Daily Activity and Foraging Patterns of Adult Golden Marmots in Pup-Rearing Burrows in Relation to Habitat Disturbance in Karakorum Range Pakistan
Animals may change the temporal dimension of their niche by shifting their activity patterns in relation to anthropogenic disturbances. Yet, few studies have documented these response phenomena in pasture land. We examined the extent to which disturbances related with habitat features altered the timing of foraging and activity patterns of golden marmot (Marmota caudata aurea). Using a scan-sampling observational method, we collected data from 34 pup-rearing marmot burrow sites in the summer pasture area of Shigar Valley, northwest Pakistan. We defined frequency of occurrences index (FOI) of anthropogenic disturbance intensity for each site based on the presence of livestock, herders, distance from livestock corrals or shepherd huts, and density of major tracks, and determined whether marmot time spent aboveground, as well as foraging distance from pup-burrows, was or not between high and low disturbance sites. We found that disturbance intensity influenced the total daily activity time and foraging distance from pup burrows. Our results indicated that marmots adjusted their diurnal patterns of activity and the distances moved from their pup burrows in relation to the timing of anthropogenic activities implying changes to the temporal and spatial dimension of their niche. In areas with high levels of disturbance, marmots were less active during morning periods and more active during midday and compensated by increasing activity when herding activity was less. Foraging activity was less during midday and showed a peak during late evening. Finally, results suggest that grazing intensity of human disturbances may have significant consequences in terms of marmot summer pup-raising periods for energy intake.
Introduction
Predation risk plays a prominent role in shaping herbivores foraging activity patterns [1, 2]. For example, herbivores may alter their activity patterns to evade or decrease the risk of predation [3, 4] or extent of inter-specific competition for resources [5]. Temporal segregation is one tactic species can differentiate their ecological niche [6], evade predation risk, and coexist with other animals [3]. For many species, temporal activity adjustment is also a beneficial approach for dodging high levels of human disturbance [7]. Theoretical studies propose that wild animals often perceive humans as a latent predator, and human stimuli are perceived to have greater consequences than the threats posed by predators in the landscape [8, 9]. However, temporal changes in animal activity patterns can suffer a cost because many species rarely adjust to maintain foraging efficiency at different times of the day [6]. Animals that are forced to forage in suboptimal activity periods to evade the risks related with human activities frequently experience deficits in their energy budgets [10, 11]. For example, species with an inadequate proficiency to move away from a threat, such as those with small home ranges or knotted to a burrow are more in danger to risks related with human disturbances [12].
In Northern Pakistan, especially in the Shigar Valley Karakorum range, wildlife populations such as red foxes (Vulpes vulpes), grey wolf (Canis lupus) experience severe punitive killings due to the availability of sanctioned firearms, closing or smoking out dens [13], and decline caused by fuel wood collection and free range livestock grazing in pasture land [14, 15]. Besides this, livestock grazing has a defined negative influence on wild animal activity patterns due to competition for habitat resources [16, 17]. These alterations are often triggered by the physical occurrence of livestock and herders [18, 19]. Livestock and their herders are consequently measured as a form of disturbance [20] and human activity in the pastureland differs throughout the day with some times being more highly disturbed than others. Accordingly, wild animals often adapt by shifting their natural patterns of activity [7]; however, few studies have recognized this phenomenon especially within free grazing pasture land [21]. In such a conflict space, foraging is principally crucial for hibernating mammals such as marmots (Marmota spp.), which display a circannual rhythm of energy consumption and body mass [22], such that heightened energy consumption during active summer months is intensely related with winter survival, reproduction, litter and population size [23, 24], along with lactation period timing in females [25]. In addition, other behaviors such as vigilance are influenced by numerous factors, including distance to burrow, visibility, distance to other conspecifics, age and sex, reproductive status, type of stimuli, parental status, human activities and livestock existence [8, 19, 26, 27, 28, 29, 30, 31, 32].
Worldwide, there are 14 marmot species which are large burrowing and hibernating squirrels [26, 33]. They have adopted various survival tactics to cope with habitat stimuli, such as changing time rhythm [18, 19], or using habitats farther away from human activity [34, 35, 36], or constructed extra burrows for shelter (e.g., Vancouver Island marmot M. vancouverensis [37]as well as selected big stones to facilitate better scanning (e.g., Alpine marmot M. marmot [34, 38] and used predated rocky burrows to reduce threats (e.g., golden marmot M. caudata aurea [15]. Ecologically, marmots are important prey for Golden Eagle (Aquila chrysaetos), Tibetan wolf (Canis lupus chancho) and snow leopard (Panthera uncia) [39, 40]. Golden marmots are currently classified as least concern by the International Union for the Conservation of Nature (IUCN) however their population is declining and requires monitoring for conservation and estimation [41].
The aim of this study was to examine the diurnal activity patterns of golden marmots in relation to risks associated with humans or livestock disturbance. Marmots depend on burrows as a shelter to escape from potential predators and as a place to raise newborn pups [27, 41]. Consequently we hypothesized that both daily activity outside the burrow (hereafter referred to as “daily activity patterns or activity budgets”) and the distance travelled from the pup burrows (hereafter, denoted as “foraging activity patterns or foraging distance”) would be related to the anthropogenic threats [42, 43]. Our specific aims were to investigate and report: 1) To what magnitude do marmot activity levels changes between areas experiencing high and low intensities of human or non-human disturbance, and 2) In relation to disturbance intensity from grazing and human activity, do marmots alter their above-ground activity patterns and foraging distance from pup-raising burrows?
Materials and Methods
Study site
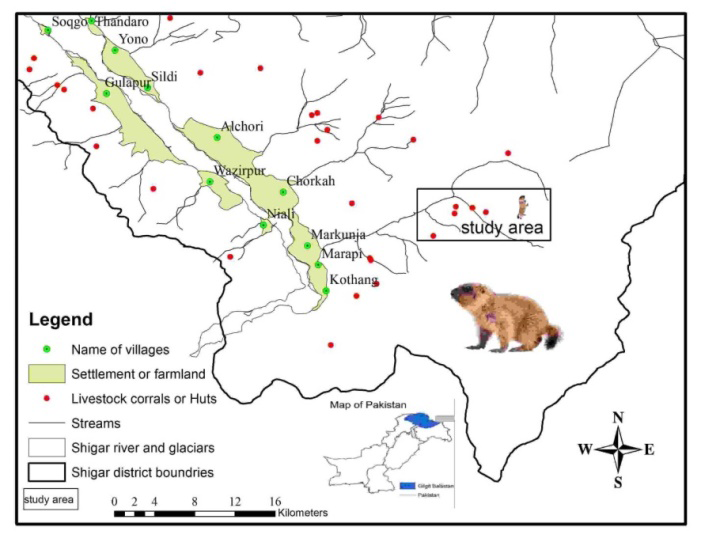
This study was conducted in the Shigar Valley of the Karakorum Range, located along the northern bank of the river Indus (Figure 1). The study sites were Blasting, range, Abasing mullah, Shendachan, Zembroq, Karfu khombu and these selected sites were provided suitable habitat for marmots. The study area lies at 25° 25′32″ N latitude and 75° 42′59″E longitude and covers an area of 4373 km2 [14, 15, 44]. The valleys are dry with an annual rainfall of aro_u_nd 200 mm and a maximum of 600 mm, and a maximum elevation of 3962 m. The mountains surrounding the whole shigar valley have a total of 84 medicinal plant species belonging to 72 genera and 36 families, which are used by the residents of the area to treat several health disorders [44]. The ungulate community consists of Himalayan ibex (Capera sibirica), musk deer (Moschus chrysogaster), and the very rare ladakh urial (Ovis orientalis vignei), the astor markhor (Capra falconeri) has been locally extirpated from the study area about 30 years ago. The small herbivores consist of Indian pika (Ochotona spp.), Cape hare (Lepus capensis) and Golden marmots (Marmota caudata). Small carnivores include weasels (Mustela spp.) and stone marten (Martes foina). Large carnivores include gray wolves (Canis lupus), snow leopards (Panthera uncia) and red fox, and raptors include Golden Eagle (Aquila chrysaetos) and Steppes eagle.
In shigar valley Balti ethnic group are still strongly tied with their culture in the construction of livestock corrals or shepherd huts. Further, in the summer season from April to September each years, the inhabitants take their livestock –including goat (Capra hircus), sheep (Ovis aries), cow (Bos indicus), yak (Bos grunniens) or local breed zou (male) or zouno (female), and horse (Equus caballus) and donkeys (Equus spp.) – into the pastureland for free grazing.
In Shigar Valley in the Karakoram Range, at the end of April snow melts on the south-facing slope and in May, golden marmots start to emerge from burrows at lower elevations; by mid-May, marmots at high elevations have also emerged. Marmots are active as a diurnal animal until the end of September, when they return to a state of hibernation in their burrows [15, 25, 27].
Experimental Site Selection
In this study, the observations were completed in sub- alpine pasture lands at high elevation (approximately 3216 – 4253 m) for the period of the active summer months (10 June – 20 August 2021), which represents the pup-raising time. Golden marmot average home range size for early season (April or May) is larger (3.1 ha) than in late season (June-September) (2.9 ha) [25]. Main burrows typically have between 1 and 6 (or more) openings and their distance between main burrows to neighboring escape burrows were 132 – 146 m. Marmot group sizes were between 1 and 10 individuals and were male-dominated colonies [25]. Pups generally emerged above ground the last days of June or first days of July; back-dating allows us to estimate their age as approximately 10 weeks old (mating generally occurred between the third week of April and second week of May, gestation is 30-33 days, plus ca. 30-40 days for weaning [15, 25].
In total, we considered 34 sites based on anthropogenic activity and they represent the variation in the landscape of threats along with easy to spots targeted marmots in which to conduct our temporal activity and foraging distance experiments. We defined a marmot “site” as an area covering the natal burrows (hereafter, we considered natal burrows which had pups to be called pup burrows) engaged by a family group or cooperating family group surrounded by a radius of around 100 m that was disconnected from other marmot sites by a minimum of 300 m. This measure was established in order to minimize pseudo-replication of recording the same individual in more than one site and to confirm that sites were spatially independent of each other. The number of adult marmots in a pup burrows site ranged between 2 and 5 individuals, and at each pup burrows, the number of pups ranged between 2 and 4.
For each marmot site, we determined frequency of occurrence index (FOI) of continuous disturbances from anthropogenic activities calculated based on the following characteristics (Figure S1): 1) Existence of livestock, or humans on foot, with donkeys, tree cutting, milking or guarding livestock (shepherds or herders), as well as local hikers (assumed livestock owners) [19, 45]; 2) Distance to each livestock corral or shepherd huts [14] and; 3) The density of major foot Marmota camtschatica bungei trails and grazing trails [23, 19] (for examples, see for details in photographic evidences (Figure S1)).
Quantitatively, at each site, the number of livestock and humans that stayed on or passed through the site was documented persistently during daylight hours (0700 – 1900hrs) over 6 successive days, and then, each factor’s values were averaged over the period to generate 1 set of values per site, such as one combined value for livestock and one collective value for people for each FOI in over 6 successive days, The total length of main human trails or grazing trails was visually assessed in each site as considering a distance from focal marmots burrows to nearest trails in meters. The distance from each site to the nearest shepherds huts or livestock corrals was measured using handheld GPS units and subsequently calculated in ArcGIS v. 10.2 (ESRI Inc.), as in our earlier work Zaman M, et al. [14]. Each feature was first scaled and assigned an ordinal value between 0 and 5 based on their higher or lower FOI of anthropogenic activity. We then calculated a combined score as FOI of disturbance for each site. Finally, as the FOI scores exhibited a major bimodal distribution, we then classified the sites based on these FOI scores as having either “low” or “high” disturbance. This method resulted in a total of 18 “low” and 16 “high” disturbance sites for performing our study of marmot observations. To elaborate, high disturbance sites were those sites that were usually proximate to shepherd huts or livestock corrals, had high levels of livestock grazing and humans involving in fuel wood collection, guarding livestock or camping, as well as using bonfires for heat and the site contained well-defined human and livestock trails. In contrast, low disturbance sites were further away from shepherd’s huts or livestock corrals and had a low existence of livestock and humans.

Figures S1: Photographs in the study area of (a) grazing intensity in pastureland, (b) tree logging or fuel wood collection, including a donkey used for heaping logged trees (junipers, birch, wild rose), (c) livestock corrals and shepherd huts, (d) a marmot pup scanning for threats (note: selection of big stones), (e) an adult female showing vigilance at a pup burrows, and (f) pup scanning, with only head out of burrow, and adult conspecific resting as well as watching for threats.
Scan Sampling Observations
The circadian activity patterns of adult focal marmots (≥ 2 years old) outside pup burrows were recorded via observations through 10 × 42 range finder binoculars; pups in this research were excluded from our datasets. Marmot activity shapes and high or low anthropogenic disturbance activities time were determined using a scan-sampling method [46, 47, 48], where the activities of all observable focal marmots were documented during 15-min intervals in daylight hours over 2 days at each site; only clear sky days were chosen for sampling. For all active focal marmots outside pup burrows at each scan, we noted the foraging distance and foraging activity patterns (e.g., slow walking in search of food resources), as well as other daily activities (resting, vigilance or whistling, nursing pups, basking, playing, fighting, digging or grooming activities) from pup burrows for was noted [49, 50]. We measured foraging distance travelled by adult marmots through following techniques, if the animal was directly between observer and the pup burrows; we calculated animal distance from burrow using visually inspection or rangefinder binoculars in meters. But, if the animal was to the left/right of the burrow, the distance measured by the rangefinder or visually inspection were the similar from observer to the animal and from animals to pup burrows as described by Poudel, et al. [46]. We do not collected group size or sex, age classes and controlled their effect in our study (no marking marmots) and Observations were completed from vantage points (≥ 70 m from the focal marmots), at which they exhibited no indication of responses to observers.
Data Analysis
A total of 48960 hours (34 sites × 2 days × 720 minutes/ day) of direct observations of 119 different focal adult marmots was collected during this study. The dataset contained 12240 scans (34 sites × 2 days/site × 180 scans/ day) and each scans we mostly found two or three Individuals above ground. For each site, the amount of time focal marmots spent above ground was recorded as daily timing activity (hours, minutes, and seconds) and percentages of time spent for diverse temporal activity were calculated by using a stopwatch on the mobile phone. The maximum number of focal marmots detected in any one scan on any day was used as an estimate of the total number of focal marmots in each study group (site). For each site, we averaged the percentage of the total individuals seen above ground at each scan to gain a single value per site. This averaged proportion was applied to determine the average time spent above ground per individual.
Daily activity patterns of focal marmot and foraging activity, as well as distance from pup burrows, were recorded during three time periods: morning (0700hrs –1000hrs), midday (1001hrs – 1600hrs), and evening (1601hrs –1900hrs) and others rest of the time hours were considered as inactive marmots. We also recorded activity time during three time periods for low or high disturbance stimuli in selected sites [42].
In our set of models, the daily activity pattern and the foraging activity of focal marmots above ground were compared with disturbance activities time for both low and high disturbance sites by using non-parametrically as kernel density functions with the package “Overlap” using the default bandwidth parameters [51, 52], following the assumption that animals’ activity through any time scale are similarly expected to be “trapped” during any period of their activity [53]. Circular density curves for focal marmots were compared with disturbance activities using the coefficient of overlap (‘overlap coefficient’∆), with values ranging from 0 (no overlap) to 1 (complete overlap), as proposed by Ridout and Linkie [54]. To calculate whether focal marmot daily activity patterns and foraging activity varied in relation to different disturbance activities time, we tested the daily activity patterns or foraging activity was related with the disturbances, we used a Spearman’s rank correlation coefficient test.
In our second set of models, we compared the percentage of energy budgets for daily activity and average foraging distance travelled by marmots between the two levels of disturbance (high or low), as well as among the three periods of day. Data were tested for outliers and normality, according to protocols designed by Zaman M, et al. [14]. Daily activity percentage data were square root transformed and average distance traveled was log-transformed to fulfill rules for normality. A Shapiro– Wilk’s test, Wald t test, Kolmogorov-Smirnov test and graphic examination of histograms were made to confirm that the transformed data were normally distributed. In our third set of models, Generalized Linear Mixed Effect Models (GLMMs) were used to assess the impact of high and low disturbance, proximity distance to livestock corrals, shepherd huts human trails and livestock grazing trails, and time of day, on marmots’ percentages of energy budgets for daily activity. The function lmer or lme of the library lme4 [55] in the R package was used for fitting the GLMMs; we set percentage of energy budgets for daily activity of marmots as a response variable and explanatory variables were set as a binary measure of disturbance activities time (high disturbance as 1, and low disturbance as 0), time of day used for collect to percentages of energy budgets for daily activity set as three levels (morning, midday, evening time) and we also included their interaction (disturbance × time of day). Distance to nearest livestock corrals or shepherd huts, and distance to human and livestock trails were included as fixed factors for analyses. Study site was used as a random factor to account for potential correlation among observations within site through periods and uneven sample sizes as well as removed from the model when not influenced on predictors [56, 57]. In our fourth set of models, we again used GLMMs to measure the influence of high and low disturbance, vicinity distance to livestock corrals or shepherd huts, human trails and livestock grazing trails, and time of day, on marmots’ average foraging distance as similar description were used above paragraph for third set of models. We designated the most parsimonious model based on AIC that best described daily activities and foraging distance of Marmots. Finally, we also used Mann–Whitney test to grouping comparison between the FOI of anthropogenic activists and distance to corrals or huts for high and low disturbance defined sites [42]. All the analyses were conducted in R 3.1.2 and the nonparametric Z-statistics were completed in SPSS 20.00 (IBM SPSS Inc., and Chicago, Illinois, USA).
- Results
- In this study’s analyses, we used 34 sites for direct observation of livestock and human disturbance and detected
- 142,469 disturbance grazing intensity by local breeds of zou and zouno was higher in both low and high disturbance sites, followed by cattle grazing and then sheep, yaks and others
- (Table 1).
- Disturbance Intensity (Number Of Individuals/Day/Site) (Mean ± SD)
- Disturbance Type
- Low Disturbance Sites
- High Disturbance Sites
- U
- P
- Sheep
- 13.55 ± 25.50
- 22.50±37.50
- 67.5
- 0
- Goats
- 16.71 ±27.50
- 18.50 ±31.50
- 127.5
- 0.585
- Cow
- 17.05 ±32.00
- 18.07 ±71.00
- 134
- 0.767
- Yaks
- 2.50 ±5.16
- 15.70 ±32.08
- 79.5
- 0
- Zou and zouno
- 247.50 ±214.00
- 257.50 ±317.50
- 113
- 0.294
- Donkeys
- ±
- 1.97 ±8.58
- 132
- 0.624
- Human activities
- 1.44 ±2.50
- 3.50 ±4.37
- 137.5
- 0
- Distance to corrals
- 929.06 ±155.21
- 195.59 ±272.23
- 127.5
- 0
- Distance to livestock trails
- 103.10 ±663.23
- 61.68 ±881.21
- 94
- 0
- Distance to human trails
- 141.57 ±340.15
- 62.65 ±112.33
- 124
- 0.52
Table 1: Frequency of occurrences index (FOI) of number of people and livestock animals in all study sites over a six-day period
A non-parametric Mann–Whitney test to grouping relationship between the FOI of anthropogenic activists and distance to corrals or huts and proximity distance to nearest livestock and human trails were explained both significantly differed or not differed for low and high disturbance activities in two different disturbance site (Table 1).
Effect of Habitat Disturbance on Diurnal Activity Patterns
Overall, adult marmots were above ground for an average of 65.83 % of day time, approximately equivalent to 4h (±38min) per day. In high disturbance sites, adult marmots spent approximately 349 h above ground per day, whereas in low disturbance sites, the time spent was 428 h per day. Disturbances had negative influence on marmot daily activity energy budgets (total time above ground), comparing high and low disturbance sites (t value = -3.781, p < 0.000).
Furthermore, time of day (t value = 2.392, p < 0.018) and distance to livestock grazing trails (t value = 1.945, p < 0.053) were significantly positive effect on daily activity energy budgets of marmots. Human used trails were significantly negative effect on daily activity energy budgets of marmots (t value= -4.563; p < 0.005) and distance to livestock corrals or huts showed no significant effects (Table 2). The proportion of time marmots spent above ground reached up to 50% during the peak midday and 38 % in afternoon periods (Figures 2a & 2b). Marmot daily activity patterns was greater during in the midday period in high disturbance activities sites and dipped activity during in the late evening, while this midday pattern was consistent in low disturbance activities sites but in the late evening increased activity was observed. Furthermore, daily activity patterns of adult marmots out of pup burrows were highly overlapped with higher disturbance activities in high disturbance sites with corresponding to low disturbance activities in low disturbance sites (Figure 2a & 2b).
| Coefficient | Std. Error | t value | P | |
|---|---|---|---|---|
| Intercept | -0.554 | 0.048 | -11.34 | 0 |
| Disturbance (high vs. low) | -0.271 | 0.072 | -3.781 | 0 |
| Time of day | 0.172 | 0.071 | 2.392 | 0.018 |
| Disturbance × time of day | 0.208 | 0.507 | 0.414 | 0.679 |
| Distance to livestock grazing trails | 0.221 | 0.114 | 1.945 | 0.053 |
| Distance to human trails | -0.258 | 0.057 | -4.563 | 0.005 |
| Distance to livestock corrals or huts | 0.248 | 0.51 | 0.484 | 0.629 |
Table 2: Percentage of daily activity energy budgets of adult marmots outside pup burrows in relation to different habitat stimul
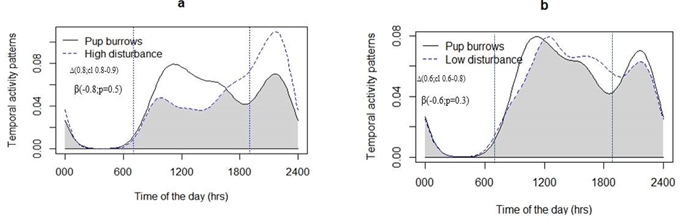
Gray shading represents the area of overlap using the coefficient of overlapping (‘overlap coefficient’∆) and cl (Confidence intervals), and Spearman rank correlation (SRC) between the adult marmots and two level of disturbance activities time and beta (coefficient), P represented significant level.
Effect of Habitat Disturbance on Foraging Activity Patterns from Pup Burrows
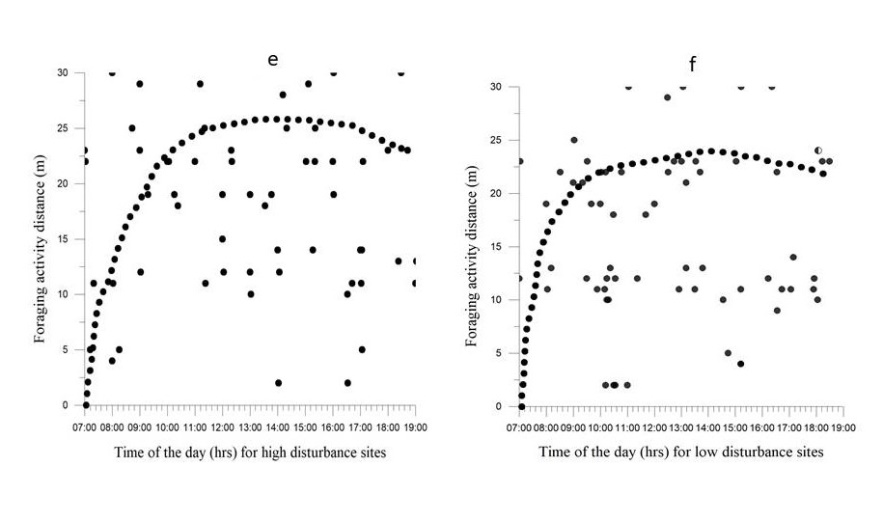
Overall, when above ground, marmots were spent most of their foraging activity time approximately 2h (±11min) per day, (19.71%) within 23 m from a burrow in both categorized sites. Marmot (30 %) of their foraging activity was midday dipped as compared to in the evening time for both high and low disturbance activities sites. The average foraging distance in high disturbance sites (13.8 ± 6.5 m) was approximately 19 % greater than that of low disturbance sites (9.7 ± 2.5 m). The maximum foraging distance traveled that we observed was 65 m in high disturbance sites and 58 m in low disturbance sites (Figure 2e & 2f). Disturbances had negative influences on marmot foraging distance, measured as total time above ground travelling in search of food, whereby searching time was higher in high disturbance sites than in low disturbance sites (t value = -2.671; p < 0.00). Furthermore, time of day had a positive significant effect on foraging distance (t value = 2.331; p < 0.016) and distance to livestock corrals or huts showed negative significant effects on foraging distance (t value = -2.216; p < 0.037) (Table 3). The foraging activity distance traveled by marmots in high disturbance activities sites was greater in the evening and in the afternoon and lower during the morning and at midday, unlike marmots in low disturbance activities sites which were more uniform across time periods, especially midday through to evening (Figures 2c & 2d). Furthermore, foraging activity of adult marmots highly overlapped with disturbance activities in high disturbance sites (Figure 2c & 2d).
| Coefficient | Std. Error | t value | P | |
|---|---|---|---|---|
| Intercept | -0.54 | 0.037 | -10.263 | 0 |
| Disturbance (high vs. low) | -0.276 | 0.062 | -2.671 | 0 |
| Time of day | 0.162 | 0.061 | 2.331 | 0.016 |
| Disturbance × time of day | -0.023 | 0.621 | -0.021 | 0.864 |
| Distance to livestock grazing trails | -0.121 | 0.133 | -1.667 | 0.074 |
| Distance to human trails | 0.46 | 1.015 | 0.453 | 0.543 |
| Distance to livestock corrals or huts | -1.856 | 2.701 | -2.126 | 0.037 |
Table 3: An averaging foraging distance travelled by adult marmots outside the pup burrows in relation to habitat stimuli in past
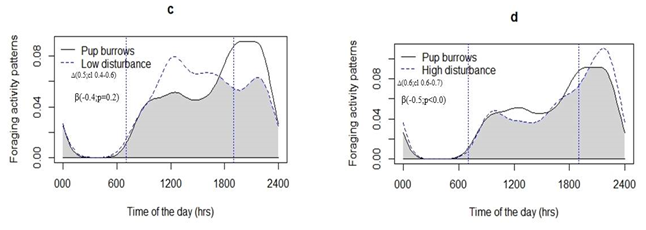
Gray shading represents the area of overlap using the coefficient of overlapping (‘overlap coefficient’∆) and cl (Confidence intervals), and Spearman rank correlation (SRC) between the adult marmots and two level of disturbance activities time and beta (coefficient), P represented significant level.
Discussion
Our results show a temporal adjustment in the activity pattern and foraging behaviors of golden marmots in relation to habitat disturbance around pup rearing burrows. While marmot activity levels and foraging distances were dissimilar between areas with high and low disturbances, there was an important positive or negative alteration in the scheduling and nature of their above-ground activity correlated to livestock grazing intensity or human stimuli (Tables 1& 2).
Effect of Habitat Disturbance on Diurnal Activity Shaping of Golden Marmots
Marmots reduced their temporal activity when herders and their livestock were in the area around the pup burrows, as also observed in our pilot research [15], especially during early morning and in the late evening when disturbances associated with pastoral activity reached a peak. In early mornings in pasture land, shepherds release their livestock includes zouno or cow from corrals for grazing. Moreover, in the case of three pup burrows, we found predatory attacks by Golden Eagle during early morning, but such predatory attacks were not found at other sampling time. Marmots were more active during times of the day when disturbances from humans and livestock were low, especially during the midday in both high and low disturbance sites. In our study area, livestock (especially cows or zou zouno) would take a rest in the midday in distinct bedding sites or leave the study sites for searching for drinking water due to heat or perspiration from sun exposure; at this time, shepherds also were involved in cooking or praying and resting inside the shepherd huts due to high sun exposure intensity in pastureland during midday. Consequently, we expected that the marmots take full advantage of this time for different activities. This behavioral adjustment suggests that marmots perceive disturbances associated with pastoral activity as a threat. Our findings are consistent with previous studies [7, 19], which have shown that animals can adjust their temporal activity patterns to avoid interactions with humans. Also, the bimodal activity pattern we noted for golden marmots has been commonly detected for other Marmota species: such as the hoary marmot (Marmota caligata) [58], yellow-bellied marmot [59], alpine marmot [60], arctic black-capped marmot (Marmota camtschatica bungei) [61] and Himalayan marmot (marmot in Himalayans [42]. Although numerous studies have described the nature of this bimodal activity pattern in relation to warm temperatures [1, 62], these conclusions are not suitable to our species because the temperature never reaches the described critical limit (21–27°C) in our study systems [63], thus we conclude other factors are at play.
Our results also show that golden marmots spend very little time out of their pup burrows as compared with other rodents [61, 64, 65]. Adult golden marmots averaged 65.83 % of the daylight hours above ground—approximately 4h (±38min) per day. In contrast, Belovsky and Slade, [47] reported that yellow-bellied marmots spend 6 h per day above ground, alpine marmots 7h per day [62, 66], and hoary marmots spend approximately 6h per day (70%) of their time above ground [64]. However, such assessments in activity are challenging because golden marmots share pastures with other livestock. For example, livestock existent were strongly effect on other marmot’s diurnal activities [46]. Therefore, we assumed adults spending less time away from burrows because they want to be inside protecting pups. For example, multiples opening in earthen burrows and rocky burrows were significantly increased in survival of herbivores in sub alpine pasture (E.g., pikas: Ochotona principals [67]. Furthermore, foraging also important for marmots because marmots have to conserve energy to survive through winter hibernation [68, 69]. The results indicate that marmots are conserving energy by reducing activity time, as energy expenditures during activity are greater than during rest in pup burrows [1].
Effects of Habitat Disturbance on Foraging Distances
We found that adult marmots foraging distance from pup burrows in relation to disturbance activities in high or low disturbance site. In sites experiencing high disturbances, marmots increased their mean foraging distances during in late afternoon and decreased during in the midday also morning time and increased in the evenings. We also found that disturbance activities time were strongly influenced on adult marmots foraging time and marmots forage much closer to their burrows during periods of high risk associated with herders, livestock are in the surrounding area (zaman personal observation).
For example foraging activity in other marmots influences by many factors, Marmots only produce alarm calls when comparatively near to forage their burrows [70], they forage more carefully if they run slowly [49, 71], they forage less when the visibility of their surroundings is shortened [72], and they forage a lesser amount of after hearing, smelling, or sighted [73, 74, 75] environmental stimuli or predators. The average foraging distance in high disturbance sites (13.8 ± 6.5 m) was similar to reports for hoary marmots in northern America (5–12 m; Karels, et al. [76]) and yellow-bellied marmotsin rocky mountain USA (<20 m; Frase and Armitage [77]). However, our findings are in contrast to those by Holmes [78] for hoary marmots in south-central Alaska (49.9 m) and for yellow-bellied marmots in California (up to 300 m (Arey and Moore [50]). Such differences are most likely associated with environmental conditions or habitat disturbances, such as Himalayan marmot (Marmota Himalayan) in Zoige wetland China [45] and further associated energy constraints, and predation risk such as golden marmots in Khunjerab national park northern Pakistan [25]. In this study, marmots exhibited short movement patterns for low disturbance due to rich food availability surrounding of pup burrows (zaman personal observation). Furthermore, marmots in high disturbance sites showed larger foraging distances, which could be because the study area provides forage for a large number of domestic livestock and the distance may be determined by risk associated with human activity or that marmots are selecting foraging sites further away in order to travel to less disturbed sites. Alternatively, livestock may facilitate quality food availability with browsing on shrubs [79]. Although food availability can affect the foraging movements, it appears unlikely to have caused the diurnal difference we observed in foraging distance because vegetation does not change according to a diurnal pattern. Because we selected both sites in the same valleys and same altitudes, there were no substantial differences in vegetation cover the habitat structure or availability of escape cover also can affect the perceived risk of predation, and hence foraging movements [80]. However, this study could not disentangle the potential effects of disturbance and habitat structure on foraging distance.
Conclusion
The Shigar valleys high altitude rangelands are in threatening condition due to excessive fuel wood collection, mining, flooding, overgrazing where resources are regularly limited. Therefore any prospective conflict between human activities and Wildlife may have a main influence on the ecology and persistence of species. For marmots, gaining sufficient food while decreasing the perceived risks associated with disturbance activities is principally significant, as they have to feed proficiently to achieve a critical body mass through the short summer feeding period to survive winter hibernation [25]. Our results propose that although the total daily foraging time is affected by disturbance activities, marmots exhibited increased levels of vigilance during foraging in areas experiencing high levels of disturbances [50]. Such as larger foraging distances travelled in less forages sites and increased in wariness of marmots in relation to disturbances, which may have harmful influences on the fitness of marmots. Further the study need to compare the vigilances and feeding time in relation to high or low disturbances sites and also need reproductive success or survival [19, 42].
Acknowledgements
Thanks to Nathan J. Roberts for assisting with editing and review of an earlier draft of this manuscript. This research was funded by the National Natural Science Foundation of China (NSFC31872241), and supported by the 111 Project (B20088) and Biodiversity Survey, Monitoring and Assessment Project of the Ministry of Ecology and Environment, China (2019HB2096001006).
References
-
Halle S, Stenseth NC (2000) Activity Patterns in Small Mammals: An Ecological Approach. 1st (Edn.), Springer Berlin, Heidelberg, pp: 322.
-
Creel S, Schuette P, Christianson D (2014) Effects of predation risk on group size, vigilance, and foraging behavior in an African ungulate community. Behavioral Ecology 25(4): 773-784.
-
Fenn MG, Macdonald DW (1995) Use of middens by red foxes: risk reverses rhythms of rats. Journal of Mammalogy 76(1): 130-136.
-
Fraser DF, Gilliam JF, Akkara JT, Albanese BW, Snider SB, et al. (2004) Night feeding by guppies under predator release: effects on growth and daytime courtship. Ecological Society of America 85(2): 312-319.
-
Harrington LA, Harrington AL, Yamaguchi N, Thom MD, Ferreras P, et al. (2009) The impact of native competitors on an alien invasive: temporal niche shifts to avoid interspecific aggression. Ecology 90(5): 1207-1216.
-
Schor NK, Dayan T (2003) Partitioning of time as an ecological resource. Annual review of ecology, evolution, and systematics 34(1): 153-181.
-
Schwartz CC, Cain SL, Podruzny S, Cherry S, Frattaroli L, et al. (2010) Contrasting activity patterns of sympatric and allopatric black and grizzly bears. Journal of Wildlife Management 74(8): 1628-1638.
-
Frid A, Dill L (2002) Human-caused disturbance stimuli as a form of predation risk. Conservation ecology 6(1).
-
Arroyo MG, Fors IM (2020) Tolerant to humans? Assessment of alert and flight initiation distances of two bird species in relation to sex, flock size, and environmental characteristics. Ethology Ecology & Evolution 32(5): 445-456.
-
Christiansen F, Rasmussen MH, Lusseau D (2013) Inferring activity budgets in wild animals to estimate the consequences of disturbances. Behavioral Ecology 24 (6): 1415-1425.
-
Houston AI, Prosser E, Sans E (2012) The cost of disturbance: a waste of time and energy? Oikos 121(4): 597-604.
-
Gill JA, Norris K, Sutherland WJ (2001) Why behavioural responses may not reflect the population consequences of human disturbance. Biological Conservation 97(2): 265-268.
-
Zaman M, Tolhurst BA, Zhu M, Jiang G (2020) Den- site selection at multiple scales by the red fox (vulpes vulpes subsp. montana) in a patchy human-dominated landscape. Global Ecology and Conservation 23: e01136.
-
Zaman M, Rakha BA, Wrensford K, Zhu M, Fan L, et al. (2020) Occurrence of conspecific in the neighboring space influence on flight initiation distance in Cape Hare Lepus capensis under human stimuli. Open Journal of Ecology 10(6): 380.
-
Zaman M, Tolhurst BA, Zhu M, Jiang G (2019) Increased Flight Initiation Distance (FID) in Golden Marmots (Marmota caudata aurea) Responding to Domestic Dogs in A Landscape of Human Disturbance. Animals (Basel) 9(9): 605.
-
Brown NA, Ruckstuhl KE, Donelon S, Corbett C (2010) Changes in vigilance, grazing behaviour and spatial distribution of bighorn sheep due to cattle presence in Sheep River Provincial Park, Alberta. Agriculture, ecosystems & environment 135(3): 226-231.
-
Reid RS, Bedelian C, Said MY, Kruska RL, Mauricio RM, et al. (2009) Global livestock impacts on biodiversity. In: Steinfeld H, et al. (Eds.), Livestock in a changing landscape. Drivers, consequences, and responses 1: 111- 137.
-
Welp T, Rushen J, Kramer DL, Bianchet MF, Passille AMBD (2004) Vigilance as a measure of fear in dairy cattle. Applied Animal Behaviour Science 87(1-2): 1-13.
-
Poudel BS, Spooner PG, Matthews A (2016) Pastoralist disturbance effects on Himalayan marmot foraging and vigilance activity. Ecological Research 31(1): 93-104.
-
Namgail T, Fox JL, Bhatnagar YV (2007) Habitat shift and time budget of the Tibetan argali: the influence of livestock grazing. Ecological Research 22(1): 25-31.
-
Valeix M, Jammes SC, Fritz H (2007) Interference competition and temporal niche shifts: elephants and herbivore communities at waterholes. Oecologia 153(3): 739-748.
-
Dark J (2005) Annual lipid cycles in hibernators: integration of physiology and behavior. Annu Rev Nutr 25: 469-497.
-
Kuhn KM, Wall SBV (2008) Linking summer foraging to winter survival in yellow pine chipmunks (_Tamias_ _amoenus_). Oecologia 157(2): 349-360.
-
Kuhn KM, Wall SBV (2009) Formation and contents of yellow-pine chipmunk (Tamias amoenus) winter larders. Western North American Naturalist 69(3): 309-318.
-
Blumstein DT, Arnold W (1998) Ecology and social behavior of golden marmots (Marmota caudata aurea). Journal of Mammalogy 79(3): 873-886.
-
Armitage KB (2012) Sociality, individual fitness and population dynamics of yellow-bellied marmots. Mol Ecol 21(3): 532-540.
-
Blumstein DT (1998) Quantifying predation risk for refuging animals: a case study with golden marmots. Ethology 104(6): 501-516.
-
Blumstein DT, Diaz A, Yin L (2018) Marmots do not consistently use their left eye to respond to an approaching threat but those that did fled sooner. Current Zoology 64(6): 727-731.
-
Zhang H, Li W, Hu Y, Zhang Y (2016) Opposite companion effect on flight initiation distance in sympatric species: plateau pika (Ochotona curzoniae) and White-rumped Snowfinch (_Onychostruthus taczanowskii_). Canadian Journal of Zoology 94(2): 109-114.
-
Childress MJ, Lung MA (2003) Predation risk, gender and the group size effect: does elk vigilance depend upon the behaviour of conspecifics? Animal Behaviour 66(2): 389-398.
-
Li C, Monclús R, Maul TL, Jiang Z, Blumstein DT (2011) Quantifying human disturbance on antipredator behavior and flush initiation distance in yellow-bellied marmots. Applied Animal Behaviour Science 129(2-4): 146-152.
-
Sproat KK, Martinez NR, Smith TS, Sloan WB, Flinders JT, et al. (2019) Desert bighorn sheep responses to human activity in south-eastern Utah. Wildlife Research 47(1): 16-24.
-
Armitage KB, Wolff J, Sherman P (2007) Evolution of sociality in marmots: it begins with hibernation. University of Chicago, Press Chicago, Illinois.
-
Borgo A (2003) Habitat requirements of the Alpine marmot Marmota marmota in re-introduction areas of the Eastern Italian Alps. Formulation and validation of habitat suitability models. Acta Theriologica 48(4): 557- 569.
-
Darimont CT, Fox CH, Bryan HM, Reimchen TE (2015) HUMAN IMPACTS. The unique ecology of human predators. Science 349(6250): 858-860.
-
Häfker NS, Raible KT (2020) Rhythms of behavior: are the times changin’? Curr Opin Neurobiol 60: 55-66.
-
Blumstein DT, Daniel JC, Bryant AA (2001) Anti‐predator behavior of Vancouver Island marmots: Using congeners to evaluate abilities of a critically endangered mammal. Ethology 107(1): 1-14.
-
Galluzzi M, Armanini M, Ferrari G, Zibordi F, Nocentini S, et al. (2017) Habitat Suitability Models, for ecological study of the alpine marmot in the central Italian Alps. Ecological Informatics 37: 10-17.
-
Anwar MB, Nadeem MS, Beg MA, Kayani AR, Muhammad G (2012) A photographic key for the identification of mammalian hairs of prey species in snow leopard (_Panthera uncia_) habitats of Gilgit-Baltistan Province of Pakistan. Pakistan J Zool 44(3): 737-743.
-
Shabbir S, Anwar M, Hussain I, Nawaz MA (2013) Food habits and diet overlap of two sympatric carnivore species in Chitral, Pakistan. Journal of Animal and Plants Sciences 23(1): 100-107.
-
McLean BS (2018) _Urocitellus parryii_ (Rodentia: Sciuridae). Mammalian Species 50(964): 84-99.
-
Poudel BS, Spooner PG, Matthews A (2015) Temporal shift in activity patterns of Himalayan marmot in relation to pastoralism. Behavioral Ecology 26(5): 1345-1351.
-
Berryman AA, Hawkins BA (2006) The refuge as an integrating concept in ecology and evolution. Oikos, 115(1): 192-196.
-
Abbas Z, Khan SM, Alam J, Khan SW, Abbasi AM (2017) Medicinal plants used by inhabitants of the Shigar Valley, Baltistan region of Karakorum range-Pakistan. Journal of ethnobiology and ethnomedicine 13(1): 53.
-
Zhou S, Krzton A, Gao S, Guo C, Xiang Z (2021) Effects of human activity on the habitat utilization of Himalayan marmot (_Marmota himalayana_) in Zoige wetland. Ecology and Evolution 11(13): 8957-8968.
-
Poudel BS, Spooner PG, Matthews A (2016) Behavioural changes in marmots in relation to livestock grazing disturbance: an experimental test. European journal of wildlife research 62(4): 491-495.
-
Belovsky GE, Slade JB (1986) Time budgets of grassland herbivores: body size similarities. Oecologia 70 (1): 53- 62.
-
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49(3): 227-267.
-
Blumstein DT, Runyan A, Seymour M, Nicodemus A, Ozgul A, et al. (2004) Locomotor ability and wariness in yellow‐bellied marmots. Ethology 110(8): 615-634.
-
Carey HV, Moore P (1986) Foraging and predation risk in yellow-bellied marmots. American Midland Naturalist 116(2): 267-275.
-
Meredith M, Ridout M (2021) Overview of the overlap package. R Proj, pp: 1-9.
-
Meredith M, Ridout M (2021) Package ‘overlap’: estimates of coefficient of overlapping for animal activity patterns. R package version 0.3. 3, pp: 1-20.
-
Linkie M, Ridout MS (2011) Assessing tiger–prey interactions in Sumatran rainforests. Journal of Zoology 284(3): 224-229.
-
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. Journal of Agricultural, Biological, and Environmental Statistics 14(3): 322-337.
-
Wikle CK, Mangion AZ, Cressie N (2019) Spatio-temporal Statistics with R. Boca Raton. FL: Chapman & Hall, CRC Press, pp: 396.
-
Pinheiro JC, Bates DM (2000) Linear mixed-effects models: basic concepts and examples. Mixed-effects models in S and S-Plus, pp: 3-56.
-
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, et al. (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol & Evol 24(3): 127-135.
-
Barash DP (1974) The social behaviour of the hoary marmot (_Marmota caligata_). Animal Behaviour 22 (1): 256-261.
-
Armitage K, Salsbury CM (2016) Pattern and variation of the time budget of yellow-bellied marmots. Ethology Ecology & Evolution 28(3): 329-365.
-
Perrin C, Allaine D, Berre ML (1993) Socio‐spatial organization and activity distribution of the alpine marmot Marmota marmota: Preliminary results. Ethology 93(1): 21-30.
-
Semenov Y, Ramousse R, Berre ML, Vassiliev V, Solomonov N (2001) Aboveground activity rhythm in Arctic black-capped marmot (Marmota camtschatica bungei Katschenko 1901) under polar day conditions. Acta Oecologica 22(2): 99-107.
-
Türk A, Arnold W (1988) Thermoregulation as a limit to habitat use in alpine marmots (_Marmota marmota_). Oecologia 76(4): 544-548.
-
Nikol’skii AA, Ulak A (2006) Key factors determining the ecological niche of the Himalayan marmot, Marmota himalayana Hodgson (1841). Russian Journal of Ecology 37(1): 46-52.
-
Taulman JF (1990) Late summer activity patterns in hoary marmots. Northwestern Naturalist 71(2): 21-26.
-
Everts LG, Strijkstra AM, Hut RA, Hoffmann IE, Millesi E (2004) Seasonal variation in daily activity patterns of free-ranging European ground squirrels (_Spermophilus_ _citellus_). Chronobiol Int 21(1): 57-71.
-
Boero DL (2001) Occupation of hibernacula, seasonal activity, and body size in a high altitude colony of Alpine marmots (_Marmota marmota_). Ethology Ecology & Evolution 13(3): 209-223.
-
Huntly NJ (1987) Influence of refuging consumers (pikas: _Ochotona princeps_) on subalpine meadow vegetation. Ecology 68(2): 274-283.
-
Arnold W, Heldmaier G, Ortmann S, Pohl H, Ruf T, et al. (1991) Ambient temperatures in hibernacula and their energetic consequences for alpine marmots _Marmota_ _marmota_. Journal of Thermal Biology 16 (4): 223-226.
-
Humphries MM, Kramer DL, Thomas DW (2003) The role of energy availability in mammalian hibernation: an experimental test in free-ranging eastern chipmunks. Physiol Biochem Zool 76(2): 180-186.
-
Collier TC, Blumstein DT, Girod L, Taylor CE (2010) Is alarm calling risky? Marmots avoid calling from risky places. Ethology 116(12): 1171-1178.
-
Blumstein DT, Ebensperger LA, Hayes LD, Vásquez RA, Ahern TH, et al. (2010) Towards an integrative understanding of social behavior: new models and new opportunities. Front Behav Neurosci 4: 34.
-
Bednekoff PA, Blumstein DT (2009) Peripheral obstructions influence marmot vigilance: integrating observational and experimental results. Behavioral Ecology 20(5): 1111-1117.
-
Blumstein DT, Recapet C (2009) The sound of arousal: The addition of novel non‐linearities increases responsiveness in marmot alarm calls. Ethology 115(11): 1074-1081.
-
Blumstein DT, Barrow L, Luterra M (2008) Olfactory predator discrimination in yellow‐bellied marmots. Ethology 114(11): 1135-1143.
-
Blumstein DT, Ferando E, Stankowich T (2009) A test of the multipredator hypothesis: yellow-bellied marmots respond fearfully to the sight of novel and extinct predators. Animal Behaviour 78(4): 873-878.
-
Karels TJ, Koppel L, Hik DS (2004) Fecal pellet counts as a technique for monitoring an alpine-dwelling social rodent, the hoary marmot in (_Marmota caligata_). Arctic, Antarctic, and Alpine Research 36(4): 490-494.
-
Frase BA, Armitage KB (1984) Foraging patterns of yellow-bellied marmosts: role of kinship and individual variability. Behavioral Ecology and Sociobiology 16(1): 1-10.
-
Holmes WG (1984) Predation risk and foraging behavior of the hoary marmot in Alaska. Behavioral Ecology and Sociobiology 15(4): 293-301.
-
Farnsworth KD, Focardi S, Beecham JA (2002) Grassland- herbivore interactions: how do grazers coexist? Am Nat 159(1): 24-39.
-
Adams JL, Camelio KW, Orique MJ, Blumstein DT (2006) Does information of predators influence general wariness? Behavioral Ecology and Sociobiology 60(5): 742-747.
- In Situ Evaluation of the Anthelmintic Effect of the Aqueous Extract of Syzygium aromaticum (L) Merr and Perry on Bovine Strongyles
- Successful in Vitro Embryo Production with Oocytes Aspirated from Live White-Tailed Deer (Odocoileus Virginianus Texanus) Donors under Captivity in Northeast Mexico
- Bangladeshi Finches with their Evolutionary Thoughts (Aves: Passeriformes)
- Note on the Survival Status of Przewalski’s Horse, Equus ferus przewalskii (Perissodactyla: Equidae)
- In-Situ and Ex-Situ Protection of White-Breasted Waterhen (Amaurornis Phoenicurus) (Pennant, 1769) (Aves: Rallidae)
- Meat Examination in the Laboratory, the Acceptability and the Human Health