Electromechanical Properties of Lipid Bilayers in Peptide-Lipid Interactions: Clinical Applications
Despite the amazing diversity and complexity of living systems, all life shares the presence of a semi-permeable lipid membrane. Throughout evolution, this permeability has been finely regulated by the function of proteins that form ion channels. Similarly, some peptides can stabilize by forming aqueous pores in a dose-dependent manner. At high concentrations, however, these peptides can compromise the integrity of the membrane, destroying it. Given this behavior, a huge diversity of antimicrobial peptides and lipo-peptides has been identified. Understanding the physical-chemical principles of their interactions with lipids can contribute to the design of synthetic peptides with antimicrobial activity as an alternative to the use of conventional antibiotics.
Introduction
Every cell responds to the mechanical stimuli from its environment to a greater or lesser degree. This class of responses can be considered, therefore, as one of the most ancient, probably universal in all living organisms. Mechano- sensitive (MS) ion channels have developed the specific ability to respond, in a regulated manner, to this class of stimuli [1]. However, several other proteins also respond to this class of stimuli to a greater or lesser degree [2]. Thus, to better understand the function of any protein inserted in any lipid membrane, studying the physicochemical context surrounding them is always important. The physical properties that, as a material, the lipids in which they are immersed have, determine directly but subtly the modulation of the activities that membrane proteins exhibit [3]. It is now known that the lipid component of biological membranes determines, for example, the distribution, organization, and functioning of distinct and diverse membrane proteins, importantly MS ion channels. In general terms, the regulation of this type of membrane protein is the result of specific lipid- protein interactions, general bilayer-protein interactions, and, on the other hand, interactions at the hydrophobic- aqueous medium interface. Such interactions occur mainly due to the coupling between the hydrophobic phase of the bilayer with respect to the hydrophobic domains of these membrane proteins and, therefore, are a function of the material properties of the bilayer as a whole: its thickness, degree of compaction, intrinsic curvature, viscoelasticity, stiffness, asymmetry, surface charge and ultimately, chemical composition [4, 5, 6].
In this context, to study the material properties that lipid bilayers exhibit and their response to different physical variables such as temperature, pressure, pH, or the ionic strength of the aqueous medium in which they are formed, it is necessary to understand the interactions between the building blocks, that is, the lipids and their aqueous medium. In responding to these physico-chemical factors, many membrane proteins, notably ion channels, directly respond to the thermo-tropic and mechanical state of lipids [7, 8], particularly channels considered intrinsically mechano- sensitive [9]. Likewise, it is known that the dynamic nature of lipids enables them to form aqueous pores [10]. The formation and stability of these pores largely depend on the lipid thermo-tropic behavior [11].
MS channels in prokaryotes function mainly in cellular osmoregulation by forming large aqueous pores under events of hypo-osmotic shocks [12] and it is interesting that, compared to these, many eukaryotic MS channels fulfill similar functions, although their function is only inferred from many homologs [13, 14]. Taking as an archetypal model the structure of the bacterial channel MscL of Escherichia coli, Ghazi et al. [15] studied separately the two domains of the EcMscL protein and proved that the N-terminal domain (TM1) can form aqueous pores in liposomes, which respond in an unregulated manner to lateral membrane tension. Additionally, Clayton, et al. [16] have achieved the In vitro chemical synthesis of the EcMscL channel and the ortholog from Mycobacterium tuberculosis. Notably, in both cases, these proteins are fully functional. This suggests that synthetic peptides can mimic the activity of biological MS channels.
Lytic Peptides
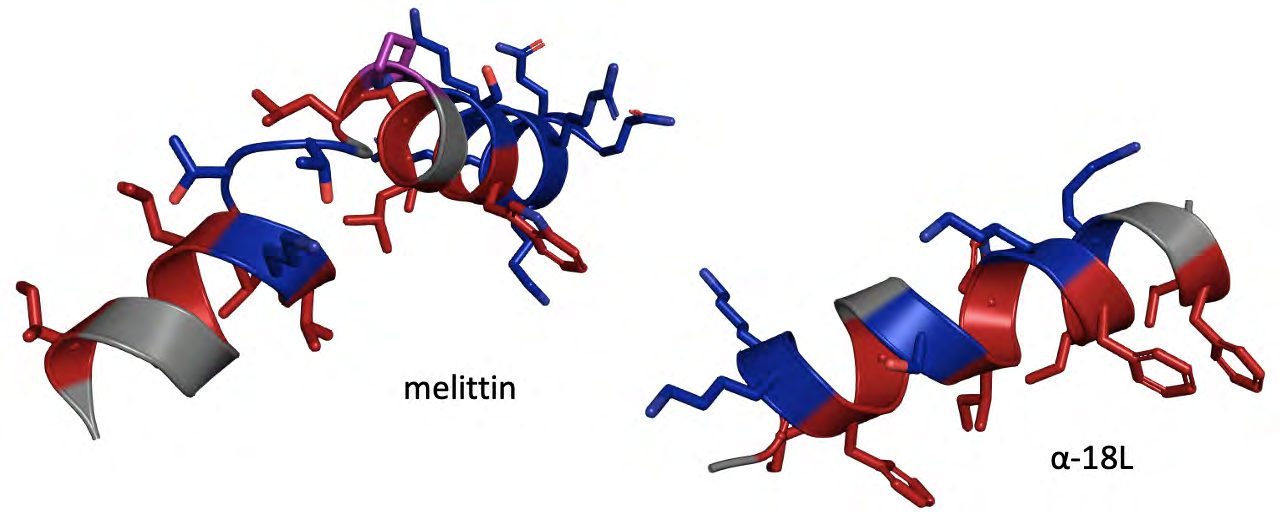
Nature always surprises us. A wide variety of pore- forming peptides (PFPs) such as alamethicin, melittin, magainin, and lipo-peptides such as dapto-mycin or feng- mycin can insert themselves into membranes, aggregating into aqueous pore-forming complexes. It is worth mentioning that the study of these peptides and other similar ones is also an interesting antimicrobial alternative [17, 18, 19]. Remarkably, using giant liposomes, it has been proven that the presence of antimicrobial peptides (AMPs) such as melittin from Apis mellifera venom (GIGAVLKVLTTGLPALISWIKRKRQQ) (Figure 1), induces the formation of aqueous pores and even induce vesicular lysis, depending on the peptide-to-lipid ratio (P/L) [20]. Therefore, at appropriate concentrations, melittin can form stable pores. Under such conditions, these model systems are able of dissipating osmotic gradients, responding to increases in membrane tension, a mechanism analogous to that exhibited by bacterial MS channels [1, 2, 9]. Interestingly, the activity of this class of natural peptides can also be directly compared to that shown by some synthetic amphipathic peptides, such as peptides of the α-18L class (GIKKFLGSIWKFIKAFVG) (Figure) that exhibit similar osmo- protective behavior [21]. Once inserted into the membrane, these peptides perturb the bilayer and create peptide nucleation sites from which aqueous pores are formed responding to micro-environmental osmotic changes and releasing several osmolytes. Therefore, at least potentially, it is possible to synthesize peptides that can directly simulate the activity of forming aqueous pores with a possible osmo- regulatory function or induce lysis at high concentrations.
À la carte Peptides
The perspective to design peptides with antimicrobial activity is not new [22, 23]. However, albeit research efforts have focused mainly on characterizing peptides obtained directly from their biological sources or synthetic ones, only a few of them have been approved by the FDA [24]. It becomes evident that it is necessary to continue characterizing these potential antimicrobial agents at the biophysical level to propose using AMPs and similar compounds such as lipo-peptides (LPs) and as alternative antibiotics. Indeed, Cubicin® (daptomycin), an LP that induces membrane permeability in Gram(+) bacteria, including methicillin- resistant Staphylococcus aurous and vanco-mycin resistant Enterococci bacteria is the first LP approved as an intravenous drug by the FDA for the treatment of serious skin (approved in 2003) and bloodstream infections (approved in 2006) [25]. Cubist Pharmaceuticals has devoted a great deal of effort to obtaining compounds derived from dapto-mycin by chemical or biosynthetic methods. The cyclic lipo-peptide CB-183,315 has been characterized In vitro and In vivo against Clostridium difficile, showing good quality standards and activity in hamsters; it is now in phase 3 clinical trials for the treatment of C. Difficile-associated diarrhea and it is pending to be finally approved by the FDA [26, 27].
Conclusion
The emergence of pathogens resistant to conventional antibiotics is increasing rapidly and globally. This problem has raised the alternative of using other antimicrobial control strategies. Antimicrobial peptides and lipopeptides are promising alternatives that target biological membranes by diverse mechanisms of action, including membrane permeabilization and detergent effect. Studying the factors that determine these interactions at the molecular level will allow obtaining the necessary knowledge for the intelligent design of synthetic peptides with potential activity against several multidrug-resistant strains. Maintaining research efforts in the study of peptide-lipid interactions will surely contribute to the development of a new generation of antimicrobial therapies and increasingly effective control strategies in the face of the growing problem of bacterial multidrug resistance.
References
-
Kung C (2005) A Possible Unifying Principle for Mechanosensation. Nature 436: 647-654.
-
Teng J, Loukin S, Anishkin A, Kung C (2015) The Force- From-Lipid (FFL) Principle of Mechanosensitivity, at Large and in Elements. Pflugers Arch 467(1): 27-37.
-
Anishkin A, Loukin SH, Teng J, Kung C (2014) Feeling the Hidden Mechanical Forces in Lipid Bilayer is an Original Sense. Proc Natl Acad Sci USA 111(22): 7898-7905.
-
McIntosh TJ, Simon SA (2006) Roles of Bilayer Material Properties in Function and Distribution of Membrane Proteins. Annu Rev Biophys Biomol Struct 35: 177-198.
-
Andersen OS, Koeppe RE (2007) Bilayer Thickness and Membrane Protein Function: An Energetic Perspective. Annu Rev Biophys Biomol Struct 36: 107-130.
-
Phillips R, Ursell T, Wiggins P, Sens P (2009) Emerging Roles for Lipids in Shaping Membrane-Protein Function. Nature 459: 379-385.
-
Seeger HM, Aldrovandi L, Alessandrini A, Facci P (2010) Changes in Single K(+) Channel Behavior by a Lipid Phase Transition. Biophys J 99(11): 3675-3683.
-
Schmidt D, del Mármol J, MacKinnon R (2012) Mechanistic Basis for Low Threshold Mechanosensitivity in Voltage-Dependent K+ Channels. Proc Natl Acad Sci USA 109(26): 10352-10357.
-
Perozo E, Kloda A, Cortes DM, Martinac B (2002) Physical Principles Underlying the Transduction of Bilayer Deformation Forces During Mechanosensitive Channel Gating. Nat Struct Biol 9(9): 696-703.
-
Blicher A, Heimburg T (2013) Voltage-Gated Lipid Ion Channels. PLoS ONE 8(6): e65707.
-
Mosgaard LD, Heimburg T (2013) Lipid Ion Channels and the Role of Proteins. Acc Chem Res 46(12): 2966- 2976.
-
Martinac B (2007) 3.5 Billion Years of Mechanosensory Transduction: Structure and Function of Mechanosensitive Channels in Prokaryotes. Current Topics in Membranes 58: 25-57.
-
Kloda A, Martinac B (2001) Mechanosensitive Channels in Archaea. Cell Biochem Biophys 34(3): 349-381.
-
Balleza D, Gómez-Lagunas F (2009) Conserved Motifs in Mechanosensitive Channels Mscl and Mscs. Eur Biophys J 38(7): 1013-1027.
-
Park KH, Berrier C, Martinac B, Ghazi A (2004) Purification and Functional Reconstitution of N- and C-Halves of the Mscl Channel. Biophys J 86(4): 2129- 2136.
-
Clayton D, Shapovalov G, Maurer JA, Dougherty DA, Lester HA, et al. (2004) Total Chemical Synthesis and Electrophysiological Characterization of Mechanosensitive Channels from Escherichia Coli and Mycobacterium Tuberculosis. Proc Natl Acad Sci USA 101(14): 4764-4769.
-
Browne K, Chakraborty S, Chen R, Willcox MD, Black DS, et al. (2020) A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. Int J Mol Sci 21(19): 7047.
-
Kabelka I, Vácha R (2021) Advances in Molecular Understanding of A-Helical Membrane Active Peptides. Acc Chem Res 54(9): 2196-2204.
-
Zhang QY, Yan ZB, Meng YM, Hong XY, Shao G, et al. (2021) Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Military Med Res 8(48): 1-25.
-
Lee MT, Sun TL, Hung WC, Huang HW (2013) Process of Inducing Pores in Membranes by Melittin. Proc Natl Acad Sci USA 110(35): 14243-14248.
-
Polozov IV, Anantharamaiah GM, Segrest JP, Epand RM (2001) Osmotically Induced Membrane Tension Modulates Membrane Permeabilization By Class L Amphipathic Helical Peptides: Nucleation Model of Defect Formation. Biophys J 81(2): 949-959.
-
Som A, Vemparala S, Ivanov I, Tew GN (2008) Synthetic Mimics of Antimicrobial Peptides. Biopolymers 90(2): 83-93.
-
Kundu R (2020) Cationic Amphiphilic Peptide: A Nature‐Inspired Synthetic Antimicrobial Peptide. ChemMedChem 15(20):1887-1896.
-
Ryu M, Park J, Yeom JH, Joo M, Lee K (2021) Rediscovery of Antimicrobial Peptides as Therapeutic Agents. J Microbiol 59(2): 113-123.
-
Enoch DA, Bygott JM, Daly ML, Karas JA (2007) Daptomycin. Journal of Infection 55(3): 205-213.
-
Mascio CT, Mortin LI, Howland KT, Van Praagh AD, Zhang S, et al. (2012) In vitro and In vivo Characterization of CB- 183,315, a Novel Lipopeptide Antibiotic for Treatment of Clostridium Difficile. Antimicrob Agents Chemother 56(10): 5023-5030.
- Gallic and Citric Acid Present in the Peels of Tropical Fruits as an Alternative in the Fight against Cancer
- Treating the Forehead Lines with Combination of Forehead and Glabellar Botulinum Toxin Among Japanese Patients
- Clinical Evaluation of Patients Suffering from Breast Cancer & Determination of Treatment Therapies and Better Strategies Related to Breast Cancer
- Medieval Recipes by Al-Zahrāwī for Heart Palpitations Treatment
- Etiology and Prescription Errors of Myocardial Infarction in Different Health Care Systems of Azad Kashmir
- Early Diagnosis and Multidisciplinary Management of Turner Syndrome: A Paediatric Case Study