Non Human Animals Responses to Social Loss
An emerging topic of interest for those studying the effects of social loss and separation in non-human animals converges upon whether non-human animals experience the negative emotions of losing an attachment figure to death. Following screening for inclusion criteria, we systematically reviewed 40 scientifically documented accounts of grief-like reactions around loss, primarily involving mother and young but also including several unrelated adults, in various animal taxa including primates, marine mammals, elephants, a canid species, and an avian species. Most accounts involved animals in the wild, but animals in captivity were also included. We summarized information about behavioral frequencies and durations and physiological responses before and after the loss, causes of loss, time lived together, and observation period. Distress behaviors such as self-grooming, allogrooming, grooming the carcass, and vocalizations were the most reported behaviors, followed by carrying, physical contact, and staying with the dead. Based on this body of evidence, we conclude that at least some non-human animals can experience grief-like emotions, making grief not exclusively a human phenomenon and, thus, worthy of animal welfare concern.
Fanucchi L¹* and Newberry RC²
¹College of Veterinary Medicine, Oklahoma State University, Stillwater, USA ²Department of Animal and Aquacultural Sciences, Norwegian University of Life Sciences, Norway *Corresponding author: Leticia Fanucchi, College of Veterinary Medicine, Oklahoma State University, 1118 Lauren Ln, Stillwater, Ok, USA, Phone: 4059925955; Email: le.fanucchi@okstate.edu Keywords: Animal; Attachment; Emotions; Grief; Loss; Welfare
Introduction
An emerging topic of interest for those studying the effects of social loss and separation in non-human animals converges upon questions such as: Can non-human animals grieve the loss of a familiar individual? Do they form emotional attachments? Is an understanding of the concept of death necessary for grief to be experienced by animals? [1, 2]
In humans, perhaps the most traumatizing life experience is the death of an attachment and grieving is the normal response to the loss [3]. For bereavement to be considered as a process resulting from loss, three key elements are necessary: an attachment figure that can potentially satisfy an individual’s needs, loss of the attachment figure, and the remaining individual who is deprived of the attachment [4]. Grief in humans has also been described to have five distinct stages that reflect the emotional phases experienced by the bereaved individual: denial (unacceptance that a loss occurred), anger (frustration that the lost figure is not coming back), bargaining (negotiating the loss, as if this was possible), depression (physical and psychological pain, realization that the loss has occurred), and acceptance (moving on after recovery) [5]. Bowlby, proposed that grief develops in four phases: shock, protest, despair, and recovery. Similarly, Parkes, proposed four stages including numbness, pining, disorganization, and reorganization. Ultimately, all behavior described in the models overlaps, and ends in the individual moving on with life [6, 7].
For grief to be present, an attachment bond needs to exist towards the lost individual [4]. Attachment is a social fondness that develops between individuals and is characterized by voluntary contact and proximity maintenance [8]. Care of infants is a prime target for the emergence of attachment bonding, with attachment of young occurring to one or more caregivers, and bonds developing between caregivers that can extend beyond the raising of one infant or brood. Mammalian parental behavior and pair bonding are based on homologous physiological mechanisms, with similar mechanisms in birds, and species differences arising from differences in the degree of expression of these mechanisms [9, 10]. Thus, for example, in monogamous prairie voles (Microtus ochrogaster), unrelated adults of opposite sex are highly prone to forming strong partner preferences even in the absence of mating [11] whereas polygynous montane voles (M. montanus) do not form enduring, selective attachment bonds. Social choices are promoted by neuropeptides such as oxytocin [12] and vasopressin [13], which have been shown to play a critical role in rodent pair bond formation [14], and species differences are influenced by the levels of oxytocin and vasopressin receptor expression in brain areas involved in affiliative behavior [15].
In humans, when an attachment figure is absent, the usual immediate distress response to loss is characterized by a “protest phase” [6], often marked by cries. The strong protest vocalizations represent a state of PANIC generated by hormones and brain circuits [16]. Separation-induced vocalizations can be blocked by opiates and induced by opiate antagonists [17, 18], highlighting the role opiates play in regulating this response in a mother-infant separation paradigm. Dog puppies (Canis lupus familiaris), when separated from their mother or siblings, display separation protest behaviors characterized by loud vocalizations, which seem effective in reuniting the youngster with the mother and littermates [19]. Interestingly, this protest behavior characteristic of separation distress is also displayed in dogs when left alone in an unfamiliar environment, away from human companions, which supports the premise that cross species attachments can be formed [20, 21, 22], and that bonding to a human can result in safety and survival [23, 24]. Distress behavior when threatened by separation from an attachment figure appears to represent an evolutionary survival strategy shared by vertebrates [9, 25].
When the “protest phase” does not result in reunion with the missing figure, a “despair phase” [26] can set in, characterized by a depressed state, which is often marked by reactions that can greatly affect an individual’s function, such as restlessness, loss of appetite, and decreased activity [27]. In humans, social loss is a major factor contributing to the onset of depression [28]. Prolonged bereavement is currently considered a depressive disorder and is included in the diagnostic criteria for depression in the 5th edition of the Diagnostic and Statistical Manual of Mental Disorders, DSM- 5 [29]. Because the architecture of the core affective network that modulates the neural activity of emotions appears to be shared across mammals [30, 31], depressive states following loss can be predicted to extend beyond humans.
In sum, from neurophysiological and behavioral points of view, there is growing evidence supporting the premise that mammals and birds, if not all vertebrates are equipped to experience at least to some degree, the distressing effects of being separated from peers, regardless of the cause of separation. The brain mechanisms are homologous across mammalians [32], extending to birds and possibly other vertebrates.
In this paper, our overall hypothesis is that non-human animal reactions around loss of a significant other due to death involve an active process, similar to the grief process described in humans, with affective (emotional distress), and cognitive (coping with the loss, learning and moving on) elements, that have an adaptive value and serve a survival function. Despite a massive number of anecdotal and scantily described accounts of grief-like symptoms in mammals and birds, some reports include precise and detailed observations of animal behavior around the death of a group member, with careful documentation that includes video recording and/or photographs before and after the loss, allowing portrayal as case reports. These cases will be discussed here in terms of proximate consequences of loss in captive and wild animals. Given this body of evidence on effects of separation, we discuss evidence for grief in mammals and birds based on commonalities in behavior patterns exhibited in a social loss context across a variety of species.
Specific Hypotheses and Predictions
Affective Components of Social Loss
Hypothesis 1: The reactions around social loss are representative of activation of brain circuitry involved in the primary process, more specifically the PANIC/separation system [8], that characterize the “protest phase” [6]. Prediction 1: After a social loss, animals undergo stages similar to the five stages of grief process occurring in humans [5], characterized by increased vocalizations (anger), ceased play (depression), loss of appetite (depression), erratic movements (anger), aggressive guarding of the carcass from other group members (anger) when applicable, inspection of the carcass (denial), aggression towards the carcass (anger), grooming the carcass (denial), showing increased self-grooming (depression), allogrooming, staying in close proximity to the carcass (denial), and increased activation of the hypothalamic-pituitary-adrenal axis (anger, denial, depression), when applicable. Hypothesis 2: A small number of animals enter a prolonged depressive state (similar to complicated grief in humans) and do not recover to normal function. Prediction 2: Some animals do not recover and perish, some cases ending in death.
Cognitive Components of Social Loss
Hypothesis 3: The prolonged reactions after a social loss are representative of activation of brain circuitry composing the secondary and/or tertiary processes responsible for the affective aspect of associative learning. Prediction 3: Most animals recover, and abandon the carcass, indicative of the last grief stage (acceptance).
Environmental and Social Aspects of Social Loss
Hypothesis 4: Responses around social loss vary depending on animals’ social environment (pairs vs larger groups), reproductive status, gender, living condition (wild or captive), and social relationships (genetically related to the dead or not, familiar/unfamiliar to the dead). Prediction 4: Most reactions around loss are reported in animals that live preferentially in pairs (in close proximity, sharing resources, of the same or different species, wild or captive), without much opportunity for coping with the loss through seeking contact with other group members. Prediction 5: The majority of the cases reporting protest behavior following a loss involve adult females (especially intact, lactating mammals), in comparison with adult males. Prediction 6: Most of the reports involve genetically related animals. Prediction 7: Most of the reports involving genetically related animals concern parent-offspring dyads, such as where a mother loses her young.
Materials and Methods
This review focuses on retrospective studies published in scientific journals, books, and other reports from credible sources from 1879 to 2012, describing behaviors occurring around loss due to death, some of which will be outlined here. Because of variation in depth of reporting, insufficient papers were found to conduct meta-analyses. Instead, we report numbers of studies reporting parameters of interest. Studies were included if they reported descriptions or data (durations and/or frequencies) on behavioral and/or physiological responses before and/or after a loss. Cause of the loss, time together before the loss, and duration of observation were also noted if presented.
Of forty-seven studies in the literature, n = 40 studies fulfilled our inclusion criteria. Of all the behaviors reported, we selected the ones that were most commonly reported and defined them (Table 1).
We classified the reports according to the animals’ taxonomic group, species, living condition, group size, relatedness to lost individual, age of the subject (who lost the individual), sex of the subject, lost individual’s condition (stillborn, newborn, infant, juvenile, adult), and reason for loss, and a summary of all accounts is presented (Appendix 1).
Results
Categorization of Reports
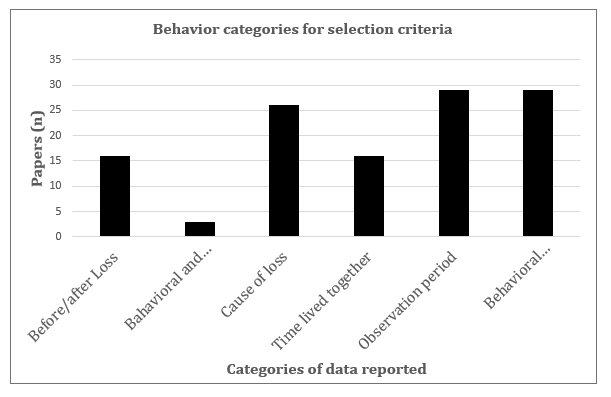
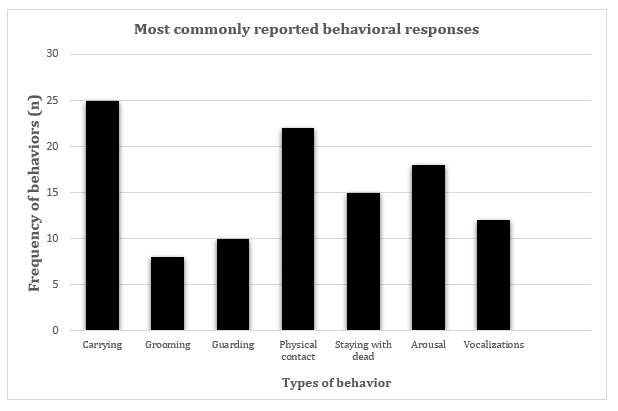
Twenty one of the 40 selected studies were on primate species, n = 15 in marine mammals, n = 2 in elephants, n = 1 in a canid species, and n = 1 in an avian species. Thirty- one were observations of animals in the wild, and n = 9 of animals in captivity. Before and after the loss information was provided in 42% of the papers, physiological as well as behavioral information in 7.5%, cause of the loss in 65%, time lived together in 40%, observation period in 75%, and duration and/or frequency of behaviors in 72.5% (Figure 1). Behavioral descriptions were provided in 75% of the papers. The number of papers reporting the categorized behaviors (Table 1) is shown in Figure 2.
Many of these behaviors were reported in different species. Carrying the dead was commonly reported (65% of the papers), as were physical contact (55%), and staying with the dead (37.5%). Other predicted reactions (Prediction 1) included: Aggression (25.6% of the papers), grooming the carcass (20.5%), self-grooming (97.4%), and allogrooming (12.8%).
| Behavior | Definition |
|---|---|
| Carrying | Carrying, pushing, supporting, pulling or dragging the dead body |
| Grooming | Grooming, licking, fly swatting, nibbling, biting nails, or cleaning the dead body |
| Guarding | Any aggressive behavior directed towards other members of the group, other animals, or people approaching the dead body |
| Physical contact | Any physical contact with the dead body not including grooming and carrying |
| Stay with the dead | Staying in close proximity with, tending, or sleeping by the dead body |
| Vocalizations | Any vocal sound emitted in the presence of the dead body, including calls, barks, whimpers, whines, screams, cries, rumbles, trumpets, growls, or whistles |
| Arousal | Any high-arousal, non-playful behaviors that appear to be specific to the loss context excluding vocalizations and guarding. Includes milling, swimming erratically, jumping, running around, shaking, nodding head, erections |
Table 1: Ethogram of commonly reported behaviors occurring in the context of death of a conspecific.
Only n = 2 (5.9%) cases of death following a social loss were reported (Prediction 2), and most of the animals recovered and abandoned the carcass n = 19 (82.6%), as in Prediction 3. Social condition included n = 20 cases of animals living in pairs (51.3%), and n = 19 (48.7%) reports of animals living in groups of more than two animals (Prediction 4). Contrary to Prediction 4, vocalizations following loss were reported equally often in cases involving animals living in pairs and in larger groups. Most of the case reports involved responses in adult females (n = 21) whereas n = 10 reports involved both males and females in mixed groups, and only n = 1 case concerned a male, as predicted (Prediction 5). Most of the accounts were on genetically related animals n = 25 (83.3%), as predicted (Prediction 6). Out of all accounts, n = 27 reports involved a parent who lost offspring, as in Prediction 7.
Selected Descriptive Summaries of Responses to Social Loss
Reports in Primates
Probably the first detailed documentation of grief- like reactions in primates in the wild was on a chimpanzee species. Brown AB, [33] described his observations on a captive male chimpanzee (species not specified) who lost a male companion, and showed distress signs for two days. Behavior was described as attempts to arouse the dead body, rage episodes, hair pulling, screams and “wraah” calls, moaning, lifting up arms and hands of the corpse, pushing and rolling body, and aggression towards the keeper when moving the body, on the first day of the observation period. On the second day, the solitary male showed clinging behavior towards the keeper, separation anxiety when left by the keeper, and a change in sleeping location.
Goodall J [34] reported one case in wild chimpanzees (Pan troglodytes), where a presumed mother lost her infant from a predatory attack, and carried the corpse for seven days, showing aggression to group members approaching the dead infant, especially one member who had previously killed one of her infants. Goodall reports that a group of chimpanzees that she followed for 18 years showed distress behaviors after the loss of offspring/siblings. The group members showing reactions to the loss consisted of eight females and five males. The female mothers showed extinguished play and carrying the corpse. The female siblings showed physiological changes such as lethargy, potbelly, delayed signs of adolescent sex swelling, until age 13 years in one female, and three females died after two weeks, seven weeks, and nine months, respectively. Out of the five males, responses such as a severe decrease in play, decreased social interactions, rocking, hair pulling, poor tool usage, hanging upside down for a prolonged period, whimpering when left alone and aggression towards siblings. One male showing lack of interest in females until age 7 years, and nervousness around males were observed. Some physiological responses included lethargy, sunken eyes, potbelly, delayed scrotum development, loss of appetite, gradual emaciation, and death of one male after three weeks [35].
Teleki witnessed the death of an adult male chimpanzee (P. troglodytes) in the wild, who fell off a tree and broke his neck in front of his group. Females (N=3) and males (N=13) showed distress behaviors for 3 hours, which included raucous calls, stamping the ground, tearing and dragging vegetation, swaggering bipedally, stone throwing, staring at the corpse, grins, mounting, touching and patting one another, standing in a circle around the body, pant hoots, and grooming each other. One male stayed with the corpse for 11 minutes after the group left before finally rushing after the group.
Other cases involving parents and loss of offspring in chimpanzees include similar behaviors, such as carrying, guarding and grooming the body, sleeping with the body and distress upon separation, staying with the body, fly swatting, gazing at the corpse, sniffing, touching, inspecting the body, and even playing with the dead infant [36, 37, 38, 39, 40].
A curious case of cross-species grief-like reactions was reported on the acclaimed captive female gorilla (Gorilla beringei grauer), called Koko, who used her ability to use sign language to communicate with her handlers [41]. Koko seemed to be attached to a kitten called All Ball, who died after being hit by a truck. When Koko was told about the death of the kitten, through sign language she expressed sadness, hand signals indicative of crying, frowning, she was found moaning in her enclosure and went to a sad state every time she was shown a photograph of the All Ball for up to five years following the death.
Gorillas are not the only great ape species where the use of sign language to express emotions has been reported. Psychologist Roger F, et al. [42] described an occurrence in a captive female chimpanzee (P. troglodytes) who lost her infant, born with a congenital heart defect. After the loss, the female had loss of appetite, was moping around the cage, and for two months following the loss, she used sign language to ask about the missing baby.
Three cases involving four wild female gorillas (Gorilla beringei beringei) have been described by Warren Y, et al. [43]. The first case involved a primiparous mother who lost an infant at age 36 days, and the second, an unrelated primiparous adult female who lost her 13-day-old infant. Both carried their dead infant intermittently for 15 days until they discarded it. During this period, they showed decreased interaction with other group members, decreased play behavior, and aggression to other family members approaching the corpse. The same responses were seen in a third mother who carried a dead infant for 20 days.
In one case of infanticide in wild black capuchins (Cebus nigritus), the newborn was killed in the presence of the mother who for one day was seen carrying the corpse and allowing female members of the group to touch and inspect the body [44]. In a case of witnessed jaguar attack, white- bellied spider monkeys (N=4) showed distress behaviors such as mobbing calls, shaking branches during the attack, following the jaguar after attack, returning to a tree near the wounded victim, staring at the victim, and grooming each other for twenty minutes [45]. Similar behaviors have been described in wild groups of gelada monkeys (Theropithecus gelada), and Japanese macaques (Macaca fuscata) [46], as well as in a captive group of black snub-nosed monkeys (Rhinopithecus bieti) [47].
Reports on Marine Mammals
Most observations reported in marine mammals involve groups of dolphins in the wild, where cause of death and relatedness to the lost individual is usually unknown. However, two cases involving captive dolphins have been reported with more detailed observations on the reactions after a loss occurred. One of the cases involved a female pacific dolphin (Delphinus bairdii), who delivered a stillborn [48]. The mother was seen carrying the corpse at the surface, until she witnessed the dead calf being swallowed by a pilot whale. The mother showed distress behaviors like whistling and swimming in erratic manner. McBride A, et al. [49] reported several stillbirth cases in captive female bottlenose dolphins (Tursiops truncatus), where carrying and supporting the dead calf at the surface was consistently observed.
Detailed observations involving two cases of wild bottlenose dolphins (Tursiops aduncus), were reported by Dudzinski KM, et al. [50] describing behaviors of males who lost a female companion (unknown relatedness). In one case where an adult female was found dead, the males (N=2) stayed with the carcass at all times for sixteen hours, alternating respiration, and showed aggression towards human swimmers attempting to retrieve the carcass. The second case involved a group (N=19 males and 2 females) where one dolphin drowned. The group showed distress behaviors for three hours, which involved vocalizations (echolocation, loud pops, whines and whistles, bubble stream), whipping the tail at peers, fast circle swims, parallel formation, inspection of carcass (touching body parts), guarding (aggressive behaviors towards males, and towards swimmers when attempting to retrieve carcass), penile erection, staying with the carcass, and remaining in the area after carcass retrieval.
The most common epimeletic behaviors in presumed parents and offspring of wild dolphins include close physical contact with the dead, pushing/carrying corpse, guarding carcass, slowed travel, and staying with the dead [51, 52, 53, 54, 55, 56, 57, 58, 59, 60]. Additionally, milling, and high-pitched vocalizations, pulling by fin, and erratic movements have also been documented [61].
Reports on Elephants
A detailed case involving the death of a female adult elephant (Loxodonta africana), the matriarch of a group of wild elephants, and the reactions of her relatives and her daughter after her passing was reported by Douglas-Hamilton and collaborators [62]. The authors had been observing the group for seven days before the death, and distress behaviors were seen up to 32 hours after the female’s passing. The offspring nuzzled the carcass, walked around confused, and stayed with it. The relatives approached, sniffed and touched the body, standing with it for long periods, guarding it, while attempting to lift it with foot/trunk/tusks. They also displayed screams, rumbles, and trumpets. The daughter associated with other females but was recorded missing (dead) within 3 months of her mother’s death.
Bone remains of elephants, bones of other wild animals, and neutral objects were presented to free ranging African elephants in a study conducted by McComb K, et al. [63]. The elephants spent significantly more time investigating the bones of elephants, regardless of the remains being from related or unrelated elephants, compared to other animals and objects, by touching and sniffing the bone remains.
Report on a Canid Species
An adult female dingo (Canis dingo) and her four pups (approximately three months old) were observed during the passing of one of her pups [64]. The pup was in distress for about 30 minutes (vocalizations and convulsion) and ended up passing away. The following day, the mother and two pups approached the dead pup. Then the mother picked it up and carried it for about 30 meters, dropped it, and stayed close to it for an unknown period (observation was suspended after 8 minutes).
Report on an Avian Species
A systematic study was conducted by Iglesias T, et al. [65] on wild Western scrub-jay (Aphelocoma californica) vocalizations, and the authors hypothesized that scrub-jays could use dead conspecifics as cues of risk and communicate the risk to others. The authors predicted that the jays would display risk-reducing behaviors, such as decreased foraging, and cacophonous aggregations in the area in response to a predator or a dead conspecific, but not to a novel object. Birds were presented with painted blue pieces of wood, a mounted predator, a mounted jay, and a scrub-jay carcass. As predicted, cacophonous aggregations and decreased foraging occurred in response to the dead jay and predator, but never to the object. Additionally, playback of jay vocalizations attracted jays to all sites where stimuli were presented [65].
Discussion
The most commonly observed behaviors included those interpreted to reflect separation distress, such as dazed walking, pacing, erratic movements (including swimming), shaking, rocking, stone throwing, redirected aggression (towards siblings, group members, and the environment), dragging vegetation, excessive grooming, hair pulling, and hanging upside down, typically seen in the protest phase (Prediction 1).
Complicated depression, characterized by loss of appetite, decreased activity, and decreased play was also observed. In two cases, death believed to be a result of the loss was reported. One of the cases involved the death of one female chimpanzee offspring two weeks after losing the mother [35], and another one involved the death of a young male who died twenty-four hours after the mother’s passing. In this last case, the young male chimpanzee was seen vocalizing, he stayed with the dead body, showed distress behaviors (rocking side to side), staying at a profound despair state [26], followed by death (Prediction 2). Even though we cannot tease out the cause of the two deaths, the two cases are similar to what has been observed in human adolescents after the loss of a parent, where complicated grief that does not resolve ends in compromised mental health, or more rarely in deat [66, 67].
Several of the behaviors observed in animals to a certain extent can be placed in the five grief stages, such as: distress behaviors and vocalizations resemble a state of anger and denial, where the protest phase is intense immediately after the loss; carrying the dead resembles a state of denial, where the animals have difficulty letting go off the carcass, especially during decomposition, when facial features are lost, and the decay smell is not of a living being; staying with the dead, restlessness, decreased activity, and loss of appetite are analogous to the depressed grief stage, where animals appeared to have lost interest in seeking for resources; abandoning the carcass, and following the group is comparable to acceptance (Prediction 3). This is not an attempt to anthropomorphize the behaviors displayed by animals, but to bring the discussion to the next level, if we accept that non-human animals can experience grief-like emotions. Animal grief is not only a public belief, especially on highly cognitive species, but also a feasible phenomenon, given that animals are all equipped with corresponding neural and hormonal mechanisms [68].
About half of the accounts involved animals living in pairs, and vocalizations were observed equally in animals living either in pairs or in groups, contrary to Prediction 4. Distress vocalizations from separation, such as high-pitched calls, rumbles, screams, whines, whistles, and moaning were among the most observed behaviors among all reports. Vocalizations from separation in animals is more a result of internal processes, regardless of social circumstances around the “grieving” individual [69]. Even though animals living in groups may have more opportunities to form multiple attachment bonds, it seems that the quality of the group is more important than the size of a group. In humans, social support from family and friends aids in the emotional healing process during bereavement [70]. However, people living in a dysfunctional social network need more professional help than those living in socially stable households/environments, and the size of the social network does not seem to be relevant when it comes to coping mechanisms [71].
Carrying, physical contact, and staying with the dead were also frequently reported by most of the papers. Carrying was mostly done by mothers, who were seen carrying their dead infant for several days, even after advanced carcass decomposition. Cases involving females (Prediction 5) accounted for most of the reports, as documented in humans [72, 73]. Staying in proximity to the dead and making physical contact were observed mostly in genetically related animals (Prediction 6), but also in non-related individuals, showing that breaking bonds can be a long lasting and an emotionally difficult process.
The great majority of the reports of grief-like reactions in animals involve a variety of relatively long-lived, highly cognitive social mammalian species, in a parent-offspring context, usually the mother of a newborn (Prediction 7). Maternal attachment is a common innate system mammalian species, as well as birds, having a survival value in altricial species. Maternal care (CARE system) is triggered by changes in neuropeptides such as oxytocin, prolactin, and estrogens [74]. Mothers have a unique neural response to their own offspring and the brain areas involved in parenting are akin in mammal species [75, 76].
Many of the separation distress behaviors occur in avian species as well, and may involve unrelated individuals, including cross-species. It appears that the losses involving non-human animals fit the bereavement criteria [4], and the behaviors observed are very similar to grief in humans, where the loss represents the disruption of a social structure, characterized by behavioral and physiological separation reactions. Nevertheless, it is important to clarify that losses do not have to involve death, or the understanding of death concept. Attachments can include a person, an animal, an object, or even a social status. In sum, an attachment is anything to which an emotional bond has been formed [77].
Several papers reported behaviors both before and after loss, cause of loss, the observation period and duration of behaviors, as well as time lived together. A few papers provided physiological measures. This type of information is important in ruling out other possible causes for behavior changes, not related to loss. Because the great majority of the papers only provided behavioral information, our strict criteria for selecting the papers discussed here allowed us to carefully extract the relevant information around loss, instead of other phenomena.
We understand the limitations in making conclusions based on behavioral observations alone, and emphasize that other measurements such as hormonal changes, heart rate, body temperature, food intake, among many others, should be included in future studies because they can provide valuable information, adding validity and reliability to qualitative research [78, 79, 80, 81, 82, 83, 84, 85, 86].
Animal Welfare Implications
Commonalities in the behavior report across multiple highly cognitive animal species demonstrates a weight of evidence that death of close relatives and social partners can have emotional consequences for survivors. Knowledge of responses to social loss has animal care implications because animals living in captivity sometimes need to be separated and may grieve the loss of a companion and/or human caregiver. The psychological and physiological consequences of the loss have the potential to impair animals’ wellbeing.
Conclusion
Based on our past review of the behaviors occurring around social loss due to death, we conclude that at least some non-human animal species can experience grief-like emotions, making grief not exclusively a human phenomenon.
Acknowledgments
We thank Hayley Clark, who assisted with the selection of papers. The authors state that there are no conflicts of interest.
References
-
Marshall M (2010) Bereaved animals grieve - if their lifestyle allows it. New Scientist 208: 12.
-
King B (2013) How Animals Grieve. The University of Chicago Press, UK.
-
Brasted WS, Callahan EJ (1984) A behavioral analysis of the grief process. Behavior Therapy 15: 529-543.
-
Corr CA, Coolican MB (2010) Understanding bereavement, grief, and mourning: Implications for donation and transplant professionals. Progress in Transplantation 20: 169-177.
-
Kubler-Ross E (1969) On Death and Dying. Macmillan, USA.
-
Bowlby J (1979) The Making and Breaking of Affectional Bonds. 1st (Edn.), Tavistock Publications, UK.
-
Parkes C (1998) Bereavement in adult life. BMJ (Clinical Research Ed) 316: 856-859.
-
Bowlby J (1969) Attachment and Loss: Basic Books, USA. 1: 296-375.
-
Panksepp J, Nelson E, Bekkedal M (1997) Brain systems for the mediation of social separation-distress and social- reward - Evolutionary antecedents and neuropeptide intermediaries. Annals of the New York Academy of Sciences 807: 78-100.
-
Kingsbury MA, Goodson JL (2014) Pair bond formation is impaired by VPAC receptor antagonism in the socially monogamous zebra finch. Behavioural Brain Research 272: 264-268.
-
Insel TR, Hulihan T (1995) A gender-specific mechanism for pair bonding: Oxytocin and partner preference formation in monogamous voles. Behavioral Neuroscience 109: 782-789.
-
Williams JR, Catania KC, Carter CS (1992) Development of partner preferences in female prairie voles (Microtus ochrogaster): the role of social and sexual experience. Hormones and Behavior 26: 339-349.
-
Winslow J, Hastings N, Carter CS, Harbaugh C, Insel T (1993) A role for central vasopressin in pair bonding in monogamous prairie voles. Nature 365: 545-548.
-
Donaldson ZR, Young LJ (2008) Oxytocin, vasopressin, and the neurogenetics of sociality. Science 322(5903): 900-904.
-
Young LJ (1999) Oxytocin and vasopressin receptors and species typical social behaviors. Hormones and Behavior 36: 212-222.
-
Newman JP (2007) Neural circuits underlying crying and cry responding in mammals. Behavioural Brain Research 2: 155-165.
-
Panksepp J, Meeker R, Bean NJ (1980) The neurochemical control of crying. Pharmacology Biochemistry and Behavior 12: 437-443.
-
Kalin NH, Shelton SE, Barksdale CM (1988) Opiate modulation of separation-induced distress in non- human primates. Brain Research 440: 285-292.
-
Cohen JA, Fox MW (1976) Vocalization in wild canids and possible effect of domestication. Behavioral Processes 1: 77-92.
-
Hatkoff I, Hatkoff C, Kahumbu DP (2006) Owen and Mzee. Scholastic Press, USA.
-
Izar P, Verderane MP, Visalberghi E, Ottoni EB, Gomes De Oliveira M, et al. (2006) Cross-genus adoption of a marmoset (Callithrix jacchus) by wild capuchin monkeys (Cebus libidinosus): Case report. American Journal of Primatology 68: 692-700.
-
Buckley C (2009) Tarra & Bella: The Elephant and Dog Who Became Best Friends. Putnam, USA.
-
Canas S, Frank D, Minero M, Godbout M, Palestrini C (2010) Puppy behavior when left alone: changes during the first few months after adoption. Journal of Veterinary Behavior 5: 94-100.
-
Steiner ET, Silver NC, Hall P, Downing C, Hurton D, et al. (2013) Raising canine: Cross-species parallels in parental investment. Human-Animal Interaction Bulletin 1: 38-54.
-
Carter C, DeVries A, Getz L (1995) Physiological substrates of mammalian monogamy: The prairie vole model. Neuroscience and Biobehavioral Reviews 19: 303-314.
-
Bowlby J (1980) Loss: Sadness and Depression: Vol. 3. Basic Books, USA.
-
Bowlby J (1960) Grief and mourning in infancy and early childhood. Psychoanalytic Study of the Child 75: 9-52.
-
Watts F (1993) Neuropsychological Perspectives on Emotion. Cognition and Emotion 7: 43-69.
-
Mojtabai R (2011) Bereavement-related depressive episodes: characteristics, 3-year course, and implications for the DSM-5. Archives of General Psychiatry 68: 920-928.
-
Coenen VA, Schlaepfer TE, Maedler B, Panksepp J (2011) Cross-species affective functions of the medial forebrain bundle: Implications for the treatment of affective pain and depression in humans. Neuroscience and Biobehavioral Reviews 35: 1971-1981.
-
Panksepp J, Normansell LA, Herman B, Bishop P, Crepeau L (1988) Neural and neurochemical control of the separation distress call. In: Newman JD (Ed.) The Physiological Control of Mammalian Vocalizations. Plenum, USA, pp: 263-300.
-
Panksepp J (2005) Affective consciousness: Core emotional feelings in animals and humans. Consciousness and Cognition 14: 30-80.
-
Brown AE (1879) Grief in the chimpanzee. The American Naturalist 13: 173-175.
-
Goodall J (1977) Infant killing and cannibalism in free- living chimpanzees. Folia Primatologica 28: 259-282.
-
Goodall J (1986) The Chimpanzees of Gombe: Patterns of Behavior. Belknap Press of Harvard University Press, USA, pp: 102-103.
-
Matsuzawa T (1997) The death of an infant chimpanzee at Bossou, Guinea. Pan Africa News 4: 4-6.
-
Hanamura S, Kiyono M, Braum ML, Mlengeya T, Fujimoto M, et al. (2000) Reactions to dead bodies of conspecifics by wild chimpanzees in the Mahale Mountains, Tanzania. Primate Research 16: 1-15.
-
Kooriyama T (2009) The death of a newborn chimpanzee at Mahale: Reactions of its mother and other individuals to the body. Pan Africa News 16: 19-21.
-
Biro D, Humle T, Koops K, Sousa C, Hayashi M, Matsuzawa T (2010) Chimpanzee mothers at Bossou, Guinea carry the mummified remains of their dead infants. Current Biology 20: 351-352.
-
Cronin KA, Van Leeuwen EJC, Mulenga I, Bodamer MD (2011) Behavioral response of a chimpanzee mother toward her dead infant. American Journal of Primatology 73: 415-421.
-
Patterson F, Gordon W (1993) The Case for the Personhood of Gorillas. In: Cavalieri P, Singer P (Eds.) The Great Ape Project. St Martin’s Griffin, USA, pp 58-77.
-
Fashing P, Barry TS, Goodale CB, Burke RJ, Jones SC, Kerby JT, Lee LM, Fouts RS, Mills ST (1997) Next of Kin. William Morrow, USA, pp: 215-233.
-
Warren Y, Williamson EA (2004) Transport of dead infant mountain gorillas by mothers and unrelated females. Zoo Biology 23: 375-378.
-
Izar P, Silva EDR, De Resende BD, Ottoni EB (2007) A case of infanticide in tufted capuchin monkeys (Cebus nigritus). MastozoologIa Neotropical 14: 73-76.
-
Matsuda I, Izawa K (2008) Predation of wild spider monkeys at La Macarena, Colombia. Primates 49: 65-68.
-
Sugiyama Y, Kurita H, Matsui T, Kimoto S, Shimomura T (2009) Carrying of dead infants by Japanese macaque (Macaca fuscata) mothers. Anthropological Science 117: 113-119.
-
Tengfei L, Baoping R, Dayong L, Yunbing Z, Ming L (2012) Maternal responses to dead infants in Yunnan snub- nosed monkey (Rhinopithecus bieti) in the Baimaxueshan Nature Reserve, Yunnan, China. Primates 53: 127-132.
-
Brown DH (1966) Observations on the behavior of wild and captive false killer whales, with notes on associated behavior of other genera of captive delphinids. Contributions in Science, Los Angeles County Museum 95: 1-32.
-
McBride A, Kritzler H (1951) Observations on pregnancy, parturition, and postnatal behavior in the bottlenose dolphin. Journal of Mammalogy 32: 251-266.
-
Dudzinski KM, Sakai M, Masaki K, Kogi K, Hishii T, et al. (2003) Behavioral observations of bottlenose dolphins towards two dead conspecifics. Aquatic Mammals 29: 108-116.
-
McBride AF (1940) Meet Mister. Natural History Magazine 65: 16-29.
-
Hubbs CL, Frum WG, Neill WT, Tester JR, Welch JF, et al. (1953) Dolphin protecting dead young. Journal of Mammalogy 34: 498-518.
-
Kasuya T, Miyazaki N (1976) An observation of epimeletic behavior of Lagenorhynchus obliquidens. The Whales Research Institute 28: 141-143.
-
Smith TG, Sleno GA (1986) Do white whales, Delphinapterus leucas, carry surrogates in response to early loss of their young? Canadian Journal of Zoology 64: 1581-1582.
-
Cockcroft VG, Sauer W (1990) Observed and inferred epimeletic (nurturant) behavior in bottlenose dolphins. Aquatic Mammals 16: 31-32.
-
Harzen S, Dos Santos ME (1992) Three encounters with wild bottlenose dolphins (Tursiops truncatus) carrying dead calves. Aquatic Mammals 18: 49-55.
-
Lodi L (1992) Epimeletic behavior of free ranging rough- toothed dolphins, Steno bredanensis, from Brazil. Marine Mammal Science 8: 284-287.
-
Fertl D, Schiro A (1994) Carrying of dead calves by free- ranging Texas bottlenose dolphins (Tursiops truncatus). Aquatic Mammals 20: 53-56.
-
Palacios DM, Day D (1995) A Risso’s dolphin (Grampus griseus) carrying a dead calf. Marine Mammal Science 11: 593-594.
-
Ritter F (2007) Behavioral responses of rough-toothed dolphins to a dead newborn calf. Marine Mammal Science 23: 429-433.
-
Felix F (1994) A case of epimeletic behavior in a wild bottlenose dolphin Tursiops truncatus in the gulf of Guayaquil, Ecuador. Investigations on Cetacea 25: 227- 234.
-
Douglas-Hamilton I, Bhalla S, Wittemyer G, Vollrath F (2006) Behavioural reactions of elephants towards a dying and deceased matriarch. Applied Animal Behaviour Science 100: 87-102.
-
McComb K, Baker L, Moss C (2006) African elephants show high levels of interest in the skulls and ivory of their own species. Biology Letters 2: 26-28.
-
Appleby R, Smith B, Jones D (2013) Observations of a free-ranging adult female dingo (Canis dingo) and littermates’ responses to the death of a pup. Behavioural Processes 96: 42-46.
-
Iglesias TL, McElreath R, Patricelli GL (2012) Western scrub-jay funerals: cacophonous aggregations in response to dead conspecifics. Animal Behaviour 84: 1-9.
-
Sandler I, Ma Y, Tein J, Ayers T, Wolchik S, et al. (2010) Long-term effects of the family bereavement program on multiple indicators of grief in parentally bereaved children and adolescents. Journal of Consulting and Clinical Psychology 78: 131-143.
-
Herberman H, Fullerton C, Shear M, Ursano R (2014) Complicated grief and depression in young adults: Personality and relationship quality. Journal of Nervous and Mental Disease 202: 539-543.
-
McGrath N, Walker J, Nilsson D, Phillips C (2013) Public attitudes towards grief in animals. Animal Welfare 22: 33-47.
-
Panksepp J (1998) Affective Neuroscience: The Foundations of Human and Animal Emotions. Oxford University Press, USA.
-
Utz R, Swenson K, Caserta M, Lund D, Devries B (2014) Feeling lonely versus being alone: Bereaved person. Journals of Gerontology Series B-Psychological Sciences and Social Sciences 69: 85-94.
-
Benkel I, Wijk H, Molander U (2009) Family and friends provide most social support for the bereaved. Palliative Medicine 23: 141-149.
-
Beutel M, Willner H, Deckardt R, Von Rad M, Weiner H (1996) Similarities and differences in couples’ grief reactions following a miscarriage: Results from a longitudinal study. Journal of Psychosomatic Research 40: 245-253.
-
Fischer A, LaFrance M (2015) What drives the smile and the tear: Why women are more emotionally expressive than men. Emotion Review 7: 22-29.
-
Panksepp J (2006) Emotional endophenotypes in evolutionary psychiatry. Progress in Neuro- Psychopharmacology and Biological Psychiatry 30: 774- 784.
-
Wan M, Downey D, Strachan H, Elliott R, Williams S, et al. (2014) The neural basis of maternal bonding. PLoS One 9(3): e88436.
-
Lorberbaum JP, Newman JD, Horwitz AR, Dubno JR, Lydiard RB, et al. (2002) A potential role for thalamocingulate circuitry in human maternal behavior. Biological Psychiatry 51: 431-438.
-
Kubler-Ross E (1975) On Death and Dying (revised). Bulletin of the American College of Surgeons 60: 7-12.
-
Patton MQ (2002) Qualitative Evaluation and Research Methods. 3rd (Edn.), Sage Publications Inc., USA.
-
Boesch C, Boesch-Achermann H (2000) The Chimpanzees of the Tai Forest: Behavioural Ecology and Evolution. Oxford University Press, USA.
-
Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP (2004) Resource Selection by Animals: Statistical Design and Analysis for Field Studies. 2nd Edition, Kluwer Academic, UK.
-
Newberry RC, Swanson JC (2008) Implications of breaking mother-young social bonds. Applied Animal Behaviour Science 110: 3-23.
-
Nishida T (2008) Chimpanzee deaths at Mahale caused by a flu-like disease. Primates 49: 7-80.
-
Nurmi NO, Venkataraman VV (2011) Death among geladas (Theropithecus gelada): A broader perspective on mummified infants and primate thanatology. American Journal of Primatology 73: 405-415.
-
Herman B, Vilberg T, Bishop P, DeEskinazi FG, Panksepp J (1980) Endogenous opioids and social behavior. Neuroscience and Biobehavioral Reviews 4: 473-487.
-
Weerts EM, Miczek KA (1996) Primate vocalizations during social separation and aggression: effects of alcohol and benzodiazepines. Psychopharmacology 127: 255-262.
-
Winslow J, Hastings N, Carter CS, Harbaugh C, Insel T (1993) A role for central vasopressin in pair bonding in monogamous prairie voles. Nature 365: 545-548.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review
- Everything about Nutritional Value of the Meat Ingredients and How we can Reduce its Microbial Hazards