Pathotyping and Genetic Characterization of a Highly Virulent Newcastle Disease Virus Strain Isolated from Recent Outbreak in District Naushahro Froze, Sindh, Pakistan
In Pakistan, growth of poultry industry is confronted by many viral diseases and Newcastle Disease (ND) is on top. The present study was conducted for pathotyping and genetic characterization of NDV isolated from an outbreak in District Naushahro Froze region of Sindh, Pakistan. Post mortem examination of fresh carcasses (n=10) was performed and various organs like trachea, spleen, lung tissues, and cloacal contents were collected from birds showing typical lesions of the disease aseptically. Tissue homogenate was inoculated in 9 days old embryonated chicken eggs and allantoic fluid was harvested. The presence of virulent virus was checked by hemagglutination, hemagglutination inhibition using hyperimmune serum and ICPI assays. NDV was purified using sucrose gradient ultracentrifugation and confirmation was done by F-gene amplification, SDS-PAGE and mass spectrometry of fusion protein. The HA titer was 1:256, and hemagglutination inhibition was achieved using Anti NDV serum. A band of ≈ 800 bp of F gene (Acc.no Naushahro Froze /Pakistan/MH607122) was amplified which revealed 99% similarity with Sindh Agriculture university/Pak/2015/MF437287, Sindh Agriculture university /Pak/2016/KX791187 and Tehran/Iran/MG871466 strains. SDS-PAGE yielded five bands including Fusion protein with the estimated mass of 58896.0 Da. The presence of fusion protein and ICPI value was 2 which confirmed virulence of isolated strain resulting in high mortality. Thus, it is the need of the hour to characterize the prevalent indigenous strains of NDV to develop some novel vaccine strategies for effective control.
Introduction
At present intensive poultry production is suffering from heavy economic losses due to many reasons including infectious diseases. Newcastle disease (ND) is a Pathotyping and Genetic Characterization of a Highly Virulent Newcastle Disease Virus Strain Isolated from Recent Outbreak in District Naushahro Froze, Sindh, Pakistan major viral infection of the poultry and other birds, responsible for high mortality [1]. Outbreaks of Newcastle disease are continuously reported from across all the continents including Europe, Asia, Africa, South and Central America. NDV is endemic in Pakistan and many J Ethol & Animal Sci
other countries, particularly in backyard poultry which is most susceptible, moreover major economic losses are to commercial poultry industry. Host species include virtually all the wild as well as domestic birds including game birds, commercial and backyard poultry [2]. Rural poultry plays significant role in transmission of circulating NDV whereas transboundary transmission is mainly attributed to activity of the migratory birds.
Newcastle disease virus (NDV) belongs to Avian Paramyxovirus type 1 (APMV-1), genus Avulavirus, subfamily Paramyxovirinae, and family Paramyxoviridae. NDV genome is negative sense non-segmented single stranded RNA (-ve ssRNA) of about 15 kb in length [3]. NDV genome encodes 6 major structural proteins (5´ to 3´); comprising of a large RNA-directed RNA-polymerase (L), hemagglutinin-neuraminidase (HN), fusion protein (F), matrix protein (M), phosphoprotein (P), and nucleoprotein (NP) (Shabbir et al., 2018). The NDV virion is enveloped and it has two peplomers on its surface i.e. haemagglutinin-Neuraminidase (HN) and fusion (F) proteins. The HN protein is multifunctional and plays role in membrane fusion, cellular tropism and pathogenicity. Whereas, F protein is the major virulence factor and helps in the fusion process between cell membrane and virus [4]. Both of these glycoproteins are the major antigens of virus and hence also elicit a strong immune response. Moreover, a third activity; namely, fusion promotion activity has also been reported for HN protein, this activity is mediated by the stalk domain of the HN protein.
NDV is categorized into four path types based on mean death time (MDT) an in vivo assay to assess pathogenicity in embryonated chicken eggs. These path types are namely valogenic with MDT<60hr, mesogenic (MDT 60 to 90hr), lentogenic (MDT>90hr) and an avirulent type that does not kill the embryo. NDV is classified into six lineages as 1-6 on the basis of phylogenetic analysis of the F gene nucleotide sequence and each lineage is further categorized into sub lineages. Moreover, the understanding of viral protein structure and composition helps in establishing the interaction of virus with the host cells. NDV pathogenicity is mainly due to the amino acids residues present at cleavage site of the F protein [5].
Monoclonal antibody (mAb) reactivity has proven to be very useful in the antigenic analysis and epidemiological study of NDV strains from different outbreaks [6], however, molecular-based techniques for identifying and characterizing the NDV strains and for tracing their origins and spread is more definitive. An intensive and vigorous vaccination program is in practice from the last two decades or so in commercial poultry. Despite these measures, disease outbreaks occur frequently in some vaccinated flocks and rural poultry. Continuous emergence of new antigenic divergent may be the possible reason for vaccination failure. With the particular interest in obtaining information on local NDV strains that has caused outbreaks in commercial broilers in the recent years, current study was carried out for the pathotyping and genetic characterization of an indigenous NDV strain isolated from District Naushahro Froze region of Sindh, Pakistan.
Materials and Methods
Sample collection
Fresh carcasses (n=10) were collected from a broiler flock located in district Naushahro Froze .The flock was vaccinated against ND as per schedule. Trachea, spleen, lung tissues, cloacal and tracheal swabs were collected aseptically and transported to Department of Microbiology, Sindh Agriculture University Tandojam. The swabs were placed in chilled glycerol containing phosphate buffered saline and tissues were properly packed in frozen gel ice packs for transportation.
Isolation and identification of virus
The nine days old embryonated chicken eggs were used for inoculation through the allantoic route and fluid was harvested after 48 hours [7]. The fluids were first subjected to slide agglutination test using 0.5% chicken RBCs suspension. Samples showing agglutination were further subjected to micro hemagglutination assay for titration of virus [8]. Hemagglutination inhibition assay was employed using known Hyperimmune serum for confirmation of NDV.
Purification of Virus
The virion was purified by sucrose gradient ultracentrifugation technique. Briefly, 50, 30 and 20% filter sterilized sucrose (w/v) was layered in centrifuge tube followed by layering of filtered harvested allantoic fluid. Tubes were ultra-centrifuged at 120K X g for 2 hours at 40C. Clear middle layer (band) containing purified virus was collected and again filtered through 0.2 μm filter and stored at -700C [7].
Extraction of RNA and cDNA Synthesis
Total genomic RNA was extracted using QIAamp viral RNA mini kit (Qiagen™, USA). Purified RNA was quantified spectrophotometrically and stored at -200C [9]. 5 μg of template (viral RNA) was subjected to reverse transcription polymerase chain reaction (RT-PCR), using commercially available Superscript™ III reverse transcriptase kit. Random 9-mers were used for the synthesis of first-strand cDNA.
Amplification and Sequencing of Fusion Gene
cDNA was used for the amplification of viral fusion gene. Reaction mixture consisting of 100 ng template cDNA, 12.5 μl of Taq 2X master mix, 0.2 μl of each forward (20 pmol) and reverse (20 pmol) oligomer. Sequence of oligomers was: forward oligomer TGGAGCCAAACCGCGCACCTGCGG and reverse oligomer GGAGGATGTTGGCAGCAT [10]. Amplicons were observed in ethidium bromide stained 1% agarose gel and size of was estimated using 1kb DNA ruler.
Sequence Analysis
PCR amplicons were purified using QIAquick PCR purification kit and dispatched to Macrogen™, Seoul, Korea for DNA sequencing. Obtained sequences were analyzed with ChromasPro™ 2.6.5 software (Technelysium Pty Ltd, Australia) and NCBI's BLAST program (http://blast.ncbi.nlm.nih.gov/). Finally, aligned sequences were submitted to GenBank via BankIt (http://www.ncbi.nlm.nih.gov/WebSub/?tool= barcode) for allotment of Accession Numbers. Evolutionary relatedness was assessed using online program Phylogeny.fr (http://phylogeny.lirmm.fr/ phylocgi/simple_phylogeny.cgi) and MEGA™ software.
Protein extraction and SDS-PAGE
Virus particles were re-suspended in 6X gel loading buffer and incubated at 95°C for 150 minutes, followed by centrifugation at 14,000 X g for 15 minutes and clear lysate was used for SDS-PAGE. Proteins were separated on 8% Coomassie Brilliant Blue stained polyacrylamide gel. Molecular mass of protein bands was estimated by pre-stained protein ladder.
In gel digestion and Mass Spectrometry
Briefly, protein samples were run on SDS-PAGE stained using Coomassie Brilliant Blue and the band of interest was excised. A buffer containing resuspended trypsin was added and reaction mixture was incubated at 37°C for 24hr. Resulting peptides were extracted from the gel and analyzed by Q TOF Ultima Global (Micromass, Manchester, United Kingdom) mass spectrometer [11]. For protein identification using GPS Explorer software (V3.6, Applied Biosystems), obtained spectrum was submitted to MASCOT searching engine (V2.1, Matrix Science, London, UK)
Results
Clinical Diagnosis
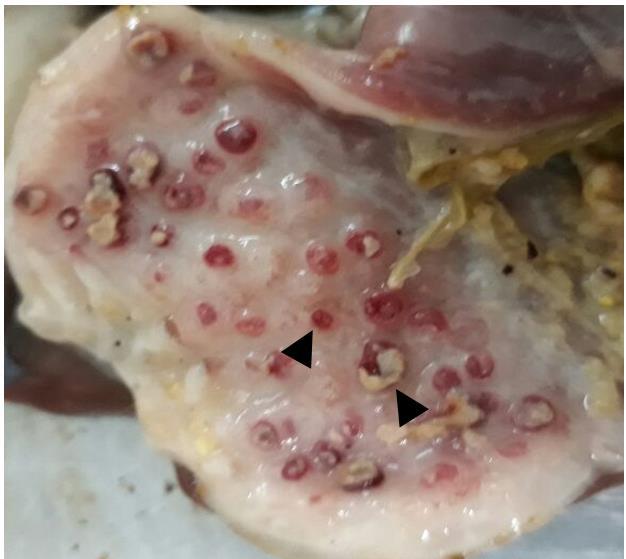
Based on the history, broiler flock was vaccinated as per vaccination schedule followed in the Naushahro Froze region; but the outbreak resulted in very high morbidity and mortality. There was loss in appetite, lethargy, weight loss, and greenish-white diarrhea in all chicken examined. The outbreak was clinically diagnosed based on respiratory distress, neurological signs (paralysis and torticollis) typical for the Newcastle disease. The gross postmortem lesions were tracheitis, pneumonia, myocarditis, encephalitis, catarrhal enteritis, nephritis, splenitis and hemorrhagic proventriculitis with some necrotic areas (Figure 1).
Serological Identification of Virus
Allantoic fluid was harvested from 9-day old embryonated chicken eggs. Initial identification was achieved through spot assay and all samples were found positive. Then samples were subjected to micro- hemagglutination assay and titer was found 1:32 to 1:256 HAU. Further, hemagglutination inhibition test using known Anti NDV serum confirmed the virus.
Purification of Virus
Filtered allantoic fluid was treated with sucrose gradient followed by ultracentrifugation, purified highly dense NDV band was obtained from the interphase of 30% and 50% sucrose gradients. The viral band was extracted out and after filtration stored at -80°C.
Genomic and Phylogenetic analysis
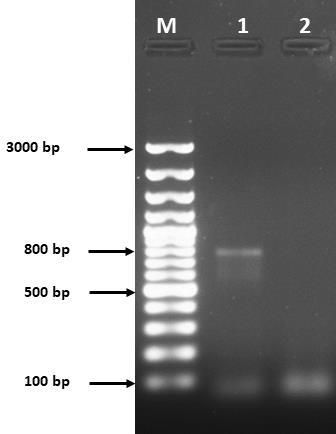
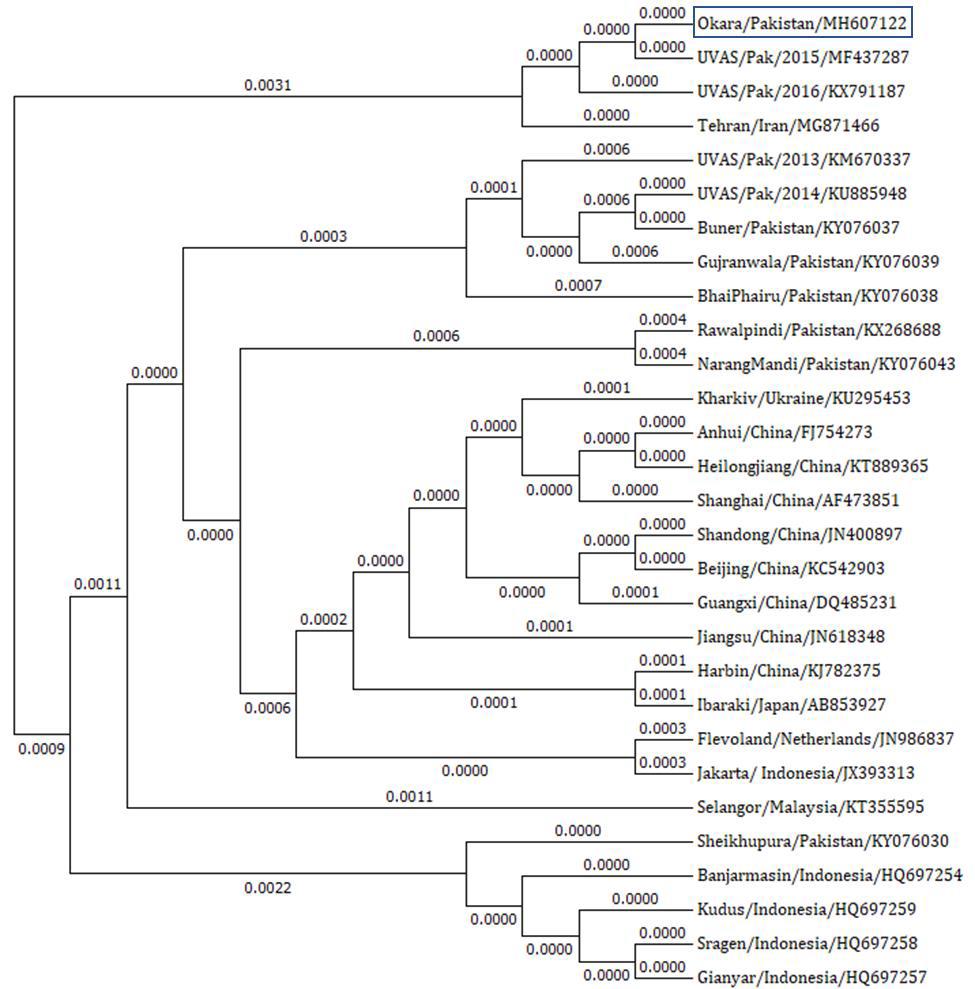
The extracted cDNA was subjected to PCR for amplification of viral fusion gene. The amplified product was visualized on ethidium bromide stained 1% agarose gel. The size of band was approximately 800 bp estimated by DNA ladder (Figure 2). The gel purified amplicon was sequenced from Macrogen® Korea. The sequence of fusion gene was analyzed by various bioinformatics tools and submitted to GenBank with accession number MH607122. The phylogenetic studies were executed and dendrogram was constructed by neighbor joining method (Figure 3). It revealed the highest identity (nearly 99%) of our sequence with UVAS-2015 strain (Accession No. MF437287) followed by UVAS-2016 strain (Accession No. KX791187) and Tehran strain (Accession No. MG871466). Our strain also revealed identity with different regional strains as follows: Iran, Ukraine, China, Japan, Netherland, Malaysia and Indonesia.
Proteomic studies
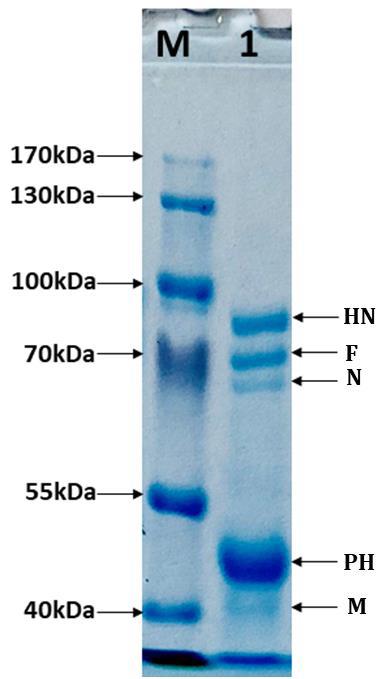
Proteins of purified virus were resolved by SDS-PAGE and visualized after Coomassie brilliant blue (Figure 4). Five different protein bands were observed in the gel and size of all bands was estimated by protein ladder. These five bands represent the viral haemagglutinin- neuraminidase (HN), fusion (F), nucleocapsid (N), phospho (PH), and matrix (M) proteins respectively.
Mass spectrometry
The band corresponding to fusion protein (approximately 55 to 60 kDa) was excised out and after in-gel digestion subjected to Q TOF Ultima Global mass spectrometer. The spectrum revealed two distinct spectral peaks which were subjected to MASCOT searching engine. From database search, the mass of protein was estimated 58896.0 Da and identified as viral fusion protein (Accession No. AAC28374).
Pathogenicity and virulence
Day-old chicks were injected with virus via intracerebral route and monitored regularly after 24 hours for period of eight days and ICPI (Intracerebral pathogenicity index) was calculated. The birds were scored from 0 to 2 (0 for normal, 1 for sick and 2 for dead cases). The score of our viral strain was 2, representing highest level of virulence. Moreover, amplification of F gene and presence fusion protein in SDS-PAGE analysis confirmed the molecular basis of pathogenicity and virulence.
Discussion
Newcastle disease (ND) is the most divesting disease of poultry industry, responsible for enormous economic losses in developing as well as developed countries. The virus can target different avian species ranging from domesticated poultry to wild bird. The infected bird develops various clinical manifestation and leads to mortality [2]. In current study, highly virulent NDV was isolated from a field outbreak of disease in vaccinated birds from district Naushahro Froze, Pakistan. Postmortem of infected birds displayed the lesions and necrosis in proventriculus, gizzard, spleen, intestine, and caecal area. Moreover, the birds depicted development of tracheitis, pneumonia, myocarditis, nephritis, splenitis and encephalitis [2]. The virus was cultivated in embryonated chicken eggs for propagation and allantoic fluid was harvested. Initial screening was achieved by slide agglutination assay with known antiserum. Titer 1:32 to 1:256 HAU was identified from micro hemagglutination tests and confirmed through hemagglutination inhibition assay, our findings are consistent with previous report [12]. The sucrose gradient ultracentrifugation was adopted for purification of virion from allantoic fluid [7].
Most appropriate method to characterize NDV is sequence analysis of fusion gene and protein profiling [13, 5, 14]. Our strain (Accession No. MH607122) displayed the highest identity (approximately 99%) with already known strains reported from Hyderabad, the SAU-2015 strain (Accession No. MF437287) and SAU- 2016 strain (Accession No. KX91187). After Hyderabad strains, it displayed highest levels of identity with Tehran strain (Accession No. KX91187). After Hyderabad strains, it displayed highest levels of identity with Tehran strain (Accession No. MG871466). According to phylogenetic analysis, our Naushahro Froze strain revealed identity with different Asian (Iran, China, Japan, Malaysia and Indonesia) and European (Ukraine and Netherland) strains, available on GenBank, NCBI (https://www.ncbi.nlm.nih.gov/genbank/) [15].
SDS-PAGE and Western blotting are rapid and appropriate methods to study the antigenic viral proteins. The viral proteins were characterized by SDS-PAGE
analysis and total five different viral proteins were identified. The 1-D gel clearly displayed five viral proteins namely haemagglutinin-neuraminidase, fusion, nucleocapsid, phospho, and matrix proteins. In SDS-PAGE gel, two distinct prominent signature protein bands corresponding to major glycoproteins the haemagglutinin-neuraminidase and fusion protein, whereas third band of the matrix proteins was observed [5, 16].
The mass spectrophotometric analysis of protein band comparable to fusion protein was performed. Two distinct spectral peaks were observed in the spectrum of fusion protein. According to the MS database search engines, the selected band was identified as viral fusion protein with 58896.0 Da and characterized as viral fusion protein (Accession No. AAC28374). These results are comparable to the data available on UniProt, TrEMBL and Swiss-Prot database (https://www.uniprot.org/uniprot/O90339). Existence of fusion protein indicates highly pathogenic nature of the virus. Virulence of virus was measured by intracerebral pathogenicity index. According to ICPI results, the isolated virus belonged to a highly virulent strain with maximum intracerebral pathogenicity index score of 2.
Generally, the outbreaks of ND are quite destructive with mortality rate up to 100%. In the current study, ND disease targeted the vaccinated flock and resulted in high degree of morbidity and mortality. There are many previous reports suggesting risk of substantial mortality even in vaccinated flocks [17]. There are multiple reasons responsible for infection in vaccinated flocks including dosage, gap between vaccination and challenge, and type of vaccine. Most important reason may be that vaccine can protect birds against clinical outcomes and mortality, but unable to block virus shedding among post-vaccinated healthy birds which is the main cause of disease spread [18, 19].
Based on the results, it was concluded that the indigenous NDV isolated from a field outbreak in Naushahro Froze region of Sindh Pakistan belonged to a very virulent strain. In Pakistan poultry industry, ND is still an endemic disease. Failure of vaccine may be due to inappropriate dosage and mismatch of vaccine. It is therefore the need of time to characterize our indigenous viral strains and to develop novel vaccine strategies like Virosome based vaccines for the better control of this destructive poultry disease from the Country in near future [20, 21].
Author’s contribution
MHR, MAZ and MS conceived the idea and designed the study. AAA helped in virus cultivation and identification. AM executed all the experiment work and analyzed the data. All authors interpreted the data, critically revised the manuscript for important intellectual contents and approved the final version.
References
-
Mariappan AK, Munusamy P, Kumar D, Latheef SK, Singh SD, et al. (2018) Pathological and molecular investigation of velogenic viscerotropic Newcastle disease outbreak in a vaccinated chicken flocks. Virusdisease 29(2): 180-191.
-
Susta L, Miller P, Afonso C, Brown CC (2011) Clinicopathological characterization in poultry of three strains of Newcastle disease virus isolated from recent outbreaks. Vet Pathol 48(2): 349-360.
-
Lefkowitz EJ, Dempsey DM, Hendrickson RC, Orton RJ, Siddell SG, et al. (2017) Virus taxonomy: the database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res 46(D1): D708-717.
-
Knipe DM, Howley P (2013) Fields virology, 6th (Edn.), Wolters Kluwer, Groningen, Netherlands.
-
Ren X, Xue C, Kong C, Zhang C, Bi Y (2012) Proteomic analysis of purified Newcastle disease virus particles. Proteome Sci 10(1): 32.
-
Russell P, Alexander D (1983) Antigenic variation of Newcastle disease virus strains detected by monoclonal antibodies. Arch Virol 75(4): 243-53.
-
Santry LA, McAusland TM, Susta L, Wood GA, Major PP, et al. (2018) Production and purification of high- titer newcastle disease virus for use in preclinical mouse models of cancer. Mol Ther Methods Clin Dev 9: 181-191.
-
Sharma L, Biswas U, Guha C, Chatterjee A, Jana PS, et al. (2018) Studies on thermo stability of newcastle disease viruses (Local isolates) for preparation of vaccine. Int J Curr Microbiol App Sci 7: 506-510.
-
Wajid A, Dimitrov KM, Wasim M, Rehmani SF, Basharat A, et al. (2017) Repeated isolation of virulent Newcastle disease viruses in poultry and captive non-poultry avian species in Pakistan from 2011 to 2016. Prev Vet Med 142: 1-6.
-
Mase M, Kanehira K (2012) Simple differentiation of avirulent and virulent strains of avian paramyxovirus serotype-1 (Newcastle disease virus) by PCR and restriction endonuclease analysis in Japan. J Vet Med Sci 74(12): 1661-1664.
-
Shevchenko A, Tomas H, Havli J, Olsen JV, Mann M, et al. (2006) In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat Protoc 1(6): 2856-2860.
-
Ullah S, Ashfaque M, Rahman SU, Akhtar M, Rehman A (2004) Newcastle disease virus in the intestinal contents of broilers and layers. Pak Vet J 24(1): 28-30.
-
Yu L, Wang Z, Jiang YL, Chang L, Kwang J (2001) Characterization of newly emerging Newcastle disease virus isolates from the People's Republic of China and Taiwan. J Clin Microbiol 39(10): 3512- 3519.
-
Shabbir MZ, Nissly RH, Ahad A, Rabbani M, Shubhada KC, et al. (2018) Complete Genome Sequences of Three Related Avian Avulavirus 1 Isolates from Poultry Farmers in Pakistan. Genome Announc 6(18): e00361-00318.
-
Wajid A, Dundon WG, Hussain T, Babar ME (2018) Pathotyping and genetic characterization of avian avulavirus-1 from domestic and wild waterfowl, geese and black swans in Pakistan, 2014 to 2017. Arch Virol 163(9): 2513-2518.
-
Xu X, Ding Z, Yuan Q, Ding J, Li J, et al. (2019) A genotype VII Newcastle disease virus-like particles confer full protection with reduced virus load and decreased virus shedding. Vaccine 37(3): 444-451.
-
Perozo F, Marcano R, Afonso CL (2012) Biological and phylogenetic characterization of a genotype VII Newcastle disease virus from Venezuela: efficacy of field vaccination. J Clin Microbiol 50(4): 1204-1208.
-
Miller PJ, King DJ, Afonso CL, Suarez DL (2007) Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 25(41): 7238-7246.
-
Van Boven M, Bouma A, Fabri TH, Katsma E, Hartog L, et al. (2008) Herd immunity to Newcastle disease virus in poultry by vaccination. Avian Pathol 37(1): 1- 5.
-
Manual (2012) Newcastle disease. Office International des Epizooties, Paris, France.
-
Resplandor G, Umali D (2018) Molecular and serological detection of Newcastle disease virus in native chickens from selected live bird markets in Batangas, Philippines. Adv Anim Vet Sci 6(1): 1-7.
- In Situ Evaluation of the Anthelmintic Effect of the Aqueous Extract of Syzygium aromaticum (L) Merr and Perry on Bovine Strongyles
- Successful in Vitro Embryo Production with Oocytes Aspirated from Live White-Tailed Deer (Odocoileus Virginianus Texanus) Donors under Captivity in Northeast Mexico
- Bangladeshi Finches with their Evolutionary Thoughts (Aves: Passeriformes)
- Note on the Survival Status of Przewalski’s Horse, Equus ferus przewalskii (Perissodactyla: Equidae)
- In-Situ and Ex-Situ Protection of White-Breasted Waterhen (Amaurornis Phoenicurus) (Pennant, 1769) (Aves: Rallidae)
- Meat Examination in the Laboratory, the Acceptability and the Human Health