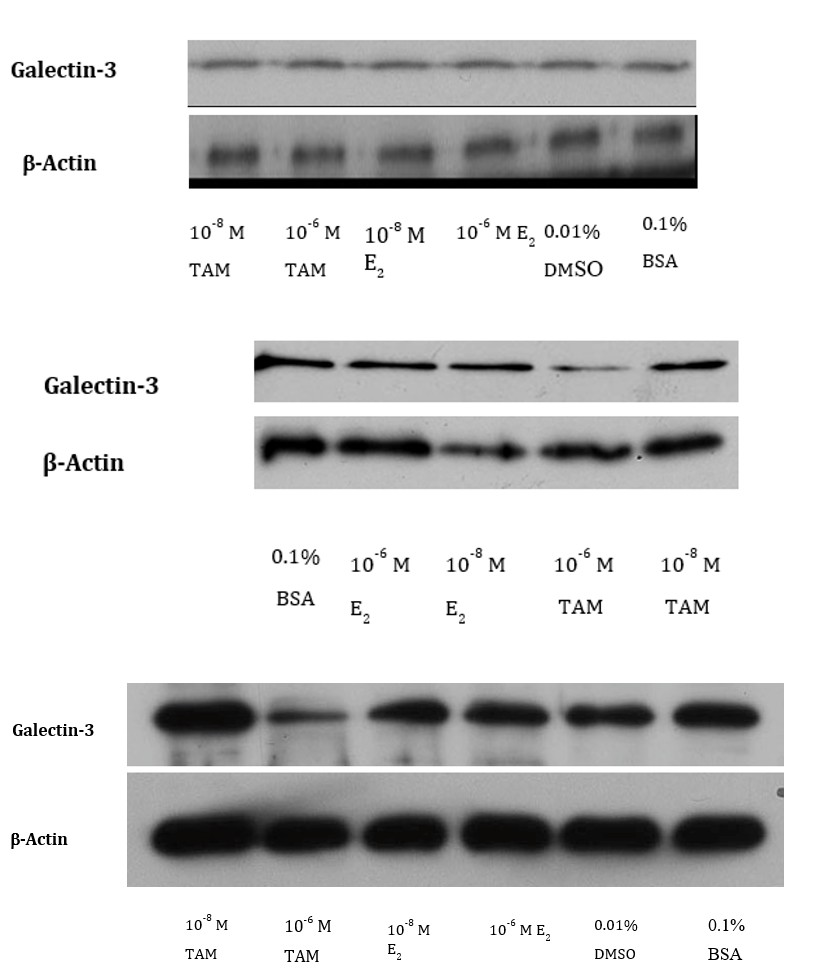
Figure 3A: Western blot of galectin-3 in MCF-7 Cells - 24 hour Treatment.
Quantification of Western blot data in terms of ratio (galectin-3/β-actin)
| Exposure | 0.1% BSA | 0.01% DMSO | 10-6M E2 | 10-8M E2 | 10-6MTAM | 10-8M TAM |
|---|---|---|---|---|---|---|
| 24h | 0.40 ± 0.002 | 0.40±0.001 | 0.35±0.0001 | 0.38±0.003 | 0.42±0.008 | 0.40±0.0004 |
Figure 3B: Western blot of galectin-3 in MCF-7 Cells - 48 hour Treatment.
Quantification of Western blot data in terms of ratio (galectin-3/β-actin).
| Exposure | 0.1% BSA | 0.01% DMSO | 10-6M E2 | 10-8M E2 | 10-6MTAM | 10-8M TAM |
|---|---|---|---|---|---|---|
| 48h | 0.50 ± 0.002 | ND | 0.65±0.0001 | 0.60±0.003 | 0.15±0.003 | 0.35±0.0002 |
Figure: 3C: Western blot of galectin-3 in MCF-7 Cells - 72 hours Treatment.
Quantification of Western blot data in terms of ratio (galectin-3/β-actin).
| Exposure | 0.1% BSA | 0.01% DMSO | 10-6M E2 | 10-8M E2 | 10-6MTAM | 10-8M TAM |
|---|---|---|---|---|---|---|
| 72h | 0.40 ± 0.001 | 0.35±0.002 | 0.40±0.002 | 0.35±0.002 | 0.15±0.003 | 0.45±0.004 |
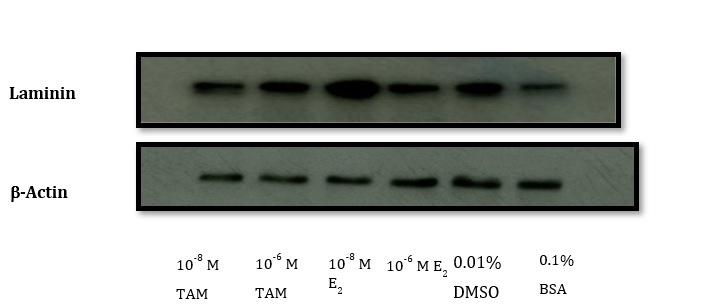
Figure 3D: Western blot of laminin in MCF-7 Cells - 48 hour Treatment.
Quantification of Western blot data in terms of ratio (laminin/β-actin)
| Exposure | 0.1% BSA | 0.01% DMSO | 10-6M E2 | 10-8M E2 | 10-6MTAM | 10-8M TAM |
|---|---|---|---|---|---|---|
| 48h | 0.50 ± 0.002 | 0.60±0.001 | 0.80±0.0001 | 1.0±0.003 | 0.60±0.008 | 0.50±0.0004 |
Discussion
For over three decades, Tamoxifen has been widely accepted as an adjuvant endocrine drug. It can bind to ERα and act as an antagonist through competition with estradiol. It modulates gene expression in ERα-positive breast cancer cells [17, 18]. In the present study, we demonstrated dose- dependent morphological changes in MCF-7 cells exposed to tamoxifen. Earlier, we reported that relatively higher TAM concentrations (10-4M) induced morphological changes and reduced DNA in MCF-7 cells when exposed for 24 hours [19]. Others observed morphological changes in apoptosis or PCD, which is cell condensation and the reorganization of the cell cytoskeleton [20]. We also noted earlier a reduced level of F-action expression in MCF-7 cells with 10-4M of TAM exposure in 24 hours [19]. Cell preparation and execution of apoptosis are associated with depolymerization and cleavage of actin and other cytoskeletal proteins [21, 22, 23, 24, 25, 26, 27, 28, 29, 30]. However, the role of galectin-3 and its association with the ECM protein laminin was not reported in earlier studies.
Western blot analysis was utilized to determine the effect of tamoxifen on galectin-3 and laminin expressions in MCF-7 breast cancer cells. Over the 24-hour incubation period, no significant changes were observed. However, in 48- and 72-hour periods, TAM (10-6M) treatment decreased galectin-3 expression in MCF-7 cells compared to the controls, and laminin levels remained unaffected. Even with the morphological changes due to the exposure of MCF-7 cells to tamoxifen, the ECM protein laminin didn’t show any alterations. Still, the cellular protein galectin-3 demonstrated marked reductions beyond the 24-hour exposure period. Laminin, a key extracellular matrix component, plays a critical role in maintaining cellular adhesion, polarity, and cell-matrix interactions. The preservation of laminin expression suggests that tamoxifen may not broadly disrupt cell-ECM interactions, underscoring a nuanced mechanism through which tamoxifen rather selectively modulates intracellular functions. Interestingly, this finding raises questions about the specificity of tamoxifen and how it modulates cell functions without compromising the architecture of the ECM, affecting the structural and signaling components.
Here, we see a differential relationship between tamoxifen [31] and estrogens [32, 33, 34, 35, 36] regarding the inhibitory effects on cell proliferation of MCF-7 growth. Therefore, the downregulation of galectin-3 may be instrumental in cell growth inhibition or even cell death caused by tamoxifen. Earlier, at relatively higher concentrations, tamoxifen- induced apoptosis in MCF-7 cells was reported by our group [37].
We have demonstrated here that tamoxifen decreases in all phases, including the S-phase, resulting in reductions in DNA synthesis in MCF-7 cells. Tamoxifen has been known to be an inducer of apoptosis in a dose-dependent manner [38]. It is also known to appear anti-proliferative in its induction of MCF-7 growth at higher concentrations of administration [39]. In this study, tamoxifen exposure for extended periods was associated with an apparent inhibition of galectin-3 protein in cells without affecting the ECM component, such as laminin. This is the first report to demonstrate an association of galectin-3 without the involvement of laminin with the transformed phenotype of MCF-7 cells due to tamoxifen treatment. It was reported that galectin-3 was a differentially expressed and downregulated gene in LCC9 cells. However, it was normalized with the therapy of Valproic acid and hydralazine. They also found animal models using in-utero ethinyl estradiol tumors in rats. They predicted that these genes could be the reason for poor survival in TAM-treated ER-positive breast cancer patients [40].
In BT-549 mammary epithelial cells, the introduction of recombinant galectin-3 resulted in tumorigenic properties [41]. This research group later reported [42] that the downregulation of galectin-3 suppressed the tumorigenicity of highly malignant triple-negative MDA-MB-231 breast cancer cells.
In summary, tamoxifen exhibited cytostatic effects on MCF-7 breast cancer cells. Morphological analysis revealed that TAM caused MCF-7 cell shrinkage and cytoplasmic extension, suggesting a loss of epithelial integrity. Correspondingly, cell viability was significantly reduced. Tamoxifen exposure also significantly reduced S-phase DNA content, indicating cell cycle arrest and reduced proliferative capacity. Notably, galectin-3 protein levels were decreased considerably after longer TAM treatment, while laminin expression remained unchanged, implying selective modulation of intracellular signaling and extracellular matrix components. These results suggest that TAM impairs breast cancer cell survival by altering morphology, inhibiting proliferation, and downregulating galectin-3, a multifunctional protein implicated in pathophysiology.
Acknowledgement
Part of the support was through the National Institutes of Health grant number GM 103499. Students presented part of the research in the Ernest E Just Science Symposium, Medical University of South Carolina, USA.
Conflicts of Interest
The authors declare no conflict of interest.
References
-
Barondes SH, Cooper DN, Gitt MA, Leffler H (1994) Galectins. Structure and function of a large family of animal lectins. J Biol Chem 269: 20807-20810.
-
Liu FT, Rabinovich GA (2005) Galectins as modulators of tumour progression. Nat Rev Cancer 5: 29-41.
-
Li LC, Li J, Gao J (2014) Functions of galectin-3 and its role in fibrotic diseases. J Pharmacol Experimental Therapeutics 351: 336-343.
-
Zhang H, Luo M, Liang X, Liu C, Zhao Z (2014) Galectin-3 as a marker and potential therapeutic target in breast cancer. PloS one 9: e103482.
-
Nangia Makker P, Akahani S, Bresalier R (2000) The role of galectin-3 in tumor metastasis. In: Caron M, Steve AP (Eds.), Lectins and Pathology. Harwood Academic Publishers, Warsaw, Poland.
-
Bahcecioglu G, Basara G, Ellis BW, Ren X, Zorlutuna P (2020) Breast cancer models: engineering the tumor microenvironment. Acta Biomater 106: 1-21.
-
Skidmore JR, Walpole AL, Woodburn J (1972) Effect of some triphenylethylenes on estradiol binding in vitro to macromolecules from the uterus and anterior pituitary. J Endocrinol 52: 289-298.
-
Jordan VC, Prestwich G (1977) Binding of [3H] tamoxifen in rat uterine cytosols: A comparison of swinging bucket and vertical tube rotor sucrose density gradient analysis. Mol Cell Endocrinol 8: 179-188.
-
Dhingra K (1999) Antiestrogens--tamoxifen, SERMs, and beyond_._ Investigational New Drugs 17: 285-311.
-
Jordan VC, O Malley BW (2007) Selective estrogen- receptor modulators and antihormonal resistance in breast cancer. J Clin Oncol 25: 5815-5824.
-
Sutherland RL, Hall RE, Taylor IW (1983) Cell proliferation kinetics of MCF-7 human mammary carcinoma cells in culture and effects of tamoxifen on exponentially growing and plateau-phase cells. Cancer Res 143: 3993-4006.
-
Osborne CK, Bolt DH, Estrada P (1984) Human breast cancer cell cycle synchronization by estrogens and anti- estrogens in culture. Cancer Res 44: 1433-1439.
-
Ree AH, Landmark BF, Eskild W, Levy Fo, Lahooti H, et al. (1989) Autologous down-regulation of messenger ribonucleic acid and protein levels for estrogen receptors in MCF-7 cells: an inverse correlation to progesterone receptor levels. Endocrinology 124(5): 2577-2583.
-
Raychoudhury SS, Kubinski D (2003) Polycyclic aromatic hydrocarbon-induced cytotoxicity in cultured rat Sertoli cells involves differential apoptotic response. Environ Health Perspectives 111: 33-38.
-
Raychoudhury SS, Flowers AF, Millette CF, Finlay Mf (2000) Toxic effects of polychlorinated biphenyls on cultured rat Sertoli cells. J Andrology 21: 964-973.
-
Raychoudhury SS, Blake C, Millette CF (1999) Toxic effects of octylphenol on cultured rat spermatogenic cells and Sertoli cells. Toxicol Applied Pharmacology 157: 192-202.
-
Burstein HJ, Lacchetti C, Anderson H, Buchholz TA, Davidson NE, et al. (2019) Adjuvant endocrine therapy for women with hormone receptor-positive breast cancer: ASCO clinical practice guideline focused update. J Clin Oncol 37: 423-38.
-
Cuzick J, Sestak I, Cawthorn S, Hamed H, Holli K, et al. (2015) Tamoxifen for prevention of breast cancer: extended long-term follow up of the IBIS-I breast cancer prevention trial. The Lancet Oncol 16: 67-75.
-
Kangaloo K, Jenkins L, Clarke N, Raychoudhury S (2007) The effects of 17β-estradiol and tamoxifen on cell cycle distribution and F-actin expression in MCF-7 Cells. J South Carolina Academy of Science 4(1): 1-12.
-
Tenniswood MP (1992) Active cell death in hormone- dependent tissues. Cancer Metastasis Rev 11: 197-220.
-
Bonfoco E, Leist M, Zhivotovsky B, Orrenius S, Lipton SA, et al. (1996) Cytoskeletal breakdown and apoptosis elicited by NO donors in cerebellar granule cells require NMDA receptor activation. J Neurochem 67: 248-249.
-
Chen Z, Naito M, Mashima T, Tsuruo T (1996) Activation of actin-cleavage interleukin 1 beta-converting enzyme (ICE) family protease CPP-32 during chemotherapeutic agent-induced apoptosis in ovarian carcinoma cells. Cancer Res 56: 5224-5229.
-
Kayalar C, Ord T, Testa MP, Zhong LT, Bredesen DE (1996) Cleavage of actin by interleukin 1β-converting enzyme to reverse DNase inhibition 93: 2234-2238.
-
Levee MG, Dabrawska MI, Lelli JL, Hinshaw DB (1996) Actin polymerization and depolymerization during apoptosis in HL-60 cells. Am J Physiol 271: 1981-1992.
-
Brancholini C, Lazarevic D, Rodriguez J, Schneider C (1997) Dismantling cell-cell contacts during apoptosis is coupled to a caspase-dependent proteolytic cleavage of beta-catenin. J Cell Biol 139: 759-771.
-
Brown SB, Bailey K, Savill J (1997) Actin is cleaved during constitutive apoptosis. Biochem J 323: 233-237.
-
Caulin C, Salvesen GS, Oshima RG (1997) Caspase cleavage of keratin 18 and reorganization of intermediate filaments during epithelial cell apoptosis. J Cell Biol 138: 1379-1394.
-
Kothakota S, Azuma T, Reinhard C, Klippel A, Tang J, et al. (1997) Caspase-3 generated fragment of gelosin: effector of morphological change in apoptosis. Science 278(5336): 294-298.
-
Ku NO, Liao J, Omary MB (1997) Apoptosis generates stable fragments of human type I keratins. J Biol Chem 272: 33197-33203.
-
Kruiudering M, van de Water B, Zhan Y, Baelde JJ, Heer Ede, et al. (1998) Cisplatin effects on F-actin and matrix proteins precede renal tubular cell detachment and apoptosis in vitro. Cell Death and Differentiation 5: 601- 614.
-
Porter AG, Janicke RU (1999) Emerging roles of caspases-3 in apoptosis. Cell Death and Differentiation 6: 99-104.
-
Schmeiser K, Grand JA (1999) The fate of E- and P-cadherin during the early stages of apoptosis. Cell Death and Differentiation 6: 377-386.
-
Horwitz KB, McGuire WL (1978) Antiestrogens: mechanisms of action and effects in breast cancer. In: McGuire WL (Ed.), Breast Cancer: Advances in Research and Treatment. Plenum Publishing Corp, New York, pp: 155-204.
-
Lippman M (1981) Hormonal regulation of human breast cancer cells in vitro. Banbury Report 8: 171-181.
-
Coezy E, Borgna JL, Rochefort H (1982) Tamoxifen and estrogen receptor and inhibition of cell growth. Cancer Res 42: 317-323.
-
Darbe P, Yates J, Curtis S, King RJB (1983) Effect of estradiol on breast cancer cells in culture. Cancer Res 43: 349-354.
-
Ogba N, Wang C, Raychoudhury S (2005) Differential effects of fluoranthene and benzo[a]pyrene in MCF-7 cells. J of Environ Sci and Health Part A 40: 927-936.
-
Ercoli A, Scambia G, Fattorossi A, Raspaglio G, Battaglia A, et al. (1998) Comparative study on the induction of cytostasis and apoptosis by ICI 182,780 and tamoxifen in an estrogen receptor-negative ovarian cancer cell line. Int J Cancer 76: 47-54.
-
Safe SH (1998) Interactions between hormones and chemicals in breast cancer. Ann Rev Pharmacol Toxicol 38: 121-158.
-
Hilakivi Clarke L, Warri A, Bouker KB, Zhang X, Cook KL, et al. (2017) Effects of In Utero Exposure to Ethinyl Estradiol on Tamoxifen Resistance and Breast Cancer Recurrence in. J Natl Cancer Inst 109(1): 188.
-
Nangia Makker P, Thompson E, Hogan C, Ochieng J, Raz A (1995) Induction of tumorigenicity by galectin-3 in a non-tumorigenic human breast carcinoma cell line. Int J Oncol 7: 1079-1087.
-
Honjo Y, Nangia Makker P, Inohara H, Raz A (2001) Down- regulation of galectin-3 suppresses tumorigenicity of human breast carcinoma cells. Clin Cancer Res 7: 661- 668.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial