Evaluation of Prebiotic Utilization and Probiotic Properties of Potential Probiotic Bacteria of Food and Human Origin
Probiotics are live organisms that, when administered adequately, confer health benefits to the host, including cancer prevention, blood cholesterol reduction, and immune system strengthening. They play an essential role in protecting the mammalian intestines against the colonization of pathogenic microbes. It increases integrity by improving the stimulation of epithelial cells. Lactic acid bacteria are generally found in foods and medicine and possess probiotic qualities. This study aimed to evaluate four probiotic isolates, BM1, H39, H31, and IF1, potential probiotic candidates isolated from humans and food origin has grown on MRS (De Man, Rogosa, and Sharpe) medium. The isolates were sequenced using 16SrRNA sequencing and were identified as Lactobacillus fermentum (BM1), Lactobacillus crispatus (H39), Enterococcus faecium (H31), and Lactobacillus helveticus (IF1). The isolate was screened and characterized based on growth characteristics, Gram’s reaction, and functional properties like auto-aggregation, salt aggregation test (SAT), and viability during oro-gastro-intestinal transit was performed. Hemolytic activity, prebiotic score, and antibiotic susceptibility were determined.
Introduction
Lactic acid bacteria (LAB) are an essential group of bacteria that comprises fermentative bacteria used in the production of fermented foods and drinks and present in vegetables, fruits, the gastrointestinal tract, and the vagina of humans and animals [1]. LAB are Gram-positive, non- sporulating, non-respiring cocci or rods that use glucose to produce lactic acid as a primary catabolic end product [2]. Certain LAB strains are characterized by the ability to convert lactose to improve the digestibility of fermented dairy products as well as preservation [3, 4]. They are generally regarded as safe because of their wide use in the food and dairy industry and their beneficial effects of probiotic properties on human health. Probiotic strains of LAB belong to the genera Lactobacillus, Lactococcus, and Bifidobacterium.
As per the WHO/FAO (2002) guidelines, “Probiotics are live microorganisms which, when administered adequately, confer health benefits to the host.” The most widely used tests for LAB to act as potential probiotics are resistance and survival in gastric acidity and bile salts. Due to large amounts of bile and gastric juice secretion, it is necessary to protect probiotic strains that strive for low pH, enzymes, and bile salts. The Prebiotic was first defined as a “Non-digestible food component that benefits the host by significantly stimulating the growth and/or activity of one or a limited number of bacteria in the gut, and thus improves host health” [5]. Dietary fibers, mostly oligosaccharides and polysaccharides, have not been degraded by the enzymes of the human digestive system, so to overcome the fiber deficiency of the diet, prebiotics is added to the food. Prebiotics are complex sugars that provide fuel for the bacteria in GIT. The ingested prebiotic stimulates the whole indigenous population of bifidobacteria and lactobacilli to grow. Inulin, fructose- oligosaccharides (FOSs), galactooligosaccharides (GOSs), lactulose, and human milk oligosaccharides (HMOs) are the most widely studied and approved prebiotics [6]. Unlike probiotics, most prebiotics is food ingredients in biscuits, cereals, chocolate, spreads, and dairy products.
One of the most valuable attributes of probiotics is providing gut microbial equilibrium. Lactobacillus strains have been characterized for their ability to adhere to and colonize the mucus layer of the intestines and produce antimicrobial substances [7]. Well-known examples of LAB widely accepted and used as probiotics are L. crispatus, L. paracasei, L. plantarum, L. brevis, L. fermentum, L. durianis, L. rhamnosus, B. longum, B. animalis, B. bifidum, Lactococcus lactis, Weissella paramesenteroides, and Enterococcus faecium. The genus Enterococcus has been very controversial due to its virulence for the last 20 years, and the two main reasons are its pathogenicity and multidrug resistance [8]. Enterococcus has also demonstrated probiotic properties by improving intestinal flora balance.
E. faecium is used in farm animals as a probiotic supplement and positively affects weight gain and diarrhea in pigs [9]. Some species like E. faecalis, E. durans, E. hirae, and E. faecium have been typically associated with human GIT and hence have a fecal origin [10]. For E. faecium to be used as a pharmaceutical probiotic, it requires the preparation of a registration file and approval from the health authority of the respective country [11].
Many studies have evaluated the effect of the Enterococcus genus probiotic strains (mainly E. faecium). Generally, human-origin studies are scarcer when compared to animal and food-origin applications. So, in this study, we aimed to isolate, characterize, and identify potential strains of probiotics isolated from human feces, breast milk, and fermented food to assess the functional properties and safety aspects of using them as a pharmaceutical probiotics.
Materials and Methods
Chemicals and Media
De Man Rogosa Sharpe (MRS) broth and MRS basal broth (MRS-BB). MRS-BB contained all the components of the MRS medium except beef extract and glucose compared to the regular MRS medium. All the chemicals used in this study have been procured from HiMedia, Mumbai, India.
Sample Collection
Four samples were collected of human and food origin (infant feces, human feces, breast milk, and idli batter). Lactobacillus Probiotic strains were isolated using MRS broth and preserved in 10% skim milk at 4°C. Cultures (24 h) were subcultured into MRS plates and incubated for 48 h. The colonies were harvested by a wire loop, resuspended in MRS broth containing glycerol 40% (v/v), and stored at -20°C.
Isolation and Identification of LAB
The samples were aseptically enriched in MRS-BB for 24- 48 h in a test tube containing 10 ml MRS medium spiked with 1% glucose, sucrose, mannitol, lactose, trehalose, or FOS. From each sample, a 1:10 dilution was subsequently made using sterile PBS, followed by a 10-fold serial dilution. 0.1 ml from the appropriate dilutions were plated on MRS agar and incubated for 24-48 h at 37°C. Spindle-shaped colonies were inoculated in MRS broth and incubated at 37°C for 48 h. As judged by microscopic observations for homogeneity of cellular morphology, pure strains were maintained in 10% skim milk at 4°C. Isolates were further tested for Gram reaction, catalase test, and cell morphology according to the methods described [12]. Twenty Gram-positive and catalase- negative isolates were randomly selected as presumptive LAB, very short to very long rods occurring singly or in pairs or chains.
Molecular identification of isolates having potential probiotic properties was carried out. These isolates were identified based on the 16S rRNA gene sequence. The colony morphology of the isolates incubated on MRS medium plates at 37°C for 48 h was evaluated. Gram staining was performed. Cell morphology was determined by SEM (Junagadh Agricultural University, Gujarat, India).
Scanning Electron Microscopy
Cells from 18 h cultures of two selected LAB isolates were harvested by centrifugation (5000xg, 10 min, 5°C), washed twice with sterile PBS (pH 7.4), and fixed overnight at 4°C with 2.5% glutaraldehyde in 0.01 M PBS. After washing with PBS, the cells were post-fixed with 2% osmium tetroxide solution for 2 h. Cells were washed with PBS and dehydrated through graded ethanol. The samples were gold-covered (1-2 nm) by cathodic spraying and examined at 15 kV using Carl Zeiss EVO 18 Scanning electron microscope.
Screening of LAB
Determination of pH, NaCl, and phenol tolerance: In- vitro assessment of probiotic strains involves resistance to low pH, bile salt, NaCl, and phenol. Bile salt, pH, and salt tolerance (sodium chloride) were evaluated as described by Ambalam, et al. [13] with some modifications. MRS broth varying in pH (2.5-4), bile salt (0.5-4%), and NaCl (2-6%) were inoculated with 0.1 ml of 24 h old culture of Probiotic bacterial strain and incubated for 24 h at 37°C. The growth of microorganisms was visually evaluated. All the tubes, including the control, were incubated (37°C, 24 h). Take a sterile microtiter plate (MTP) and add 250μl MRS broth. Take selected LAB isolates, add 20μl into the MTP, and incubate for 24 h at 37oC. After incubation, take 180μl Phosphate Buffer Saline, add into another MTP, and add a 20μl sample from the incubated MTP. Regular MRS broth is added as a control, and bacterial growth is determined by measuring OD600nm using a microplate photo reader (BioTek instruments).
Prebiotic scor: A prebiotic score (PS) is the highest growth achieved by a strain on a prebiotic NDO relative to their growth on glucose that was considered as 100% was determined using the formula: PS = (A/ B) x 100%, where A and B is the mean OD600nm value of a strain grown with each of the oligosaccharide substrate or glucose after 24 h of growth [14].
Functional properties
Auto-Aggregation: Activated cultures were inoculated into MRS broth containing 0.05% and 0.15% (w/v) bile and incubated for 24 h at 37°C. Activated cultures were harvested by centrifugation at 5000×g for 10 min, washed twice in PBS, and re-suspended in PBS to give a final OD660nm as a 0 h. After 4 h of incubation at room temperature, the OD of suspensions was measured at OD 660 nm. The percent of auto-aggregation was expressed as follows: % Auto - aggregation = [(OD1 – OD2)/OD1] × 100 Where; OD1 is the first optical density (0 h), and OD2 is the optical density after 4h. Antimicrobial activity: The antimicrobial activity of LAB isolates was determined using McVay and Rolfe (2000) with some modifications [15]. The cell-free supernatant (CFS) obtained by harvesting 24 h old cultures from MRS- BB containing 1% glucose by centrifugation (Eppendorf) at 5000×g for 10 min was sterilized through a 0.22 μm cellulose nitrate membrane filter. Pathogens (Bacillus cereus, Bacillus subtilis, Enterococcus faecalis, Escherichia coli, Streptococcus mutans, Staphylococcus aureus, Pseudomonas aeruginosa, and Shigella flexneri) grown in N-broth for 24 h at 37ºC. Wells of 7 mm were made using a cup-borer, and 100 μL CFS of the isolate was added. The plates were then refrigerated for 2-3 hours at 4°C and later were transferred for incubation at 37°C. After 24h, the surrounding well measured the zone of inhibition.
Salt Aggregation Test (SAT): The cell surface hydrophobicity of isolates was also determined by salt aggregation test (SAT) as previously described by Lindahl M, et al. [16]; 10μl aliquot of fresh cell suspension in PBS was mixed on a glass slide with 10 μl of ammonium sulfate (pH 6.8) of various molarities (0.02-4 M) [16]. The formation of cell aggregates was observed after 1 min by visual reading. The lowest concentration of ammonium sulfate, giving visible aggregation, was scored as the SAT hydrophobicity value.
Viability during oro-gastro-intestinal transit: The viability of the cells in the presence of lysozyme was determined, as described by Pinto, et al. [17]. To simulate the in vivo dilution by saliva, 100μl of bacterial suspension was mixed in a sterile electrolyte solution consisting of g/l of CaCl2 (0.22), NaCl (6.2), KCl (2.2), NaHCO3 (1.2) in the presence of 0.01% of lysozyme (Himedia, Mumbai). Bacterial suspension in electrolyte solution without lysozyme was included as a control. Samples were incubated at 37°C for 10 min. After that, 0.1ml sample was harvested serially diluted up to 107 dilutions; the last three dilutions were plated on MRS agar and incubated for 48 h at 37°C to determine the viable cell counts, expressed as log CFU/ml. Viability during simulated gastric (SGF) and intestinal fluid (SIF) transit was evaluated as described by Charteris, et al. [18]. A 100μl cell suspension was mixed with 2 ml of simulated gastric fluid (SGF) and incubated for 2h at 37°C. The cells pelleted by centrifugation were resuspended in 1ml of simulated intestinal fluid (SIF) and incubated at 37°C for 3h. SGF and SGF–SIF-treated cells were serially diluted and plated on MRS agar to determine cell viability. The control consisted of cells treated with a phosphate buffer instead of SGF and SIF. The SGF comprised 0.01% pepsin, and pH 2 was adjusted with 1M HCl. SIF comprised 0.03% pancreatic, 0.5% bile salt, and pH 6.8 adjusted with 1M NaOH.
Prebiotic utilization: Washed cells of probiotic strains were inoculated in 5 ml of modified MRS medium, supplemented with 1% (w/v) of each prebiotic (FOS and Inulin) as the sole carbon source. The cell density was adjusted to an OD of 0.5 (A600nm), and cells were incubated at 37°C for 72 h. The growth of bacterial probiotic strain was determined by measuring the OD600 and pH of the culture broth at various time points.
Safety Aspects
Haemolytic activity: The isolates were streaked on MRS agar supplemented with 5% human blood and incubated at 37°C for 48 h and observed for complete hemolysis with transparency (β hemolysis), partial hemolysis without transparency (α hemolysis), and no hemolysis (γ hemolysis), hemolytic activity [19]. Bacillus megaterium served as a positive control.
Biogenic amine formation: Activated cultures (24h) are streaked on tyrosine-containing agar medium and incubated for 24 h at 37ºC. A zone of clearance surrounding the colony was observed. Enterococcus faecalis served as a positive control for biogenic amine formation.
Results
Isolation of LAB
Fourteen probiotic isolates were isolated that formed round, creamy white colonies on the MRS agar plate. Isolates were (Table 1) Gram-positive, catalase-negative rods, and coccid. These LAB were isolated from different sources such as human and infant fecal, food origin like idli batter, and human breast milk samples. The preliminary characterization and microscopic investigation showed that all isolates were Gram-positive, catalase-negative, and curdling.
| No. | Isolates | Gram’s reaction | Curdling | Catalase |
|---|---|---|---|---|
| 1 | IST1 | Gram-positive long rods | + | + |
| 2 | IF1 | Gram-positive long rods | + | - |
| 3 | IL1 | Gram-positive long rods | + | - |
| 4 | IL3 | Gram-positive short rods | - | - |
| 5 | BM1 | Gram-positive short rods | + | - |
| 6 | BM12 | Gram-positive short rods | + | - |
| 7 | H31 | Gram-positive cocci | + | - |
| 8 | H32 | Gram-positive long rods | + | - |
| 9 | H33 | Gram-positive long rods | + | - |
| 10 | H34 | Gram-positive short rods | - | - |
| 11 | H35 | Gram-positive short rods | + | - |
| 12 | H37 | Gram-positive long rods | + | - |
| 13 | H38 | Gram-positive long rods | + | - |
| 15 | H39 | Gram-positive long rods | + | - |
| 16 | H54 | Gram-positive long rods | + | - |
| 17 | H56 | Gram-positive short rods | - | - |
Table 1: Isolation, characterization, and identification of LAB from human and food-origin samples. *(+) positive result (-) nega
Identification and Scanning Electron Microscopy
Molecular identification of the isolates was made based on 16S rRNA sequence analysis. Isolate H31 was identified as Enterococcus faecium, IF1 was identified as L.helveticus, BM1 as L.fermentum, and H39 as L. crispatus (Figures 1a-1d).
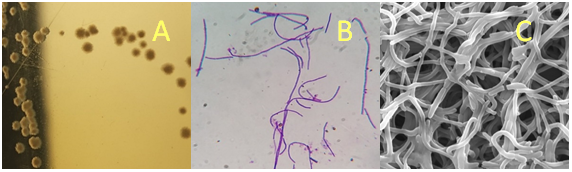
Figure 1(a): Colony characteristics and morphology of L. helveticus IF1 (A) Colony characteristics inoculated in MRS medium; (B) Gram staining; (C) Scanning Electron Microscopy (SEM).
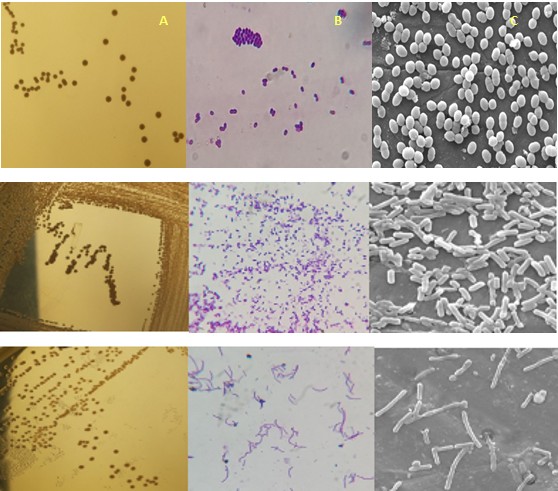
Figure 1(b). Colony characteristics and morphology of E. faecium H31 (A) Colony characteristics inoculated in MRS medium; (B) Gram staining; (C) Scanning Electron Microscopy (SEM).
(A) (B) (C) Figure 1(c). Colony characteristics and morphology of L.fermentum BM1 (A) Colony characteristics inoculated in MRS medium; (B) Gram staining;(C) Scanning Electron Microscopy (SEM).
(A) (B) (C) Figure 1(d). Colony characteristics and morphology of L.crispatus H39 (A) Colony characteristics inoculated in MRS medium; (B) Gram staining;(C) Scanning Electron Microscopy (SEM).
Screening of LAB
Growth in the presence of low pH, bile, NaCl, and phenol: Based on Table 2, all the selected isolates showed tolerance and growth conditions prevailing in the gastrointestinal tract, including 1% bile salts, 2.5 pH,0.4% phenol, and 4%NaCl. All isolates grew in the presence of 1% bile, 2.5 pH, 0.4% phenol, and 4% NaCl while the isolates tolerated conditions except H35, H54, and H56.
| Isolates | Bile | pH | Phenol | NaCl | ||||
|---|---|---|---|---|---|---|---|---|
| 0.50% | 1% | 3 | 2.5 | 0.20% | 0.40% | 2% | 4% | |
| IST1 | T | T | T | T | T | T | T | T |
| IF1 | G | G | G | G | T | T | G | G |
| IL1 | G | G | G | G | G | G | G | G |
| IL3 | T | T | T | T | T | T | T | T |
| BM1 | G | G | G | G | G | G | G | G |
| H28 | G | G | G | G | G | T | T | T |
| H30 | G | G | G | G | G | G | G | G |
| H31 | G | G | G | G | G | G | G | G |
| H32 | G | G | G | G | T | T | T | T |
| H37 | G | G | T | T | G | T | T | T |
| H38 | G | G | G | G | G | G | G | G |
| H39 | G | G | G | G | G | G | G | G |
| H56 | T | T | T | T | T | T | T | T |
Table 2: The ability of LAB isolates to grow and tolerate conditions of low pH, bile, phenol, and NaCl. *G - Growth T – Tolerance
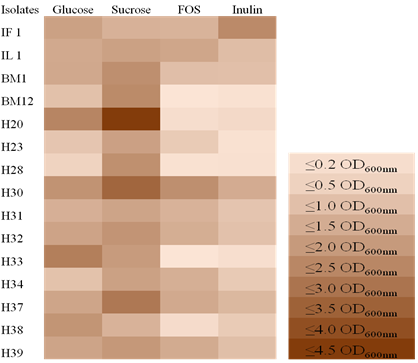
Prebiotic score: The growth of LAB isolates in modified MRS broth supplemented with different carbohydrate sources was evaluated in terms of OD600nm at 24 h (Figure 2). The growth of LAB strains on glucose was used as a control. All isolates utilized sucrose and glucose effectively, while H30 and IF1 gave the highest prebiotic utilization in FOS and Inulin compared to other isolates. Here, isolates BM12, H20, H28, and H33 gave the lowest prebiotic utilization compared to the other isolates.
Functional Properties
Auto-aggregation
Bacterial aggregation between microorganisms of the same strain (auto-aggregation) is vital in several ecological niches, especially in the human gut. Both Lactobacillus strains showed high auto-aggregation abilities (Table 3). In the present study, the auto-aggregation of probiotic strains (BM1, H39, and GG) ranged from 13-50%, whereas E. faecium H31 was 21%.
| Isolates | % Auto aggregation | Salt aggregation test | ||
|---|---|---|---|---|
| MRS | MRS + 0.05% Bile | MRS+ 0.15% Bile | ||
| E. faecium H31 | 21±6 | ≥ 4.00 | ≥ 4.00 | ≥ 0.02 |
| L. fermentum BM1 | 17±5 | ≥ 3.20 | ≥ 3.20 | ≥ 2.00 |
| L. crispatus H39 | 13±7 | ≥ 1.20 | ≥ 1.20 | ≥ 0.02 |
| L. helveticus IF1 | 50±2 | ≥ 0.02 | ≥ 0.02 | ≥ 0.02 |
| L. rhamnosus GG | 26±8 | ≥ 3.20 | ≥ 3.20 | ≥ 0.02 |
Table 3: % auto-aggregation after 4 hours and SAT of LAB.
Salt Aggregation Test (SAT)
Bacterial strain H31 showed high hydrophobicity by SAT assay with a minimum ammonium sulfate concentration of <1.2M, and L. rhamnosus GG was used as a control (Table 3).
Viability during oro-gastro-intestinal transit (SGF-SIF)
The selected LAB isolates were treated with sterile electrolyte solution in the presence of lysozyme, pepsin, and pancreatin to check the tolerance towards the SGF-SIF (Figure 2). When LAB strains were incubated in the presence of lysozyme for 10 min; all strains showed 95% viability. The gastric phase (pepsin) E. faecium H31 showed the highest viability of 87%, and the intestinal phase (pancreatin with bile salt). Isolates H31 and GG retained up to 60% viability in simulated oro-gastro-intestinal transit (OGIT) (Figures 3a, 3b).
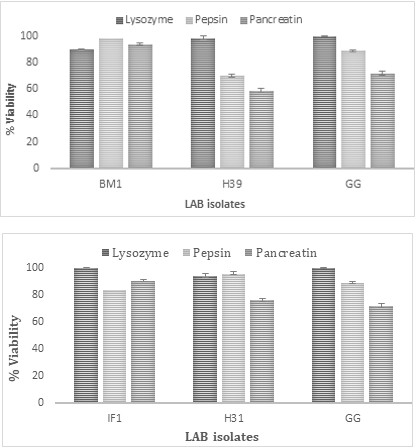
Figure 3(a): Showing the % viability of BM1 and H39 during OGIT with GG as a positive control.
Figure 3(b): Showing the % viability of IF1 and H31 during OGIT with GG as a positive control.
Antimicrobial Properties
CFSs of seven isolates obtained by growing in the presence of either 1% glucose or FOS or Inulin were evaluated for antimicrobial activity (AMA) against E. coli, S. typhi, E. fecalis, and Shigella spp at 1:1 dilution (Figure 6).
Against Shigella spp AMA was higher in the presence of FOS, followed by Inulin. With FOS, AMA was significantly higher than the rest of the carbon source. On the contrary, against S. typhi, AMA was higher in the presence of glucose, followed by sucrose, FOS, and Inulin.
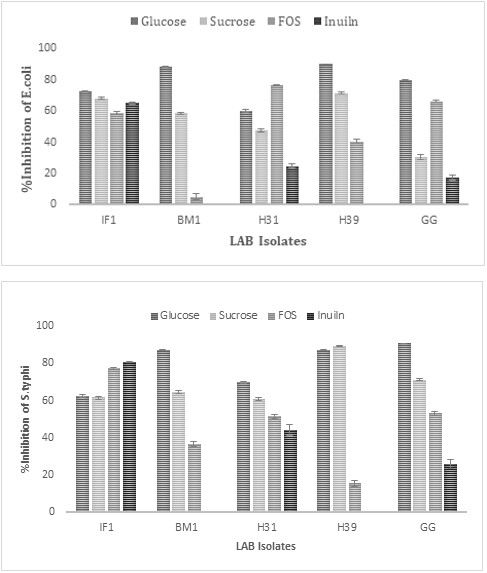
Figure 4(a): Antimicrobial activity of LAB against E.coli.
Figure 4(b): Antimicrobial activity of LAB against S.typhi.
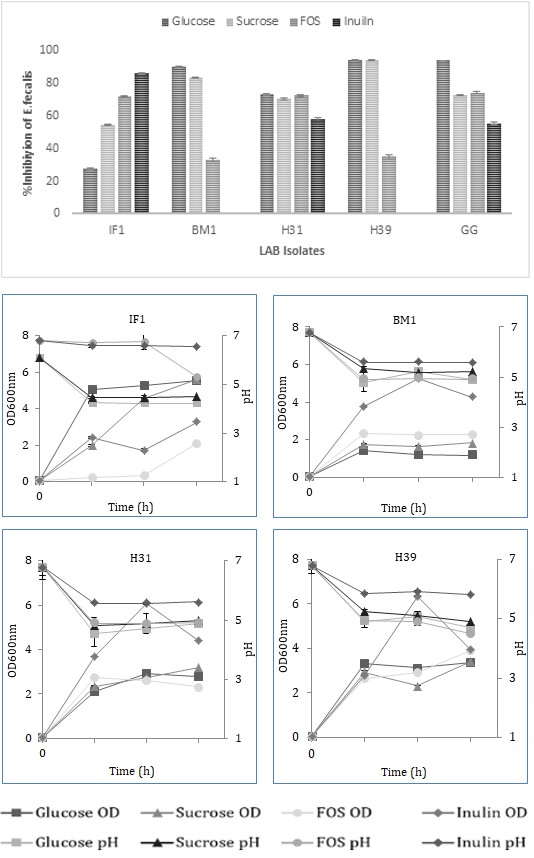
Figure 4(c): Antimicrobial activity of LAB against E. faecalis. Prebiotic utilization Isolates showed varying degrees of growth, pH drop, and prebiotic utilization when grown in the presence of different prebiotics, FOS, or Inulin measured after 24, 48, and 72 h of growth. Strains showed maximum prebiotic utilization and pH drop at 48 h of growth. Strains H39, H31, H34, and H32 showed high prebiotic utilization with a pH drop up to pH 4.9 of the medium in the presence of FOS or Inulin. H39 showed the highest prebiotic utilization in the presence of Inulin, followed by FOS. H31, H34, and H32 showed the lowest prebiotic utilization in the presence of Inulin or FOS. The prebiotic utilization of the three strains (IF1, IL1, and BM1) was 50% in the presence of Inulin with the pH drop of CFS up to pH 5.7 (Figure 5).
Safety aspects of LAB strains
Haemolytic test
None of the probiotic strains produced a clearance zone on the plate containing 5% human blood.
Biogenic amine formation
None of the probiotic strains produced a zone of clearance on the plate containing 1% tyrosine, suggesting that the tyrosine decarboxylase activity was non-detectable. Enterococcus faecalis, included as a positive control, produced a clear zone around colonies on the tyrosine plate, providing evidence of the production of tyrosine decarboxylase activity.
Discussion
This study aimed to isolate, screen, and characterize LAB isolated from human feces, infant feces, human breast milk, and idle batter. The isolate was screened for tolerance to low pH, bile, NaCl, and phenol, biochemical tests, functional properties like auto-aggregation, SAT, and antimicrobial activity against Gram-positive and negative pathogens. The evolution of prebiotic non-digestible oligosaccharides preference of probiotic LAB has also been performed. Tests were executed to check auto-aggregation, biogenic amine production, and hemolytic activity.
Twenty isolates were Gram-positive rods, non-motile, non-sporulating, and catalase-negative were considered Lactic Acid Bacteria (LAB). LAB isolates in the human GIT are non-pathogenic and form a vital biodefense factor. Therefore, it is worth exploring the healthy native microflora of GIT as probiotics. Screening and characterizing these LAB strains is essential to obtain ideal and novel probiotics, as some LAB does not confer health benefits to the host. The potential probiotic lactobacilli should be of healthy human origin, non- pathogenic, and non-hemolytic though the lactobacilli have a long history of being safe for human-conferred GRAS status.
Molecular identification of potent probiotic strains was made by 16S rRNA sequence analysis. Potential probiotic strains were obtained from different habitats, such as idli batter and human fecal belonging to L. helveticus IF1 and E. faecium H31, respectively. The strain BM1 was identified as L. crispatus, and H39 was identified as L. fermentum isolated from human breast milk and infant feces.
The most important step towards selecting a probiotic candidate is investigating the strain under conditions mimicking the GIT. For bacterial cells, the stress begins in the mouth, with lysozyme-containing saliva, and continues in the stomach and upper intestine. Probiotic bacteria are first exposed to saliva containing lysozyme and electrolytes in the oral cavity. LAB strains retained viability in the presence of lysozyme provides evidence of successful transit through the oral cavity. Before reaching the intestinal tract, probiotic bacteria must survive during transit through the stomach, where gastric acid secretion constitutes a primary defense mechanism against the ingested microbes. The gastric fluid is a crucial barrier to overcome before reaching the site of action [20]. This enables their viable passage through the GIT to flourish in the existing nutritional and ecological conditions. Isolates were evaluated under in-vitro conditions for their acid, bile, NaCl, and phenol tolerance properties.IF1, BM1, H31, and H39 isolates could grow at pH 2.5, whereas other isolates tolerated up to pH 3. Such survival would be promising as probiotic bacteria are not directly exposed to such low pH [21]. Acid tolerance of the isolates is essential to withstand the gastrointestinal environment and for their use as dietary adjuncts in acidic food preparations [22].
After overcoming the harshness of gastric juices, they are now exposed to bile salts. The gallbladder secretes bile salts into the duodenum, which is helpful in fat digestion. In the GIT, the bile concentration fluctuates from 1.5 to 2% (w/v) in the first hour of digestion and decreases to 0.3% [23]. Table 3 shows that isolates IF1, BM1, H31, and H39 can grow with 1% bile salt, while IST1, IL3, H35, H54, and H56 tolerate 1% bile salt. Isolates IF1, BM1, H31, and H39 can grow with 4% NaCl. Such potential would be necessary for food processing and preservation and confer a competitive advantage over undesirable organisms. The strains also tolerated 0.4% phenol, a toxic metabolite produced upon deamination of some aromatic amino acids during putrefaction by intestinal bacteria [24].
Another essential functional characteristic feature of a potential probiotic strain is to exert AMA through which they prevent various infections while helping in the homeostasis of gut microbiota, which is principally attributed to extracellular antibacterial metabolites, such as organic acids, antimicrobial peptides, and hydrogen peroxide [25]. CFSs of seven isolates obtained by growing either 1% glucose, FOS, or Inulin was evaluated for AMA against E.coli, S.typhi, E. faecalis, and Shigellaspp at 1:1 dilution, Against Shigella spp AMA, was higher in the presence of FOS, followed by Inulin. With FOS, AMA was significantly higher than the rest of the carbon source. On the contrary, against S.typhi, AMA was higher in the presence of glucose, followed by sucrose, FOS, and Inulin.
Studies on the auto-aggregation mechanism in probiotics showed that proteins present in the culture supernatant and proteins or lipoproteins and polysaccharides located on the cell surface area are involved in cell aggregation [26]. LAB strains showed lower SAT values (<1.2 M) and auto-aggregation up to 50% after 4 h, implicating the hydrophobic nature of their cell surface, which may facilitate the colonization of these strains in the gut. Many researchers have reported that growth in bile alters auto aggregation [27, 28]. In the present study, the auto-aggregation of some strains significantly decreased after bile treatment.
Biogenic amines are molecules with allergenic properties. They are found in fermented products and are synthesized by LAB through the decarboxylation of amino acids in the food matrix. None of the probiotic strains produced detectable amine from tyrosine. Biogenic amines are related to health hazards, including food poisoning. Histamine and tyramine, in particular, cause hypertension and headaches [29]. L. helveticus IF1, E. Faecium H31, L. crispatus BM1, and L. fermentum H39 strains are safe; they do not produce biogenic amine or hemolytic activity.
Prebiotics increase beneficial microflora and reduce the build-up of disease-causing pathogens. Changing cytokine expression can positively affect the immune system’s functioning [30]. The significant types of prebiotics include Inulin, fructooligosaccharides (FOS), galactooligosaccharides (GOS), and starch-derived oligosaccharides. Enterobacteria, Bacteroidetes, and Firmicutes are stimulated by GOS [31]. To archive the benefits of synbiotics, exploring the prebiotic utilization potential of these four isolated Probiotic strains mentioned above is noteworthy. The four strains showed varying growth, pH drop, and prebiotic scores in the presence of Inulin and FOS (Figure 5).
Conclusion
From this presented research, it can be concluded that the isolated probiotic Lactobacillus and Enterococcus strains display interesting probiotic characteristics such as antibacterial activity and auto-aggregation. These properties are advantageous for a probiotic culture to successfully colonize and compete with pathogens in the GIT environment. As well as the response to the simulated gastrointestinal transit (GIT) explains the ability of these strains to survive and colonize in gastric stress conditions. They also can ferment prebiotic-NDOs and can be used to prepare symbiotics, which impart beneficial health- enhancing effects on the host.
References
-
Pabari K, Pithva S, Kothari C, Purama RK, Kondepudi KK, et al. (2020) Evaluation of probiotic properties and prebiotic utilization potential of Weissella paramesenteroides isolated from fruits. Probiotics & Antimicro Prot 12: 1126-1138.
-
Khalid K (2011) An overview of lactic acid bacteria. International Journal of Biosciences 1(3): 1-13.
-
Weinberg Z, Shatz O, Chen Y, Yosef E, Nikbahat M, et al. (2007) Effect of lactic acid bacteria inoculants on in vitro digestibility of wheat and corn silages. Journal of Dairy Science 90(10): 4754-4762.
-
Abdelbasset M, Djamila K (2008) Antimicrobial activity of autochthonous lactic acid bacteria isolated from Algerian traditional fermented milk “Raïb”. African Journal of Biotechnology 7(16): 2908-2914.
-
Gibson GR, Probert HM, Van Loo J, Rastall RA, Roberfroid MB (2004) Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr Res Rev 17(2): 259-275.
-
Driessche MVD, Malderen NV, Geypens B, Ghoos Y, Veereman-Wauters G (2000) Lactose-[13C] ureide breath test: a new, noninvasive technique to determine orocecal transit time in children. J Pediatr Gastroenterol Nutr 31(4): 433-438.
-
Pithva S, Shekh S, Dave J, Vyas BRM (2014) Probiotic attributes of autochthonous _Lactobacillus rhamnosus_ strains of human origin. Appl Biochem Biotechnol 173(1): 259-277.
-
Scardaci R, Varese F, Manfredi M, Marengo E, Mazzoli R, et al. (2021) Enterococcus faecium NCIMB10415 responds to norepinephrine by altering protein profiles and phenotypic characters. J Proteomics 231: 104003.
-
Lodemann U, Strahlendorf J, Schierack P, Klingspor S, Aschenbach JR, et al. (2015) Effects of the probiotic Enterococcus faecium and pathogenic Escherichia coli strains in a pig and human epithelial intestinal cell model. Scientifica 2015: 235184.
-
Holzapfel WH, Wood BJ (2014) Lactic acid bacteria: biodiversity and taxonomy. In: Wood BJ, et al. (Eds.), The Chemistry of Food. Wiley Blackwell Publishing, Hoboken, New Jersey, United States.
-
Holzapfel W, Arini A, Aeschbacher M, Coppolecchia R, Pot B (2018) Enterococcus faecium SF68 as a model for efficacy and safety evaluation of pharmaceutical probiotics. Beneficial microbes 9(3): 375-388.
-
Kebede A, Viljoen BC, Gadaga TH, Narvhus JA, Lourens- Hattingh A (2007) The effect of container type on the growth of yeast and lactic acid bacteria during the production of Sethemi, South African spontaneously fermented milk. Food Research International 40(1): 33- 38.
-
Ambalam PS, Prajapati JB, Dave JM, Nair BM, Ljungh A, et al. (2009) Isolation and characterization of antimicrobial proteins produced by a potential probiotic strain of human _Lactobacillus rhamnosus_ 231 and its effect on selected human pathogens and food spoilage organisms. Microbial Ecology in Health and Disease 21(3-4): 211- 220.
-
Kneifel W (2000) In vitro growth behaviour of probiotic bacteria in culture media with carbohydrates of prebiotic importance. Microbial Ecology in Health and Disease 12(1): 27-34.
-
McVay CS, Rolfe RD (2000) In vitro and in vivo activities of nitazoxanide against Clostridium difficile. Antimicrobial agents and chemotherapy 44(9): 2254-2258.
-
Lindahl M, Faris A, Wadström T, Hjerten S (1981) A new test based on ‘salting out’ to measure relative surface hydrophobicity of bacterial cells. Biochim Biophys Acta 677(3-4): 471-476.
-
Pinto MGV, Franz CM, Schillinger U, Holzapfel WH (2006) Lactobacillus spp.with in vitro probiotic properties from human faeces and traditional fermented products. International journal of food microbiology 109(3): 205-214.
-
Charteris WP, Kelly PM, Morelli L, Collins JK (1998) Ingredient selection criteria for probiotic microorganisms in functional dairy foods. International journal of dairy technology 51(4): 123-136.
-
Mourad K, Nour-Eddine K (2006) In vitro preselection criteria for probiotic Lactobacillus plantarum strains of fermented olives origin. International Journal of Probiotics and Prebiotics 1(1): 27.
-
Dunne C, O’Mahony L, Murphy L, Thornton G, Morrissey D, et al. (2001) In vitro selection criteria for probiotic bacteria of human origin: correlation with in vivo findings. Am J Clin Nutr 73(2): 386s-392s.
-
Conway PL, Gorbach SL, Goldin BR (1987) Survival of lactic acid bacteria in the human stomach and adhesion to intestinal cells. J Dairy Sci 70(1): 1-12.
-
Minelli EB, Benini A, Marzotto M, Sbarbati A, Ruzzenente O, et al. (2004) Assessment of novel probiotic Lactobacillus casei strains for the production of functional dairy foods. International Dairy Journal 14(8): 723-736.
-
Noriega L, Gueimonde M, Sánchez B, Margolles A, de los Reyes-Gavilán CG (2004) Effect of the adaptation to high bile salts concentrations on glycosidic activity, survival at low pH, and cross-resistance to bile salts in Bifidobacterium. Int J Food Microbiol 94(1): 79-86.
-
Sathyabama S, Vijayabharathi R, Brunthadevi P, Ranjithkumar M, Priyadarisini V (2012) Screening for probiotic properties of strains isolated from feces of various human groups. J Microbiol 50(4): 603-612.
-
Liévin-Le MV, Servin AL (2014) Anti-infective activities of lactobacillus strains in the human intestinal microbiota: from probiotics to gastrointestinal anti-infectious biotherapeutic agents. Clin Microbiol Rev 27(2): 167- 199.
-
Kos BVZE, Šušković J, Vuković S, Šimpraga M, Frece J, et al. (2003) Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J Appl Microbiol 94(6): 981-987.
-
Kociubinski G, Zavaglia AG, Pérez PF, Disalvo EA, De Antoni GL (2002) Effect of bile components on the surface properties of bifidobacteria. J Dairy Res 69(2): 293-302.
-
Zavaglia AG, Kociubinski G, Pe´rez P, Disalvo E, Antoni GD (2002) Effect of bile on the lipid composition and surface properties of bifidobacteria. J Appl Microbiol 93(5): 794- 799.
-
Savi L (2004) Is There a Relationship between Chocolate Consumption and Headache? In: Nehlig A, (Ed.), Coffee, tea, chocolate, and the brain. CRC Press, Taylor & Francis, Boca Raton, Florida, 219-226.
-
Khangwal I, Shukla P (2019) Potential prebiotics and their transmission mechanisms: Recent approaches. J Food Drug Anal 27(3): 649-656.
-
Davani-Davari D, Negahdaripour M, Karimzadeh I, Seifan M, Mohkam M, et al. (2019) Prebiotics: definition, types, sources, mechanisms, and clinical applications. Foods 8(3): 92.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends