Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
Acrosome reaction (AR) is a major controlling step in the mammalian fertilization. Failure or improper AR is one of the reasons of infertility. Numerous unresolved questions remain regarding signaling events and biochemical mechanisms promoting the membrane fusion of the mammalian sperm AR. Previously, we characterized an acrosomal matrix component from the acrosome named the outer acrosomal membrane-associated matrix complex (OMC), consist of three major (54, 50, and 45kDa) and several minor (38-19kDa) polypeptides, termed “rpf”. A 32kDa polypeptide (OMC32) was characterized from the “rpf” polypeptides. The present study demonstrates that OMC32 is present in one large complex within the acrosomal matrix and there is a physiological interaction between OMC 32 and IZUMO1/SPACA3 polypeptides of bovine sperm acrosome. We have shown that there is a significant release of SPACA3 during lysophosphatidyl choline-induced acrosome reaction and SPACA3 was not released in the presence of acrosin inhibitors. This study reveals that acrosin involves in the release of SPACA3 during acrosome exocytosis. To describe the function of OMC32 polypeptide, Fab fragments prepared from the IgG fractions of anti- OMC32 were utilized and the inhibition of acrosomal exocytosis was examined using the release of acrosin as a potential marker. The inhibition of acrosin release was observed in the presence of 10μg Fab fragments. Our established permeabilizedcell protocol for exocytosis strongly reveals that OMC32 plays a significant role in the regulation of acrosin release during acrosomal exocytosis. This study exhibits the role of acrosomal hydrolases and matrix proteins in mammalian fertilization.
Introduction
The highly polarized architecture of the mammalian spermatozoa leads to the segregation of organelles. Head and tail are the two main structures of a mammalian spermatozoon connected by a narrow neck. Fertilization, the unification of male and female gametes, is an essential biological event that depends on numerous coordinated biochemical and physiological events [1]. After leaving the testis, sperm must undergo several physiological events to acquire fertilization competency that include posttesticular maturation in the epididymis and the capacitation in the microenvironment of the female reproductive tract. These physiological steps involve several proteins, signaling pathways, cell-cell contact, and cellular components. An extensive fusion between the outer acrosomal membrane and plasma membrane of anterior sperm head occurs during the acrosome reaction leading to the release of acrosomal hydrolases [2]. The mammalian sperm acrosome is a membrane-bounded organelle, comprises hydrolytic enzymes which are needed for fertilization. Some hydrolases are sorted within morphologically discrete domains of the acrosomal matrix [3, 4]. The insoluble matrix components enunciate acrosomal shape, and they have the ability in binding specific hydrolases [5, 6, 7]. It has been suggested that the interaction of acrosomal enzymes with domain-specific matrix proteins involve in stabilization of acrosomal enzyme before the acrosome reaction and control the chronological release of acrosomal enzymes during the acrosome reaction [3, 4, 6, 8]. Hence, the characterization of acrosomal proteins is requisite to comprehend the role of acrosome in fertilization. Our previous studies reveal the characterization, and the localization of a stable acrosomal matrix component of the bovine sperm acrosome termed the outer acrosomal membrane-associated matrix complex (OMC) [6, 9]. OMC fraction consists of three major (54, 50, and 45kDa) and several minor (38-19kDa) polypeptides. We termed the set of minor polypeptides (38-19kDa) “rpf”, solubilized by high pH solubilization, and other three major polypeptides (55, 50 and 45kDa) stayed with the sedimentable “stripped” OMC [10]. A 32kDa polypeptide isolated from high pH soluble fraction of OMC (OMC32) showed N-terminus amino acid sequence homology of SP-10 proteins of baboon [11], human [12], mouse [13], and fox [14] spermatozoa. Our proteomic analysis of the OMC32 polypeptide by MALDI-TOF-TOF revealed the acrosin-binding protein and OMC32 polypeptide binds to acrosin, lactadherin, SPACA3, and IZUMO1 [15]. We propose that the interaction of OMC32 with detergent soluble acrosomal proteins is a receptor-ligand type of binding. Numerous unresolved questions remain regarding signaling events and biochemical mechanisms promoting the mammalian sperm acrosomal exocytosis. Does the interaction of OMC32 to other acrosomal proteins regulate the hydrolases release and other acrosomal proteins during the acrosome reaction? Is acrosin involved in the release of acrosomal proteins during the acrosome reaction? In the present study, we examined whether OMC32 exist in one large complex within the acrosomal matrix and the role of OMC32 polypeptide in acrosome reaction. In addition, we also examined whether acrosin plays an important role in the release of acrosomal proteins during acrosomal exocytosis.
Materials and Methods
Sperm Preparation
Bovine epididymides were obtained from Randloph Packing, Asheboro, North Carolina, maintaining at 4oC throughout the transport and used for sperm preparation within 30 minutes of arrival. Cauda epididymal regions were removed from the organ, minced, and incubated for 5 minutes at 37oC in Hank’s balanced saline solution, pH 7.4, having 5mM HEPES, 2 mM benzamidine, and 0.05% sodium azide. Sperm suspensions centrifuged at 100 x g for 1 minute to remove epididymal tubule fragments. Supernatants were then centrifuged at 1500 x g for 10 minutes at 4oC using an Eppendorf Centrifuge. Pellets were washed 3 times by resuspension in Hanks balanced saline solution, as stated above. Then the sperm pellet resuspended in a Tris-saline- protease inhibitor solution (TNI) containing 150 mM NaCl, 25 mM Tris-HCl, (pH 7.5), 2 mM benzamidine, 1 µg/mL leupeptin, 1 µg/mL pepstatin, and 0.05% sodium azide and centrifuged at 1500 x g for 10 minutes at 4oC. Pellets were washed 2 more times in TNI as stated above.
SDS-PAGE, Western Blotting, and Immunofluorescence Microscopy
SDS-PAGE was executed on 12% or 4-20% gradient gel (Bio-Rad) polyacrylamide gels [16]. Western Blot analysis was accomplished by the electrophoretic transfer of polypeptides to PVDF membranes [17]. Immunoblots were rinsed with TBS buffer containing 10mM Tris-HCl pH 7.5, 100 mM NaCl and 0.1% Tween-20 (TBS-0.1% Tween-20). To preclude non-specific binding, membranes were incubated in LI-COR Intercept Antibody Diluent for two hours at room temperature. Blots were then incubated with primary antibodies (1:2000 dilution) in LI-COR Intercept Antibody Diluent having 0.1% Tween-20 for two hours at room temperature. After three times (3 minutes per wash) washing with TBS-0.1% Tween-20, membranes were incubated with IRDye ® 680RD, goat anti-rabbit for LICOR Odyssey CLx Imaging System or an affinity purified horseradish peroxidase-conjugated secondary antibody (1:5000 dilution) for color reaction. Immunoreactive protein bands were detected using LICOR Odyssey CLx Imaging System or by color development using H2O2 and diaminobenzidine. Coomassie blue was used for the staining of polypeptide bands [18]. Proteins were estimated by the method of Bradford [19].
Immunofluorescence microscopy was performed following our published protocol [15]. Briefly, cauda epididymal sperm were enclosed to poly-L-Lysine smeared coverslips after fixation at 4oC in 4% formaldehyde in 0.1 M sodium phosphate buffer, pH 7.6, for 30 minutes, followed by washing in PBS, and then permeabilized was done by incubation for 10 minutes in -20oC acetone. After three washing in PBS, nonspecific protein binding sites were impeded in PBS containing 0.1% Tween-20, 5% goat serum, and 2.5% BSA (blocking solution) followed by incubation with anti-PG10 (Millipore, 1:2000 dilution) in blocking solution for 1 hour at room temperature. After washing in blocking solution, it was incubated with Cy3 conjugated goat anti-mouse IgG (KPL Inc., Gaithersburg, MD) in blocking solution for 1 hour at room temperature. After washing the coverslips with PBS and the cells were analyzed by phase contrast and epifluorescence microscopy.
Cross-Linking Experiments
High pH soluble fraction of bovine cauda epididymal sperm was obtained incubating sperm in 100 mM CAPS buffer (3-[cyclohexamino]-1-propanesulfonic acid; Sigma Chemical Co., St. Louis, MO), pH 10.5 overnight at 4°C followed by centrifugation at 100,000 X g in a Beckman SW40 rotor for 30 min [10]. The supernatant obtained after the high pH extraction was dialyzed against PBS (phosphate buffered saline, pH 7.4), then the cross-linking was performed chemically using the homobifunctional cleavable cross- linkers, dithiobis [sulfosuccinimidyl propionate] (DTSSP) following the Pierce manual. Cross-linking was concluded by the addition of 10mM glycine. Then, the supernatant was analyzed by non-reducing SDS-PAGE (4-20% gradient gel) and reducing 12% SDS-PAGE, transfer to PVDF membrane, and immunostained with anti-OMC32, anti-IZUMO1, and anti-SPACA3 antibodies or Coomassie Blue for the staining of proteins.
Capacitation and Acrosome Reaction
Sperm (4-6 X 107/ml) were capacitated following the established protocol using heparin (10 mg/ml) in a modified Tyrode’s medium (pH 7.4) for 4 hours at 39oC with a 95% air: 5% CO2 atmosphere [20]. Then the acrosome reaction was initiated by incubation with 100 μg/mL lysophosphatidyl choline (LPC) for 15 minutes. Samples were centrifuged at 4000 x g for 10 minutes at 4oC and the volume of all pellets and supernatants fractions were adjusted to equal volumes and subjected to SDS-PAGE analysis and acrosin assay. Acrosome-reacted spermatozoa were examined using fluorescent PSA lectin [21].
Acrosin Assay
Spectrophotometric assay of acrosin activity of the pellet and supernatant fractions of capacitated and acrosome reacted spermatozoa were performed at 410nm following our published procedure [5, 6]. One unit of acrosin is defined as the quantity of acrosin required to hydrolyze one micromole of N-p-Tosyl-Gly-Pro-Arg-p-niroanilide per min.
Development of a Digitonin-Permeabilized Protocol and the Inhibition of Acrosomal Exocytosis by Fab Fragments of Anti-OMC32
Permeabilization of bovine ejaculated sperm by digitonin was examined by assaying lactate dehydrogenase activity. Noncapacitated spermatozoa (5 X 107 cells) were permeabilized using various digitonin concentrations (0.001%-0.01%) or without digitonin (as a control) in 30mM triethanolamine (pH 7.8), 150mM NaCl, 1mM DTT for 15 seconds at 24oC [22]. Sperm were pelleted after centrifugation at 14,000g for 10 mins. The assay of lactate dehydrogenase (LDH) activity [23] was performed of the supernatant fractions. Noncapacitated and capacitated spermatozoa (5 X 107 cells) were permeabilized using 0.005% digitonin, for 15 seconds at 24oC. Fab fragments were isolated from IgG fractions anti-OMC32 using Pierce Fab Micro Preparation Kit and used the LPC-induced acrosome reaction for the inhibitory activities of acrosomal exocytosis. Controls were performed using Fab fragments prepared from preimmune IgG. Noncapacitated and capacitated spermatozoa after permeabilization were incubated with Fab fragments for 15 minutes; then LPC was added for the initiation of the acrosome reaction. The inhibition of acrosomal exocytosis was examined using the release of acrosin as a potential marker.
Results
OMC32 Exists in one Large Complex Within the Acrosomal Matrix
High pH supernatant of bovine cauda sperm was cross-linked using DTSSP and the cross-linked fraction was subjected to non-reducing SDS-PAGE (4-20% gradient gel), and immunostained with anti-OMC32, anti-IZUMO1, and anti-SPACA3 antibodies after transferring to PVDF membrane. A Coomassie Blue stained high molecular weight protein complex (~400kDa) exhibits in the supernatant after cross-linking under non-reducing condition (Figure. 1A, lane 2, shown by an arrow). Identical lanes immunostained with anti-OMC32, anti-IZUMO1, and anti-SPACA3 revealed that the same high molecular weight protein complex (400kDa) exhibits immunoreactive bands with OMC32 antibody (Figure 1A, lane 3), IZUMO1 antibody (Figure 1A, lane 4) and SPACA3 antibody (Figure 1A, lane 5). Next, the high molecular weight protein complex (~400kDa) was excised from the non- reducing gel and analyzed in a 12% SDS-PAGE under reducing conditions, transfer to PVDF membrane, and immunostained with anti-OMC32, anti-IZUMO1, and anti-SPACA3 antibodies (Figure 1B). The 38-19kDa related polypeptide family polypeptides was present (Figure 1B, lane 1) when stained with anti-OMC32 antibody. Identical lanes stained with anti- IZUMO1 and anti-SPACA3 antibodies exhibit the presence of IZUMO1 (Figure 1B, lane 2) and SPACA3 (Figure 1B, lane 3) immunoreactive bands, respectively. An identical lane was stained with Coomassie Blue for the staining of proteins (Figure 1B, lane 4). Our results strongly demonstrate that OMC32 and IZUMO1/SPACA3 polypeptides are present in one large complex within the acrosomal matrix.
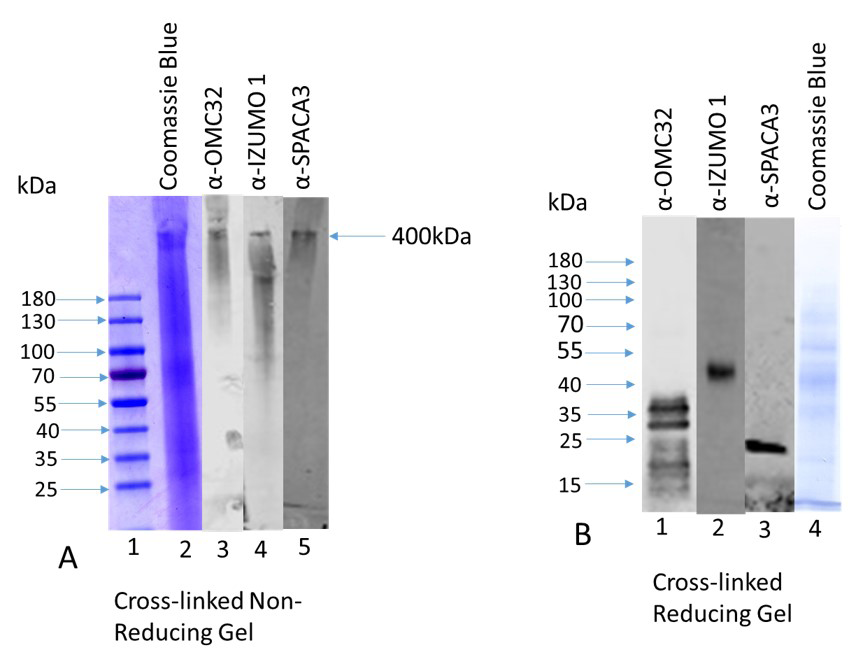
Figure 1: A: A high molecular weight protein complex (~400kDa) exhibits in the supernatant after cross-linking (25 µg protein) when fractionated in a 4-20% gradient gel under non-reducing condition and stained with Coomassie Blue (Figure A, lane 2, shown by an arrow). Identical lanes immunostained with anti-OMC32, anti-IZUMO1, and anti-SPACA3 revealed that the same high molecular weight protein complex (400kDa) exhibits immunoreactive bands with OMC32 antibody (lane 3), IZUMO1 antibody (lane 4) and SPACA3 antibody (lane 5). Lane 1 stands for the prestained protein ladder. B: The high molecular weight protein complex (~400kDa) was analyzed in a 12% SDS-PAGE under reducing conditions, transfer to PVDF membrane, and immunostained with anti-OMC32, anti-IZUMO1, and anti-SPACA3 antibodies (Figure 1B). The presence of 38-19kDa related polypeptide family polypeptides (Figure. B, lane 1) when stained with anti-OMC32 antibody. Identical lanes stained with anti-IZUMO1 and anti-SPACA3 antibodies exhibit the presence of IZUMO1 (Figure 1B, lane 2) and SPACA3 (Figure B, lane 3) immunoreactive bands, respectively. Lane 4 exhibits a Coomassie Blue stained lane under reducing conditions.
Acrosin is Involved in the Release of SPACA3 During the Acrosome Reaction
Following capacitation and LPC-induced acrosome reaction, both the supernatant and pellet fractions of all treatments were analyzed by Western blot analyses and immunostained with anti-SPACA3 antibody (Figure 2A). A 25kDa SPACA3 immunoreactive band was present in control pellet (lane1), and capacitated pellet (lane 3); no band was seen both in control supernatant (lane 2) and capacitated supernatant (lane 4). SPACA3 was released completely during LPC-induced acrosome reaction as evident the presence of a 25kDa SPACA3 immunoreactive band in the supernatant (lane 6) and no band was seen in acrosome reacted pellet (lane 5). LPC-induced acrosome reaction was performed in the presence of acrosin inhibitors (5mM benzamidine [Bz] and 1mM 4-Nitrophenyl 4-guanidinobenzoate hydrochloride [NPGB]). In the presence of 5mM benzamidine (a reversible inhibitor of acrosin), a major fraction of SPACA3 was retained in the pellet (lane7) whereas a minor fraction was released in the supernatant (lane 8). NPGB is an irreversible inhibitor of acrosin. In the presence of 1mM NPGB, SPACA3 was retained in the pellet (lane 9) and no release of SPACA3 occurred in the supernatant (lane10) fraction.
The related polypeptide family “rpf” polypeptides (38- 19kDa) where OMC32 is a major polypeptide (Figure 2B) are present in control (lane 1), capacitated (lane 2), and acrosome reacted (lane 3) pellet whereas no immunoreactive bands were seen in the control (lane 4), capacitated (lane 5), and acrosome reacted (lane 6) supernatant lanes. An immunoreactive IZUMO1 band (Figure 2C) is present in control (lane 1), capacitated (lane 2), and acrosome reacted (lane 3) pellet whereas no immunoreactive bands were seen in the control (lane 4), capacitated (lane 5), and acrosome reacted (lane 6) supernatant lanes.
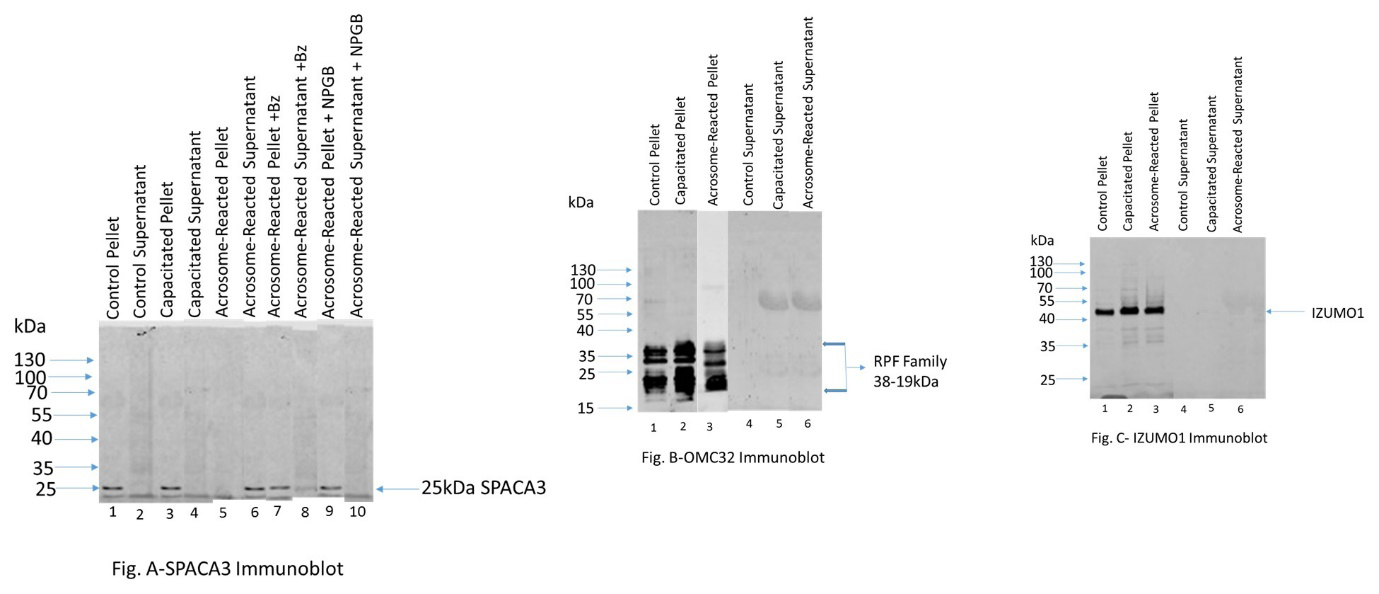
Figure 2: Figure A: 25kDa SPACA3 immunoreactive band was present in control pellet (lane1), and capacitated pellet (lane 3); no band was seen both in control supernatant (lane 2) and capacitated supernatant (lane 4). SPACA3 was released completely during LPC-induced acrosome reaction (supernatant-lane 6) and no band was seen in acrosome reacted pellet (lane 5). Benzamidine is a reversible inhibitor of acrosin. In the presence of 5mM benzamidine, a major fraction of SPACA3 was retained in the pellet (lane7) whereas a minor fraction was released in the supernatant (lane 8). NPGB is an irreversible inhibitor of acrosin. In the presence of 1mM NPGB, SPACA3 was retained in the pellet (lane 9); no release of SPACA3 occurred (supernatant-lane10). This study demonstrates that acrosin is involved in the release of SPACA3 during acrosome exocytosis. Figure B: The related polypeptide family “rpf” polypeptides (38-19kDa) where OMC32 is a major polypeptide are present in control (lane 1), capacitated (lane 2), and acrosome reacted (lane 3) pellet whereas no immunoreactive bands were seen in the control (lane 4), capacitated (lane 5), and acrosome reacted (lane 6) supernatant lanes. Figure C: An immunoreactive IZUMO1 band is present in control (lane 1), capacitated (lane 2), and acrosome reacted (lane 3) pellet whereas no immunoreactive bands were seen in the control (lane 4), capacitated (lane 5), and acrosome reacted (lane 6) supernatant lanes.
These studies reveal that SPACA3 is released during the acrosome reaction and acrosin is involved in the release of SPACA3 during acrosome exocytosis. The “rpf” polypeptides (38-19kDa; OMC32 is a major polypeptide) and IZUMO1 are not released during the LPC-induced acrosome reaction.
Role of OMC32 Polypeptide During the Acrosome Reaction Using A Permeabilized Sperm Model
Bovine cauda epididymal sperm (5 X 107 cells) was permeabilized using 0.005% digitonin following the method of Noland and Olson [22]. Permeabilization by digitonin was monitored by assaying lactate dehydrogenase activity (Figure 3A). The LDH activity in the supernatant was increased in a dose-dependent manner and it reaches at plateau in 0.005% digitonin. As a control, concentration dependent effect of digitonin on the acrosin activities was assayed (Figure 3B). Acrosin activity remained the same in control (digitonin untreated), and the different concentrations digitonin (0.001%-0.01%) treated sperm pellet. These studies suggest that the acrosomal membrane integrity remains unaltered by digitonin (0.001%-0.01%) treatment. All our biochemical studies confirm that we developed a permeabilization protocol, using 0.005% digitonin.
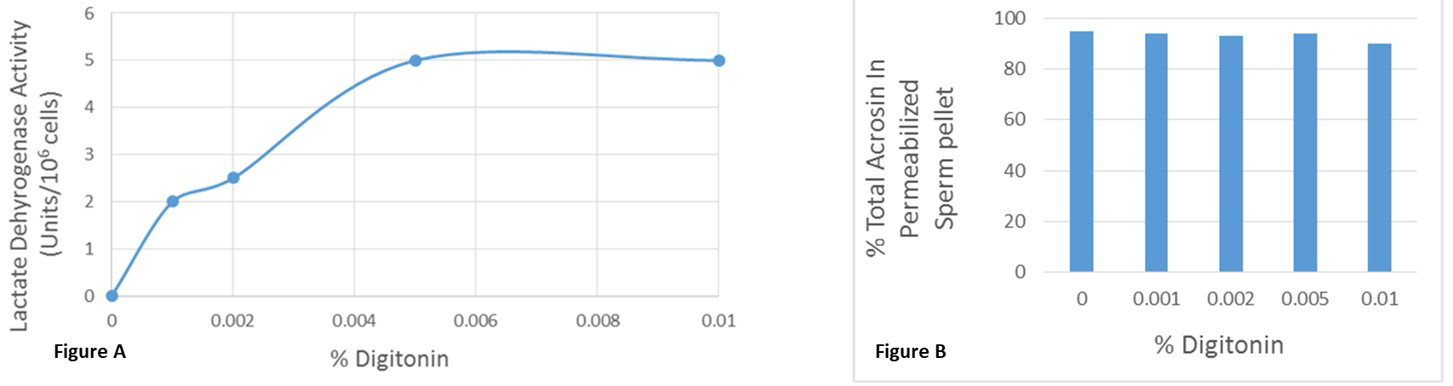
Figure 3: A: Permeabilization of bovine ejaculated sperm by digitonin was monitored by assaying lactate dehydrogenase (LDH) activity. Noncapacitated spermatozoa (5 X 107 cells) were permeabilized using various digitonin concentrations (0.001%-0.01%). Sperm were pelleted after centrifugation at 14,000g for 10 mins and the LDH activity was measured in the supernatant. The LDH activity in the supernatant was increased in a dose-dependent manner and it reaches at plateau in 0.005% digitonin. The data are representative of three separate experiments. B: To examine the acrosomal membrane integrity, acrosin activity was assayed in the control (digitonin untreated), and the different concentrations digitonin (0.001%-0.01%) treated sperm pellet. Acrosin activity of permeabilized sperm pellet remained unaltered by digitonin (0.001%-0.01%) treatment. The data are representative of three separate experiments.
Then to demonstrate a functional requirement for OMC32, Fab fragments prepared from the IgG fractions of anti-OMC32 were utilized and the inhibition of acrosomal exocytosis was examined using the release of acrosin as a potential marker. Noncapacitated and capacitated spermatozoa (5X107 cells) were permeabilized using 0.005% digitonin. Most of the acrosin activity remained in the pellet fraction (~86%) of noncapacitated permeabilized sperm; whereas the LPC induced acrosome reacted capacitated permeabilized spermatozoa resulted in the release of ~76% of the total acrosin into the supernatant fraction (Table 1).
In the presence of 10µg Fab fragments isolated from IgG against OMC32 polypeptide, 25% of the total acrosin was released into the supernatant (75% retained in the pellet) showing a significant reduction of acrosin release during the LPC-induced acrosome reaction (Table 1). As a control, approximately, 80% acrosin was released in the supernatant in the presence of 10µg Fab fragments isolated preimmune IgG. These experiments reveal the specificity of inhibition of acrosin release by 10µg Fab fragments isolated from IgG against OMC32 polypeptide during LPC-induced acrosome reaction of digitonin-permeabilized bovine sperm.
| % Total Acrosin | ||
|---|---|---|
| Supernatant | Pellet | |
| Noncapacitated Permeabilized Sperm | 14±3 | 86±5 |
| Acrosome Reacted Permeabilized Sperm | 76±4 | 24±6 |
| Acrosome Reacted Permeabilized Sperm in the Presence of 10µg Fab Fragments Isolated From IgG against the OMC32 Polypeptide | 25±5 | 75±6 |
| Acrosome Reacted Permeabilized Sperm in the Presence of 10µg Fab Fragments Isolated from pre-immune IgG | 80±4 | 20±6 |
Table 1: Inhibition of Acrosin Release from Digitonin-Permeabilized Sperm during the Acrosome Reaction in the Presence of 10µg Fa
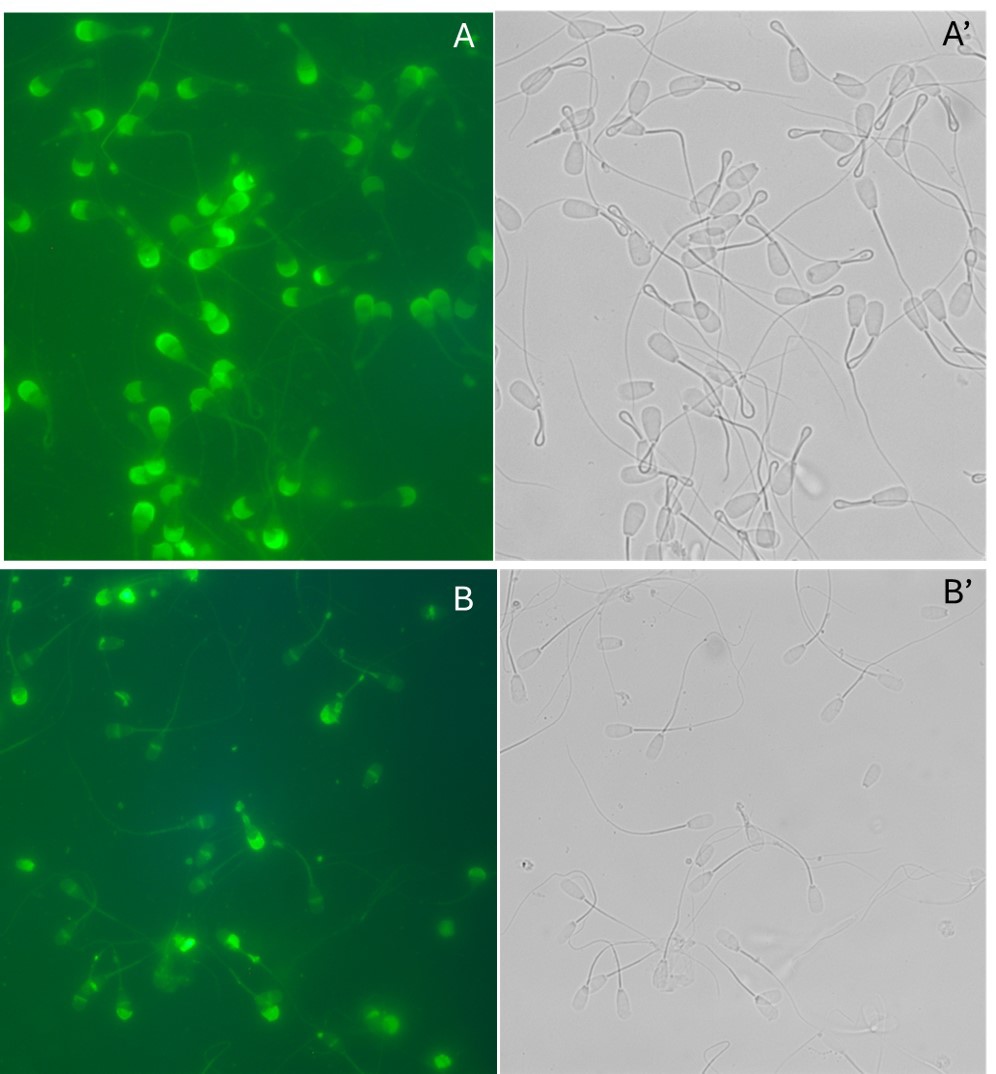
Capacitation efficacy was examined by detection of phosphorylation in capacitated sperm fractions both by immunocytochemically and biochemically (Figure 4). Immunofluorescence localization of tyrosine- phosphorylated polypeptides is presented in (Figure 4-I). The midpiece of flagellum of bovine cauda capacitated sperm population displayed an intense immunostaining with ant-PG10, whereas the head was almost unstained (Figure 4-IA). In contrast, noncapacitated spermatozoa displayed no fluorescence stained with anti-PG10 (Figure 4-IB). An array of polypeptides (Figure 4-II) underwent phosphorylation in the presence of heparin (capacitated pellet-lane 2 and acrosome reacted pellet-lane3). Since the
40kDa and 37.5kDa phosphorylated polypeptides existed in control pellet (lane 1), capacitated, and acrosome reacted sperm pellet fractions suggesting that both polypeptides are basal phosphorylated proteins. No bands were seen in the supernatant lanes of control (lane 4), capacitated (lane 5), and acrosome reacted (lane 6). All these biochemical studies reveal that the capacitation reaction was properly fulfilled. Acrosome reaction efficiency was also examined microscopically (Figure 5) using FITC-conjugated PSA (Pisum Sativum agglutin). A uniform signal over the entire acrosomal region (A) was observed for the acrosome-intact sperm and the acrosome-reacted sperm exhibited a signal over the equatorial region (B).
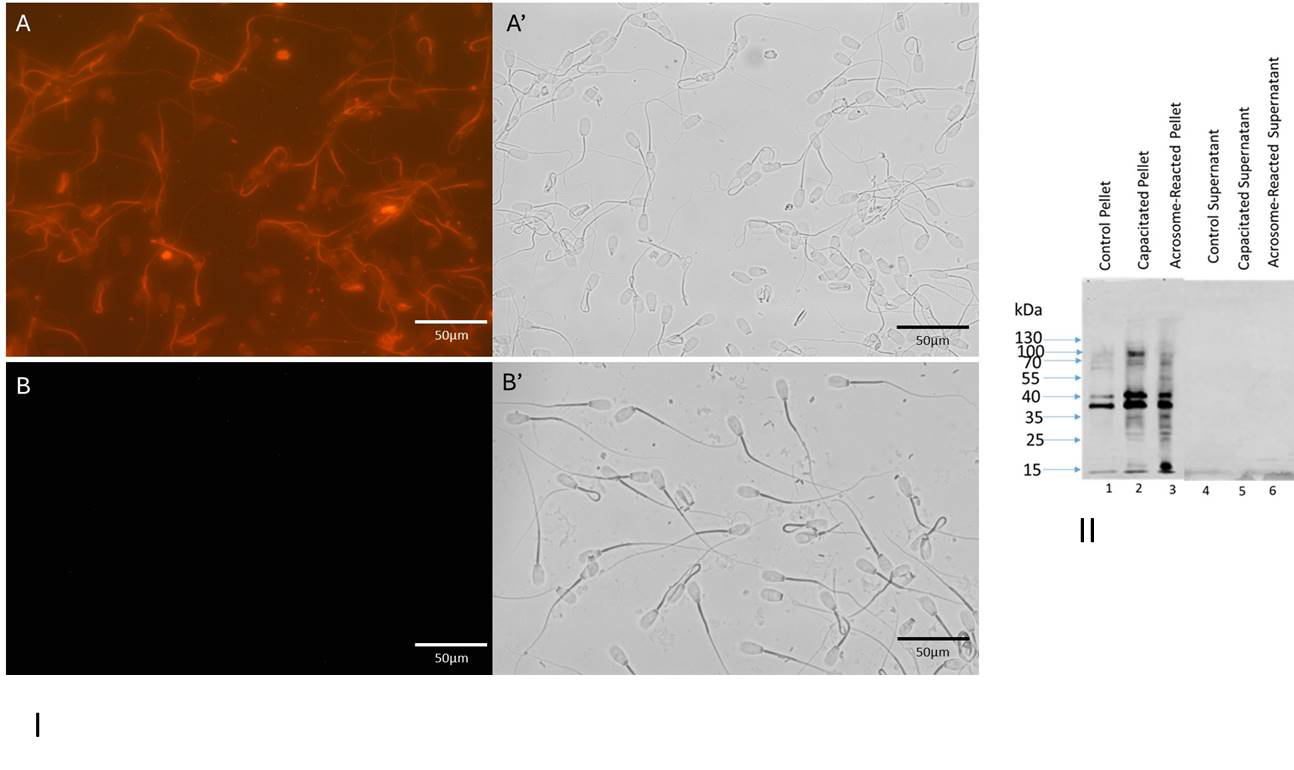
Figure 4: I: Immunocytochemical localization of tyrosine-phosphorylated proteins in capacitated (A and A’) and noncapacitated (B and B’) spermatozoa. Matched phase contrast (A’ and B’) and fluorescence (A and B) photomicrographs of spermatozoa stained with anti-PG10. Bar= 50µm. II: Protein tyrosine-phosphorylation pattern of bovine capacitated spermatozoa: Western blots of control pellet (lane 1), capacitated pellet (lane 2), and acrosome reacted pellet (lane3) stained with anti-PG10. No bands were seen in the supernatant lanes of control (lane 4), capacitated (lane 5), and acrosome reacted (lane 6).
Our results, utilizing permeabilized-cell models for exocytosis, strongly provide evidence that OMC32 plays a significant role in the regulation of acrosin release during acrosomal exocytosis.
Discussion
During mammalian fertilization, an array of acrosomal hydrolases is released from sperm acrosome required for sperm passage via the vestments surrounding the egg. In mouse, the capacitated spermatozoon penetrates through the cumulus oophorus and the zona pellucida of the egg to reach the oocyte plasma membrane [1]. In the current study, we have demonstrated that OMC32, IZUMO1, and SPACA3 exist in one large complex within the acrosomal matrix. Two proteins are critical for the gamete adhesion/fusion step during molecular fertilization (sperm IZUMO1 and oocyte JUNO) and their interaction is a ligand-receptor pair [24, 25]. Ellerman, et al. [24] reported that other mouse sperm proteins interact with IZUMO1 suggesting formation of a multiprotein membrane complex. It has been shown that IZUMO1 interacts with ACE3 (angiotensin1-converting enzyme 3) [25]. A human lysozyme-like protein, SLLp1, was known as an intra-acrosomal sperm protein, and SPACA3 stands encoded gene [26]. We have demonstrated the presence of a 25kDa SPACA3 immunoreactive band in the detergent- soluble fraction of bovine cauda epididymal sperm, and a sprinkle-type intense staining of SPACA3 observed over the apical segment staining with anti-SPACA3 [15]. Mandal, et al. [27] observed that the number of sperm fused per egg was decreased in the presence of anti-SLLP1 serum and indicated the role of SLLP1 in sperm/egg adhesion. It has been shown by immunofluorescent staining that the higher expression levels of SPACA3 and HSPA2 (heat shock 70kDa protein 2) in ZP-bound human spermatozoa and proposed that these two proteins are involved in sperm-ZP binding ability [28]. Our Co-immunoprecipitation data reveals that OMC32 interacts with SPACA3, and IZUMO1 [15]. Previously, it was shown an IZUMO-1 is present in a multiprotein complex [24, 25]. In this study, we demonstrate the presence of one large acrosomal complex consisting of three polypeptides (OMC32-IZUMO1- SPACA3). It could be possible that this complex may be involved in the recruitment of another essential molecular actor(s) involved in mammalian fertilization. Future study will address this issue.
Acrosomal matrix proteins exhibit different fates during the acrosome reaction. Several studies exhibit that during the acrosome reaction acrosomal proteins are proteolytically processed and released [3, 4, 10, 29], whereas electron microscopic studies revealed that other acrosomal matrix elements remain attached with the hybrid complex even after the acrosome reaction [30]. A 50kDa guinea pig acrosomal matrix protein is insoluble, disulfide-cross-linked, high molecular weight complex in intact spermatozoa which is converted to a 42kDa polypeptide and the 42kDa polypeptide is released from the acrosomal matrix during the acrosome reaction [29]. Acrosin, a trypsin-like serine protease, functions in both sperm binding and the penetration of the zona pellucida [1, 2]. It has been shown that in the presence of soybean trypsin inhibitor (acrosin inhibitor) human spermatozoa failed to penetrate zona pellucida [31] whereas in rabbits’ in vitro fertilization significantly reduced in the presence of anti-acrosin antibodies [32]. On the contrary, acrosin-deficient mice able to penetrate the zona pellucida and there is a delay of cumulus oophorus dispersion occurs than wild type [33]. Another study by Moreno, et al. [34] demonstrated a polyclonal antibody developed against a unique 18-amino-acid domain located in the polysulfate- binding domain of acrosin inhibits the in vitro fertilization of mouse zona-intact oocytes. Acrosin-knockout rat sperm can penetrate the zona and fertilize oocytes [35]. Hirose, et al. [36] generated acrosin-null hamsters using an in vivo transfection CRISPR/Cas9 system and showed that homozygous mutant males were completely sterile and acrosin-null sperm can travel to ovulated oocytes and are incompetent to fertilize the oocytes. Interestingly, it has been shown in human that a novel homozygous nonsense mutation of ACR (acrosin) lead to acrosin deficiency and the human spermatozoa lacking acrosin cannot penetrate the zona pellucida resulting in a total fertilization failure [37]. Various studies reveal that in acrosin plays a vital role in hamster and human fertilization. It is possible that several acrosome matrix polypeptides may serve as substrates of acrosin and release from the acrosomal matrix. The release of several acrosomal proteins (e.g., sp56, MC101) in the acrosin-deficient mice is delayed during the acrosome reaction compared to wild type mice [38], suggesting that acrosin is a potential marker for the proteolytically cleavage and release of acrosomal proteins during acrosomal exocytosis. Our results demonstrate the release of a substantial portion of SPACA3 (Figure 2A, lane 6) whereas OMCrpf remain particulate (Figure 2B, lane 3). Previously, we have demonstrated that OMCrpf remain amalgamated with the hybrid membrane complex after the LPC-induced acrosome reaction [10]. No release of SPACA3 occurred in the presence of acrosin inhibitors implying that acrosin contributes in the release of SPACA3 during acrosome exocytosis. Our future studies will investigate whether other acrosomal matrix proteins are proteolytically processed and released during the acrosome reaction.
We have demonstrated the function of OMC32 polypeptide during the acrosome reaction using a permeabilized sperm model. Digitonin-permeabilized guinea pig spermatozoa retained intact acrosomal membrane [22] and digitonin-permeabilized rat spermatozoa exhibit normal IP3-dependent acrosomal calcium efflux [39]. The permeabilization regimen creates pores in the plasma membrane, which permits the introduction of large molecules such as antibodies and permits slow release of cytosolic proteins; however, secretory vesicles remain functionally intact. Previously, we have shown that acrosin is released during LPC-induced acrosome reaction of bovine sperm [15], and the acrosome-reacted sperm displayed a signal over the equatorial region using FITC-conjugated PSA [40]. In the present study, we have shown a significant reduction of acrosin release during the LPC-induced acrosome reaction in the presence of 10µg Fab fragments isolated from IgG against the OMC32 polypeptide. Whereas as a control a significant release of acrosin of LPC-induced acrosome reacted bovine spermatozoa was observed in the presence of 10µg Fab fragments isolated from pre-immune IgG. Our study strongly demonstrates a functional requirement for OMC32 polypeptide, an acrosomal matrix polypeptide, in regulating the release of acrosomal hydrolases in the calcium- dependent membrane fusion events of acrosomal exocytosis. Future studies will address to elucidate the functional role of other acrosomal proteins during acrosomal exocytosis using our established permeabilized-cell models.
Funding: This work was supported by NSF grant 2153783 (Subir K Nagdas), and NC LSAMP NSF grant (2010124-FSU Subrecipient PI--Subir K Nagdas) Conflict of Interest: The authors declare that they have no conflict of interest. Authors Contribution Statement: Subir K Nagdas (Principal Investigator) planned the study, examined data, and wrote the paper. All experiments were performed by Tyrone Salters, Beonka Sharpe, and Deron Donald Jr.
References
-
Yanagimachi R (1993) Mammalian Fertilization In: Knobil E, Neill JD (Eds.), The Physiology of Reproduction. Raven Press, New York, USA, pp: 189-317.
-
Kopf GS, Gerton GL (1991) The mammalian sperm acrosome and the acrosome reaction. In: Wassarman PM (Ed.), Elements of Mammalian Fertilization. CRC Press, Boca Raton, Ann Arbor, Boston, USA, pp: 153-203.
-
DiCarlantonio G, Talbot P (1988) Evidence for sequential deployment of secretory enzymes during the normal acrosome reaction of guinea pig sperm in vitro. Gamete Res 21: 425-438.
-
Hardy DM, Oda MN, Friend DS, Huang TT (1991) A mechanism for differential release of acrosomal enzymes during the acrosome reaction. Biochem J 275: 759-766.
-
NagDas SK, Winfrey VP, Olson GE (1996) Identification of hydrolase binding activities of the acrosomal matrix of hamster spermatozoa. Biol Reprod 55: 1405-1414.
-
NagDas SK, Winfrey VP, Olson GE (1996) Proacrosin- acrosomal matrix binding interactions in ejaculated bovine spermatozoa. Biol Reprod 54: 111-121.
-
Baba T, Niida Y, Michikawa Y, Kashiwabara S, Kodaira K, et al. (1994) An acrosomal protein, sp32, in mammalian sperm is a binding protein specific for two proacrosins and an acrosin intermediate. J Biol Chem 269: 10133- 10140.
-
Nuzzo NA, Anderson RA, Zaneveld LJD (1990) Proacrosin activation and acrosin release during the guinea pig acrosome reaction. Mol Reprod Dev 25: 52-60.
-
Olson GE, Winfrey VP, Garbers DL, Noland TD (1985) Isolation and characterization of a macromolecular complex associated with the outer acrosomal membrane of bovine spermatozoa. Biol Reprod 33: 761-779.
-
Olson GE, Winfrey VP, Neff JC, Lukas TJ, NagDas SK (1997) An antigenically related polypeptide family is a major structural constituent of a stable acrosomal matrix assembly in bovine spermatozoa. Biol Reprod 57: 325-334.
-
Freemerman AJ, Wright RM, Flickinger CJ, Herr JC (1993) Cloning and sequencing of baboon and cynomolgus monkey intra-acrosomal protein SP-10: Homology with human SP-10 and a mouse sperm antigen (MSA-63). Mol Reprod Dev 34: 140-148.
-
Herr JC, Flickinger CJ, Homyk M, Klotz K, John E (1990) Biochemical and morphological characterization of the intra-acrosomal antigen sp-10 from human sperm. Biol Reprod 42: 181-193.
-
Liu MS, Aebersold R, Fann CH, Lee CY (1992) Molecular and developmental studies of a sperm acrosome antigen recognized by HS-63 monoclonal antibody. Biol Reprod 46: 937-948.
-
Beaton S, ten Have J, Cleary A, Bradley MP (1985) Cloning and partial characterization of the cDNA encoding the fox sperm protein FSA-Acr.1 with similarities to the SP- 10 antigen. Mol Reprod Dev 40: 242-252.
-
Nagdas SK, Smith L, Medina Ortiz I, Hernandez Encarnacion L, Raychoudhury S (2016) Identification of bovine sperm acrosomal proteins that interact with a 32-kDa acrosomal matrix protein. Mol Cell Bioche 414: 153-169.
-
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685.
-
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc Natl Acad Sci 76: 4350-4354.
-
Fairbanks G, Steck TL, Wallach DFH (1971) Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 10: 2606- 2617.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254.
-
Parrish JJ, Susko Parrish J, Winer MA, First NL (1988) Capacitation of bovine sperm by heparin. Biol Reprod 38: 1171-1180.
-
Mendoza C, Carreras A, Moos J, Tesarik J (1992) Distinction between the true acrosome reaction by a one- step staining method using Pisum sativum agglutinin. J Reprod Fertil 95: 755-763.
-
Noland TD, Olson GE (1989) Calcium-induced modification of the acrosomal matrix in digitonin- permeabilized guinea pig spermatozoa. Biol Reprod 40: 1057-1066.
-
Worthington V (1993) Lactate Dehydrogenase. In: Worthington Manual Enzyme Related Biochemicals, Worthington V (Ed.), Worthington Biochemical Corporation, USA, pp: 230-235.
-
Ellerman DA, Pei J, Gupta S, Snell WJ, Myles D, et al. (2009) Izumo is part of a multiprotein family whose members form large complexes on mammalian sperm. Mol Reprod Dev 76: 1188-1199.
-
Inoue N, Ikawa M, Okabe M (2011) The mechanism of sperm-egg interaction and the involvement of IZUMO1 in fusion. Asian J Androl 13: 81-87.
-
Zhang K, Gao R, Zhang H, Cai X, Shen C, et al. (2005) Molecular cloning and characterization of three novel lysozyme-like genes, predominantly expressed in the male reproductive system of humans, belonging to the c-type lysozyme/alpha-lactalbumin family. Biol Reprod 73: 1064-1071.
-
Mandal A, Klotz KL, Shetty J, Jayes FL, Wolkowicz MJ, et al. (2003) SLLP1, a unique, intra-acrosomal, non-bacteriolytic, c lysozyme-like protein of human spermatozoa. Biol Reprod 68: 1525-1537.
-
Leung ETY, Lee BKM, Lee CL, Tian X, Lam KKW, et al. (2023) The role of spermatozoa-zona pellucida interaction in selecting fertilization-competent spermatozoa in humans. Front Endocrinol (Lausanne) 14: 1135973.
-
Westbrook Case VA, Winfrey VP, Olson GE (1994) A domain-specific 50-kilodalton structural protein of the acrosomal matrix is processed and released during the acrosome reaction in guinea pig. Biol Reprod 51: 1-13.
-
Yanagimachi R, Phillips DM (1984) The status of acrosomal caps immediately before fertilization in vivo. Gamete Res 9: 1-19.
-
Liu DY, Baker HW (1993) Inhibition of acrosin activity with a trypsin inhibitor blocks human sperm penetration of the zona pellucida. Biol Reprod 48: 340-348.
-
Dudkiewicz AB (1983) Inhibition of fertilization in the rabbit by anti-acrosin antibodies. Gamete Res 8: 183-
-
Baba T, Azuma S, Kashiwabara S, Toyoda Y (1994) Sperm from mice carrying a targeted mutation of the acrosin gene can penetrate the oocyte zona pellucida and effect fertilization. J Biol Chem 269: 31845-31849.
-
Moreno RD, Bustamante E, Schatten G, BarrosInhibition C (2002) Inhibition of mouse in vitro fertilization by an antibody against a unique 18-amino acid domain in the polysulfate-binding domain of proacrosin/acrosin Fertil Steril 77: 812-817.
-
Isotani A, Matsumura T, Ogawa M, Tanaka T, Yamagata K, et al. (2017) A delayed sperm penetration of cumulus layers by disruption of acrosin gene in rats. Biol Reprod 97: 61-68.
-
Hirose M , Honda A , Fulka H, Tamura Nakano M, et al. (2020) Acrosin is essential for sperm penetration through the zona pellucida in hamsters. Proc Natl Acad Sci USA 117: 2513-2518.
-
Hua R , Xue R , Liu Y, Li Y , Sha X, et al. (2023) Acrosin deficiency causes total fertilization failure in humans by preventing the sperm from penetrating the zona pellucida. Hum Reprod 38: 1213-1223.
-
Yamagata K, Murayama K, Okabe M, Toshimori K, Nakanishi T, et al. (1998) Acrosin accelerates the dispersal of sperm acrosomal proteins during acrosome reaction. J Biol Chem 273: 10470-10474.
-
Walensky LD, Snyder SH (1995) Inositol 1,4,5-triphosphate receptors selectively localized to the acrosomes of mammalian sperm. J Cell Biol 130: 857- 869.
-
Nagdas SK, Baccas M, Dejean C, Richardson L (2016) Expression of Syntaxin-2 in bovine sperm. J Cell Biol Mol Sci 1: 1-7.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial